Body Temperature, Feed Intake and Plasma Metabolites of Indigenous Chicks by Oral Administration of Watermelon Rind Extract with Two different Colors of Flesh: Yellow and Red
One of the challenge in poultry production is heat stress because of it tend to increase body temperature in chickens. Watermelon rind (WR) contain abundant of amino acid L-Cirtulline (L-Cit). Oral administration of L-Cit has been shown to have hypothermic agent that could improve thermotolerance in layer chicks. However, no information has been available for comparison of oral administration of WR extract (WRE) with two different colors of flesh: yellow and red in indigenous chicken. The present study was performed to examine whether oral administration of yellow or red WRE affect feed intake, body temperature and plasma metabolites in indigenous chicks. 5-day-old indigenous chicks were given acute oral administration of WRE (10 ml/ kg) under control thermoneutral temperature (CT). Oral administration of both yellow and red WRE did not alter feed intake and body temperature and plasma metabolites in indigenous chicks. These results indicate that oral administration of yellow or red WRE may not alter thermoregulation, feed intake and plasma metabolites in indigenous chicks.
Introduction
The summer heat stress (HS) and global warming are becoming a serious concern around the world. It was reported by The Intergovernmental Panel Climate Change (IPCC) that the global surface temperature has been increasing [1]. Hence, summer HS is becoming more unbearable in not only in tropical but also subtropical countries. As environmental temperature is steadily increasing over the globe, HS is considered one of the major challenges for poultry production in some countries. Summer HS is causing a great economic loss in commercial poultry sector. Leeson, et al. [2] reported that the most important inhibiting factors to poultry production in hot regions is high ambient temperatures (HT), apparently because poultry cannot dissipate quickly the excess heat produced under HT, which subsequently leads to decrease performance with lower body weight gain in broilers and low egg production in layers with increasing mortality [3, 4]. Compared with other domestic animals, birds are more sensitive to HT [5] as birds lack sweat gland and they rely on evaporative cooling (panting) to keep them cool [6]. Birds respond to hot environments by changing the circulating levels of hormones, glucose, leukocytes, electrolytes, and the functions of organs [7]. Detrimental effects of HT not only affect performance parameters, but also various physiological [8, 9, 10, 11, 12] and immunological [13] adaptations of birds with the heat stress. The understanding of biochemical blood parameters is very important to define the biochemical profile, metabolism disorders, the energy metabolism, bone abnormalities, liver function and according of these factors could be used to determine the adaptation level of animals to climatic challenges [14]. Numerous reports have shown that in a HT, the concentration of glucose and total cholesterol in the blood decrease, which is an indicator of failure in homeostasis [15, 16]. Several nutritional supplementations have been recommended to alleviate the negative effects of heat stress on the body temperature in poultry. For instance, providing essential or non-essential amino acids with or without conjugated with emulsifier of lauric acid in their diets [17, 18, 19, 20, 21, 22, 23]. However, nutritional manipulation with low cost involvement is a common approach in poultry industry [24].
Previous studies have been reported that oral administration of L-citrulline (L-Cit) decreased body temperature in layer chicks and afforded a degree of thermotolerance [25]. It is well known that the inclusion of synthetic L-Cit in poultry diets has not been approved in some countries, therefore one of alternative way might be the use of natural source of L-Cit from watermelon. Currently, this fruit is the third most popular fruit around the world and it also the largest consumed by people in hot weather, King, et al. [26] stated that according to designation of flesh color of watermelon, there are eight color: white, salmon yellow, orange, crimson red, scarlet red, pale yellow, canary yellow and green. Instead of its flesh, watermelon rind (WR) as agricultural waste has been reported contains rich of L-Cit [27, 28]. The different color of flesh watermelon affect differences carotenoid contents [29]. We also have determined that L-Cit contents in red WR dried powder (WRP) was a greater than yellow WRP (unpublished data). Recently, it was reported that watermelon rind extract (WRE) contains 6638 pmol/mg [30]. Furthermore, Nguyen, et al. [30] revealed that oral administration of WRE significantly decreased body temperature in layer chicks. Similarly, in the previous study we found that oral administration of WRE decreased body temperature in broiler chicks (unpublished data). However, to the best of our knowledge, there has been no reports regarding the effect oral administration of different colour especially yellow or red WRE as a natural source of L-Cit on feed intake, body temperatures and plasma metabolites in indigenous chicks.
Materials and Methods
Experiment
Oral administration of WRE on feed intake, body temperature and plasma metabolites in indigenous chicks. One day-old (DOC) indigenous chicks were purchased from a local hatchery and housed in a wire-meshed cage (50 x 35 x 33 cm) in a group (2025 birds) at a constant temperature of 30±1°C and with continuous light. Chicks were all of the same age and were housed without any adult. Feed (Charoen Phokpand) and water were provided ad libitum. Feed composition is presented at Table 1. One day before the experiment, chicks (4 days old) were reared individually and assigned for treatment and control groups on the basis of their body weight in order to produce uniform groups.
| Nutrient | Commercial ration |
|---|---|
| Crude Protein (%) | 23.5 |
| Crude Fiber (%) | 1.88 |
| Crude Fat (%) | 5.87 |
| Ca (%) | 0.29 |
| P (%) | 0.15 |
| ME (Kcal/kg) | 3,050 |
Table 1: The percentage of nutrient content in commercial ration.
Ca: Calcium, P: Phosphor, ME: Metabolizable Energy; *Commercial feed: CP511 PT, Charoen Pokphand, Indonesia; Mineral Premix: Supplemented for kg of the diets: Vit. A, 12000 IU; D3, 2000 IU; E, 20 mg; K3, 3 mg; B2, 7 mg; B3, 12 mg; B5, 3 mg; B12, 0.03 mg; biotin, 0.1 mg; choline chloride, 300 mg; Mn, 130 mg; Fe, 70 mg; Zn, 60 mg; Cu,12 mg; I,1 mg; Se, 0.2 mg, and adequate antioxidant. Table 1:** The percentage of nutrient content in commercial ration.
Following a habituation period, chicks were randomly selected and divided into three groups each consisting of 10 chicks. The birds were reared individually in experimental cages and had ad libitum access to diet up to the time of the experiments. On the day of the experiment, each chick (5 days old) was orally administered distilled water, red WRE or yellow WRE by the elastic plastic needle on small syringe. The birds were fed ad libitum diets for 2 h immediately after the treatment. Feed intake (at 30, 60 and 120 min) was measured by calculating the reduction in the amount of feed consumed from a pre-weighed feeder. Similarly, body temperature of chicks was measured at 30, 60 and 120 min after. The weight of the feeder was measured using an electric digital balance. At the end of the experiments, birds were decapitated following anesthesia with isoflurane (Mylan Inc., Japan). Blood samples were collected in heparinized tubes and centrifuged for 15 min at 5,000g, and the plasma was collected and stored at 20oC until analysis took place.
Body Temperature Measurement and of Plasma Metabolites
A digital thermometer with an accuracy of ± 0.1°C (Thermalert TH-5, Physitemp Instruments Inc., USA) by inserting the thermistor probe into the cloaca to a depth of 2 cm was used to measure the rectal temperature of chicks. The plasma metabolites, including glucose (Glu), total cholesterol (TCHO), total protein (TP), triacylglycerol (TG) were measured by Microlab 300 (Vital Scientific, Netherland) as per the manufacturer’s instructions. The samples were assayed together and in a random sequence for each metabolite.
Statistical Analysis
A repeated-measures two-way ANOVA were applied for the analysis of feed intake and body temperature in Experiment 1. Plasma metabolites were applied by one way ANOVA. Tukey-Kramer test was performed as a post-hoc test. Significant differences were denoted as P<0.05. Values were presented as means ± S.E.M. Statistical analysis were the commercially available package Statview (1998). All data in each group were first subjected to a Thompson rejection test to eliminate outliers (P<0.01), and the remaining data were used for the analysis among groups.
Results and Discussion
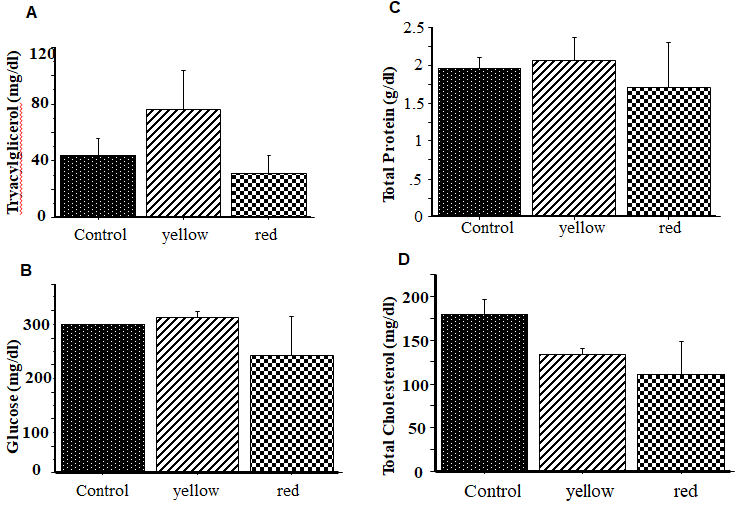
Experiment Effects of oral administration of yellow or red WRE on feed intake, body temperature and plasma metabolites in indigenous chicks. As shown in Figure 1 body temperature did not change significantly (P<0.05) following oral administration of yellow or red WRE. Similarly, oral administration of yellow or red WRE did not significantly (P>0.05) alter feed intake (data not shown). Figure 2 shows the effects of yellow or red WRE on plasma metabolites. Similarly, oral administration of WRE did not significantly (P>0.05) alter TCHO, TG, Glu and TP. In the current study, we confirmed that oral administration both red and yellow WRE did not affect feed intake in indigenous chicks compared with a control group (data not shown). This results was consistent with previous studies. Ngunyen, et al. [30] demonstrated that oral administration of WRE failed to affect feed intake in 14-days old layer chicks. Other supportive data was reported by Chowdury, et al. [32] who demonstrated that oral administration of L-Cit did not influence feed intake in layer chicks. Similarly, Erwan, et al. [33] revealed that oral administration of L-Cit did not alter feed intake in KUB chicks. Additionally, Uyanga, et al. [34] and reported that dietary supplementation of L-Cit did not influence feed intake in laying hens. Furthermore, Poduri, et al. [35] demonstrated that supplemention of WRE did not alter feed intake in mice. In the light of information mentioned above and of the results from the present study, it might be suggested that oral administration of WRE or L-Cit seemed to similar effects on feed intake in domestic chicks or indigenous chicks and mice.
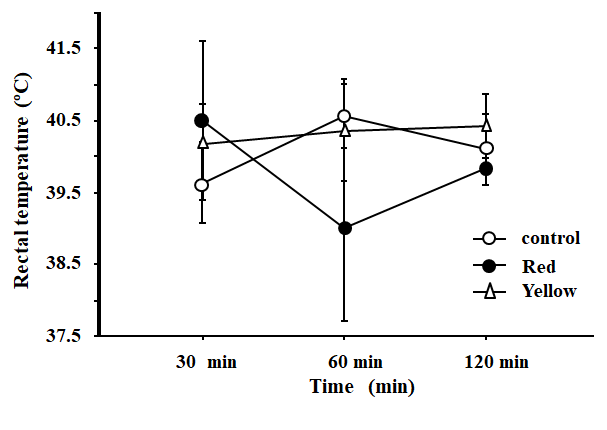
We also demonstrated here that both red and yellow WRE did not alter body temperature in indigenous chicks. This finding is consistent with Uyanga, et al. [34] who revealed that dietary supplementation of L-Cit did not influence body temperature in laying hens. In addition, Poduri, et al. [35] also demontrated that there ware no differences in body temperature when mices were supplemented by WRE. Furthermore, Chowdhury, et al. [25] revealed that intracerebroventricular (i.c.v) of L-Cit failed to affect body temperature in layer chicks. However, the effect of WRE on body temperature differed with other reports. Nguyen, et al. [30] demontrated that body temperature was significantly decreased when layer chicks were orally administrated with WRE. Additionally, Chowdhury, et al. [25] revealed that oral administration of L-Cit significantly lowered body temperature in layer chicks. The reason for these discrepancies on body temperature in respon to oral administration of WRE or both i.c.v. and oral administration of L-Cit in birds perhaps due to varietas of watermelon, variations in experimental period, differences in doses, breed, strain or species. It is well known that indigenous breeds of chickens in tropical countries are better able to withstand high ambient temperatures than faster growing strains. Further experimentation would be needed to clarify the factors involved in this disparity.
Many factors may influence metabolic alterations in mammal and poultry such as physiological state, pharmacological condition, age, husbandry condition, genetic type and feed [22, 33, 35]. In general, the status of physiological of birds can be determined according to their hematological parameters and their concentrations are varied affected by dietary supplements. The absence of significant changes in the concentrations of TCHO, TG, Glu and TP in current study was consistent with previous study [32], Uyanga, et al. These findings alse were similar to the previous findings [33] who reported that there was no changes these plasma metabolites in KUB chicks following oral administration of L-Cit. This non-significant effect of WRE suggests that it does not affect blood cell formation, constituents and their function. We speculate that the short period of oral administration of WRE was not enough to alter the metabolic in the plasma metabolites.
Conclusion
To our knowledge, this is the first study to report oral administration of WRE may not alter feed intake body temperature and plasma metabolites in indigenous chicks.
Conflict of Interest
The authors declare no conflict of interest.
Acknowledgement
This work was supported by The Directorate of Islamic Religious Higher Education of The Indonesian Ministry of Religion through a Grant-in-Aid for Scientific Research no 4842 to EE.
References
-
(2013) Intergovernmental Panel on Climate Change (IPCC). Climate Change. The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change.
-
Leeson S (1986) Nutritional considerations of poultry during heat stress. Worlds Poult Sci J 42: 69-81.
-
Teeter RG, Smith MO, Owens FN, Arp SC, Sangiah S, et al. (1985) Chronic heat stress and respiratory alkalosis: Occurrence and treatment in broiler chicks. Poult Sci 64: 1060-1064.
-
Quinteiro Filho WM, Ribeiro A, Ferraz-de-Paula V, Pinheiro ML, Sakai M, et al. (2010) Heat stress impairs performance parameters, induces intestinal injury, and decreases macrophage activity in broiler chickens. Poult Sci 89(9): 1905-1914.
-
Geraert PA, Guillaumin S, Leclercq B (1993) Are genetically lean broilers more resistant to hot climate?. Br Poult Sci 34(4): 643-653.
-
Ensminger ME, Oldfield JE, Heinemann WW (1990) Feeds and Nutrition. The Ensminger Publishing Company, Clovis pp: 8-120.
-
Geraert PA, Padilha JC, Guillaumin S (1996) Metabolic and endocrine changes induced by chronic heat exposure in broiler chickens: growth performance, body composition and energy retention. Br J Nutr 2: 195-204.
-
Koelkebeck KW, Odom TW (1995) Laying hen responses to acute heat stress and carbon dioxide supplementation: Changes in plasma enzymes, metabolites and electrolytes. Comp Biochem Physiol 112(1): 119-122.
-
Maak S, Melesse A, Schmidt R, Schneider F, Von Lengerken G (2003) Effect of long-term heat exposure on peripheral concentrations of heat shock protein 70 (Hsp70) and hormones in laying hens with different genotypes. Br Poult Sci 44(1): 133-138.
-
Tao X, Zhang ZY, Dong H, Zhang H, Xin H (2006) Responses of thyroid hormones of market-size broilers to thermoneutral constant and warm cyclic temperatures. Poult Sci 85(9): 1520-1528.
-
Chowdhury VS, Tomonaga S, Nishimura S, Tabata S, Furuse M (2012) Physiological and behavioral responses of young chicks to high ambient temperature. J Poult Sci 49: 212-218.
-
Chowdhury VS, Tomonaga S, Ikegami T, Erwan E, Ito K, et al. (2014) Oxidative damage and brain concentrations of free amino acid in chicks exposed to high ambient temperature. Comp Biochem Physiol A Mol Integr Physiol 169C: 70-76.
-
Star L, Juul Madsen HR, Decuypere E, Nieuwland MGB, de Vries Reilingh G, et al. (2009) Effect of early life thermal conditioning and immune challenge on thermotolerance and humoral immune competence in adult laying hens. Poult Sci 88: 2253-2261.
-
Swenson MJ, Reece WO (2006) Dukes – Fisiologia dos Animais Domésticos.Rio de Janeiro: Guanabara Koogan S.A.
-
Ocak, Guney, Ocak S, Guney O (2010) Physiological responses and some blood parameters of bucks under Mediterranean climate condition. Anadolu J Agric Sci 25: 113-119.
-
Ribeiro, Ribeiro NL, Costa RG, Pimenta Filho EC, Ribeiro MN, et al. (2018) Effects of the dry and the rainy season on endocrine and physiologic profiles of goats in the Brazilian semi-arid region. Ital J Anim Sci 17: 454-461.
-
Mendes AA, Watkins SE, England JA, Saleh EA, Waldroup AL, et al. (1997) Influence of dietary lysine levels and arginine:lysine ratios on performance of broilers exposed to heat or cold stress during the period of three to six weeks of age. Poult Sci 76: 472-481.
-
Rose SP, Salah Uddin M (1997) Effect of temperature on the response of broiler chickens to dietary lysine balance. Br Poult Sci 38: S36-S7.
-
Daghir NJ, Farran MT, Barbour GW, Beck MM (2003) Nutritive value of high-oil corn grown under semi-arid conditions and its impact on broiler performance and carcass composition. Poult Sci 82: 267-271.
-
Hamasu K, Shigemi K, Tsuneyoshi Y, Yamane H, Sato H, et al. (2010) Intracerebroventricular injection of Lproline and D-proline induces sedative and hypnotic effects by different mechanisms under an acute stressful condition in chicks. Amino Acids 38: 57-64.
-
Erwan E, Tomonaga S, Yoshida J, Nagasawa M, Ogino Y, et al. (2012) Central administration of L- and Daspartate attenuates stress behaviors by social isolation and CRF in neonatal chicks. Amino Acids 43: 1969-1976.
-
Erwan E, Chowdhury VS, Nagasawa M, Goda R, Otsuka T, et al. (2014) Oral administration of D-aspartate, but not Laspartate, depresses rectal temperature and alters plasma metabolites in chicks. Life Sci 109: 65-71.
-
Yoshida J, Erwan E, Chowdhury VS, Ogino Y, Shigemura A, et al. (2005) Comparison of centrally injected tryptophan-related substances inducing sedation in acute isolation stress-induced neonatal chicks. Pharmacology Biochemistry and Behavior 129: 1-6.
-
Shane SM (1998) Factors influence health and performance of poultry in hot climates. Poultry Biology 1: 247-269.
-
Chowdhury VS, Shigemura A, Erwan E, Ito K, Bahry MA, et al. (2015) Oral administration of L-citrulline, but not L-arginine or L-ornithine, acts as a hypothermic agent in chicks. J Poult Sci 52: 331-335.
-
King SR, Davis AR, Bang H (2009) New flesh colors in watermelon?. HortScience 44: 576.
-
Rimando, Veazie P, Rimando AM, Perkins Veazie P (2005) Determination of citrulline in watermelon rind. Journal of Chromatography A 1078(1-2): 196-200.
-
Tarazona Díaz MP, Viegas J, Moldao Martins M, Aguayo E (2011) Bioactive compounds from flesh and by-product of fresh-cut watermelon cultivars. Journal of the Science of Food and Agriculture 91: 805-812.
-
Zao (2013) Studies on carotenoids in watermelon flesh. Agricultural Sciences 4(7): 13-20.
-
Nguyen LTN, Han G, Yang H, Ikeda H, Eltahan HM, et al. (2019) Dried watermelon rind mash diet increases plasma L-citrulline level in chicks. J Poult Sci 56: 65-70.
-
SAS (1998) Stat View, Version 5, SAS Institute Institute, Cary, NC, USA.
-
Chowdhury VS, Han G, Bahry MA, Tran PV, Do PH, et al. (2017) L-Citrulline acts as potential hypothermic agent to afford thermotolerance in chicks. J Therm Biol 69: 163-170.
-
Erwan E, Adelina T, Koto A, Maslami V (2020) The potency of oral administration of L-citrulline as anti heat stress agent in KUB chickens. J World‘s Poult Res 10: 36- 40.
-
Uyanga VA, Jiao H, Zhao J (2020) Dietary L-citrulline supplementation modulates nitric oxide synthesis and anti-oxidant status of laying hens during summer season. J Animal Sci Biotechnol 11: 103.
-
Poduri A, Rateri DL, Saha SK, Saha S, Daugherty A (2013) Citrullus lanatus ‚senti nel‘ (watermelon) extract reduces atherosclerosis in LDL receptor-deficient mice. J Nutr Biochem 24(5): 882-886.
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System