Bioactivity of Crude Extracts of Secondary Metabolites of the Endophytes Phyllosticta capitalensis (Tg06) and Curvularia sp. (G6-32) against Aedes aegypti Larvae (L.1762)
Aedes aegypti is known to transmit several arboviruses, causing economic impacts in several countries. Biolarvicides produced by endophytic fungi are a potential tool in combating mosquito proliferation. The objective of this work was to examine the potential of secondary metabolites of the endophytic fungi Phyllosticta capitalensis strain Tg06 and Curvularia sp. strain G6-32 in the control of third instar Aedes aegypti larvae. To extract the metabolites, the fermented broth was filtered with the aid of a glass and cotton funnel and then centrifuged in 50 mL conical tubes at 1400 rcf for 15 minutes. The bioassays were carried out for five days in a biological chamber at 25 ± 2°C, with a 12/12 photoperiod. The tests were performed on the following solutions: 5% dimethylsulfoxide (DMSO) and water, control solution with distilled water and a solution of crude extract of metabolites diluted in 5% DMSO and water at a concentration of 50 μg/mL. The crude extract of Phyllosticta capitalensis Tg06 caused mortality in the larvae throughout the period evaluated and Curvularia sp. G6-32 induced the interruption of larval development. The use of crude extracts of endophytic microorganisms Curvularia sp. and P. capitalensis appears to be a promising strategy for the biocontrol of Aedes aegypti and could be an alternative to the use of conventional chemical insecticides, which would result in reduction of environmental contamination and toxic effects for animals, plants, and humans.
Introduction
Aedes aegypti (Diptera: Culicidae) is able to transmit several arboviruses, with Dengue, Chikungunya, and Zika being the main endemic diseases spread by this mosquito when infected with the virus [1]. The substances that are currently used as chemical and biological controls on the insect population are limited because of the increasing resistance mechanisms of these vector-insects over time [2], in addition to not being selective for associated fauna. One of the emerging approaches to fight the eggs and larvae of Ae. aegypti is through the application of biolarvicides, which can minimize consequences for the health of the human population and the environment as well as reduce the selection of pesticide-resistant insects, according to a 2020 report by the Pan American Health Organization [3]. Additionally, improved control of vector-insect populations can contribute to mitigating the transmission of serotypes DenV-1, DenV-2, DenV-3, and DenV-4 of the virus that causes Dengue Fever, which affects numerous countries.
Microorganisms have peculiar biochemical pathways which synthesize specific metabolites of plant species [4], including promising, efficient, and environmentally safe natural compounds that can serve as tools for reducing the larval Ae. aegypti (Linnaeus, 1762) mosquito population [5]. Innovative compounds, such as penicillin, produced by Penicillium notatum fungi, opened paths for the exploration of new bioactive compounds for commercial use belonging to several structural groups, such as alkaloids, steroids, phenols, flavonoids, peptides, quinones, and terpenoids [6].
Numerous primary and secondary metabolites from endophytic fungi have been highlighted in research owing to the biological potential of their molecules, with direct application to human health [7, 8]. According to Silva [9], a vast number of reports in the literature establish that these microorganisms produce bioactive compounds with potential application in several areas of interest to medical and pharmaceutical industries, such as anticancer, antioxidant, antimicrobial, and antiviral activities.
In nature, endophytic fungi adapt to the tissues of plant parts without showing any symptoms of fungal disease and support the physiological and ecological attributes of the host plant or forest species [6]. When symbiotic interactions occur, these microorganisms induce the production of secondary metabolites that confer advantages to the plant [10], and these have aroused biotechnological and industrial interest because they have properties that can be applied in numerous sub-areas of biotechnology and related areas, especially for human health.
Among the biodiversity of endophytic microorganisms, two species stand out for their capacity to produce bioactive secondary metabolites. Crude extract from Phyllosticta capitalensis strain Tg06, which was isolated from the Tibouchina granulosa (Vell.) Cogn (Melastomataceae), showed antiprotozoal activity against Leishmania amazonensis, L. infantum, and Trypanosoma cruzi, and produced 18 compounds, including fatty acids based on linoleic acid and derivatives [11]. Curvularia sp. strain G6-32, which was isolated from the medicinal plant Sapindus saponaria L. (Sapindaceae), produced secondary metabolites containing (-)-Asperpentyn, with antioxidant and anticholinesterase activities [12].
This study reports the potential of secondary metabolites of the endophytic fungi Phyllosticta capitalensis Tg06 and Curvularia sp. G6-32 in the control of Ae. Aegypti at the third instar larvae stage.
Materials and Methods
Endophytic Fungi
The endophytic fungi Phyllosticta capitalensis Tg06 and Curvularia sp. G6-32 used in this study were obtained from the Collection of Endophytic and Environmental Microorganisms of the Microbial Biotechnology Laboratory of the State University of Maringá, Paraná - Brazil (CMEA/ LBIOMIC-UEM).
The endophytes were activated in dextrose potato agar (BDA - Himedia®, Bombay, India), and incubated for 7 days at 28°C. Then, three 6 mm diameter discs of the fungal colonies were transferred to an Erlenmeyer flask containing 250 mL of potato dextrose broth (BD - Acumedia®), and were incubated for 21 days at 28°C.
Obtaining the Crude Extract
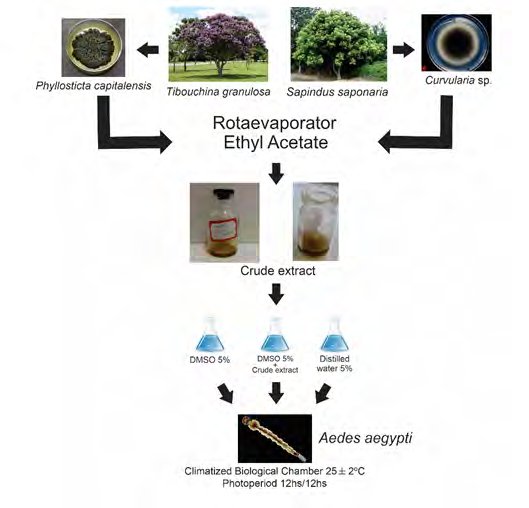
For the extraction of secondary metabolites, the fermented broth was filtered with the aid of a glass and cotton funnel and subsequently centrifuged in 50 mL conical tubes at 1400 rcf for 15 minutes. The supernatant was transferred to a separating funnel, and the solvent was added in a 1:5 ratio (ethyl acetate: fermented broth), and this step was repeated three times. The solvent was collected and subjected to rotary evaporation at 37°C and pressure of 600 mmHg (Tecnal TE-210). The metabolites were lyophilized and then stored (Figure 1).
Larvicidal Assay
The larvicidal activity of the extract was evaluated using Ae. aegypti maintained at the Laboratory of General and Medical Entomology at the State University of Londrina. Larvae were obtained from a permanent colony maintained at a controlled temperature of 25 ± 2°C and a relative humidity of 70 ± 5%, with a 12/12 photoperiod.
Three solutions were prepared: (1) a 5% dimethyl sulfoxide (DMSO) and water solution, (2) a control solution with distilled water, and (3) a solution of crude extract of metabolites diluted in 5% DMSO and water at a concentration of 50 µg/mL. For each bioassay, 50 mL of each test solution was applied to 12 third-stage larvae of Ae. aegypti, performed in duplicate. Observations were performed daily for five days. Larvae were considered dead when there was a total absence of movement and darkening of the body (Figure 1).
Statistical Analysis
The values obtained were statistically evaluated using analysis of variance (ANOVA) and the means were compared using the Scott–Knott test (p < 0.05 was considered statistically significant) to correct for multiple comparisons. Statistical tests were performed using the Sisvar 5.6 statistical program. To adjust a prediction curve to the data, a non-linear regression by using the least squares method, through a Three Parameter Logistic function as represented by Equation 1 was used.
( ) 1 b x c a y e − − = −
(Equation 1) The parameters a, b, and c represent, respectively, the carrying capacity (Upper limit), the slope around the inflection point, and the horizontal displacement of the function around the sampling points. The analysis of the adjustment curve was performed using the R programming language, a free software widely used in statistical analyses.
Aiming to determine a mathematical function that optimally describes the collected data, the nonlinear model was chosen because it describes survival analysis behaviors. The nls function from the nlme package of the R software was used to calculate the optimal parameters of the fitted curve. Because we use numerical methods to seek the best fitting parameters, it is relevant to highlight the values of the initial guess in order to allow the repeatability of the analysis by other researchers.
Results and Discussion
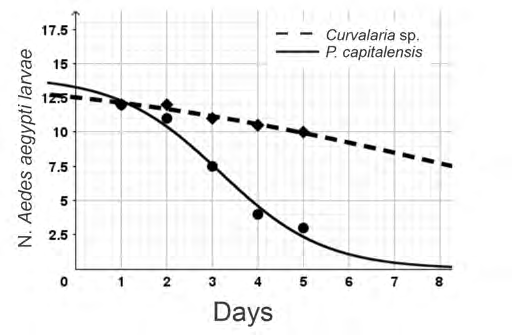
The crude extract of secondary metabolites of the endophytic fungus P. capitalensis showed larvicidal activity in Ae. Aegypti throughout the evaluation period, showing statistical difference starting from the 4th day (Figure 2 & Table 1). No statistically significant difference was observed between the water and DMSO samples (Table 2). When taking the mean of the repetition of the five values obtained from the larval count of Ae. aegypti in the metabolites of Phyllosticta capitalensis, and taking the initial parametric vector as (a = 12.5, b = –0.015, c = 7.51), the summary of best fit parameter results provides the following function:
( ) 0.8844 3.1880 14.0765 (Equation 2)
1 x y e − = +
The fitted curve (Figure 2) described the behavior of the collected data accurately. In fact, in the case of the adjustment for P. capitalensis Tg06 metabolites, the Residual Standard Error calculated using R was 0.7804. The adjusted parameters a, b, and c for Curvularia sp. G6-32 were 14.4808, –0.2157, and 8.6250, respectively. Although the crude extract of Curvularia sp. G6-32 caused a lower mortality than the extract of P. capitalensis Tg06, with statistically significant differences starting from day 5 (Table 2), it showed larval growth arrest activity, which did not allow the mosquitos to progress to the fourth larval stage, and thus they did not reach the vector phase.
| Days | Phyllosticta capitalensis Tg06 | Curvularia sp. G6-32 | ||
|---|---|---|---|---|
| Means | Classes* | Means | Classes* | |
| 1 | 12 | a1 | 12 | a1 |
| 2 | 11,66 | a1 | 12 | a1 |
| 3 | 10,5 | a1 | 11,66 | a1 |
| 4 | 8,66 | a2 | 11,5 | a1 |
| 5 | 8,33 | a2 | 11 | a2 |
Table 1: Statistical mean ANOVA compared using the Scott-Knott test of larval survival for each day of treatment using crude extr
*Means within the same class do not differ statistically from each other. Table 1: Statistical mean ANOVA compared using the Scott-Knott test of larval survival for each day of treatment using crude extract of the endophytic fungus Phyllosticta capitalensis Tg06 and Curvularia sp. G6-32.
| Solution | Phyllosticta capitalensis Tg06 | Curvularia sp. G6-32 | ||
|---|---|---|---|---|
| Means | Classes* | Means | Classes* | |
| Metabolic extract | 7.5 | a1 | 11.1 | a1 |
| Distilled water | 11.6 | a2 | 12 | a2 |
| DMSO 5% | 11.6 | a2 | 11.8 | a2 |
Table 2: Statistical mean ANOVA compared using the Scott–Knott test between solutions tested against Ae. Aegypti larvae.
*Means within the same class do not differ statistically from each other. Table 2: Statistical mean ANOVA compared using the Scott–Knott test between solutions tested against Ae. Aegypti larvae.
This is the first report of the action of secondary metabolites of these endophytes against Ae. aegypti, and it is known that some metabolites produced in common by other fungi have already been described with insecticidal activity, which corroborates the results obtained in this work.
The endophytic strain P. capitalensis Tg06 used in this study was previously shown to produce compounds similar to linoleic acid [11], a chemical compound that has been reported to show activity against 4th instar larvae of Ae.
aegypti with an LC50 of 100 µg/mL [13]. In the present study, the extract of this endophyte showed 75% activity against larvae in the 3rd instar when using a lower concentration of extract. Other studies report compounds found in different species of the Phyllosticta genus, such as heptelidic acid (HA), an inhibitor of glyceraldehyde-3-phosphate dehydrogenase (GAPDH), which inhibits glycolysis in mosquitoes [14], and can cause the death of the caterpillar Choristoneura fumiferana. In addition, phyllostin, which belongs to the phytotoxin family, showed insecticidal activity against Plutella xylostella larvae, in addition to reducing the fecundity of this pest [16].
Several studies reported in the literature show the action of secondary metabolites of fungi with larvicidal action. Ragavendran, et al. [17] evaluated the toxicity of the secondary metabolite of Penicillum sp. isolated from the soil against larvae of Ae. aegypti at a range of concentrations, obtaining optimal results at a concentration of 500 µg/ mL, with mortality of stage 1–4 instar larvae, in addition to arrest of growth and larval development using the lowest tested concentration of metabolites. Ragavendran et al [18] evaluated the larvicidal and ovicidal efficacy of Aspergillus terreus metabolites against Ae. aegypti and verified histopathological changes such as cuticle demelanization and shrinkage of the inner cuticle of anal papillae, with zero hatchability of mosquito eggs observed at a concentration of 500 μg/mL. Abutaha, et al. [19] evaluated the extract of metabolites of the endophyte Cochliobolus spicifer (Nelson), which is isolated from the date palm Phoenix dactylifera (Linnaeus), against larvae of Aedes caspius (Pallas) and Culex pipiens (Linnaeus), obtaining arrest of the mosquito life cycle and toxic effect in 3rd instar larvae stage, with 100% mortality at concentrations of 300 ppm.
The extract of secondary metabolites of the endophyte Curvularia sp. G6-32 (isolated from S. saponaria) paralyzed larval development at the concentration tested. The extract from this endophytic strain previously presented anticholinesterase activity [12], which may have influenced the larvae’s defense system against the extract, making them vulnerable to it because esterase, oxidase and transferase enzymes contribute at least part of the mechanism of resistance of mosquitoes to insecticides [20]. Baskar, et al. [21] evaluated the larvicidal effect of the endophytic fungus Aspergillus tamarii, which is isolated from the stem of the Opuntia ficus-indica cactus, against Ae. aegypti and verified that the metabolite extract presented an LC50 of 18.69 µg/mL and an LC90 of 8.29 µg/mL for the tested concentration over a 48-h period, in addition to significantly inhibiting the larval activity of acetylcholinesterase.
The genus Curvularia has been shown to produce various bioactive compounds with cytotoxic, anticholinesterase, antioxidant, antimicrobial and phytotoxic activities, among others [12, 22, 23]. Furthermore, studies report that the interaction between fungi and plants, especially plants with medicinal properties, can cause metabolic changes in the endophyte and/or in the host plant, which can be an alternative source for the production of bioactive molecules with various technological, pharmaceutical and industrial applications [24, 25, 26]. Barreto, et al. [27] evaluated the crude ethanol extract of the fruit peel of S. saponaria against Ae. aegypti and reported a CL90 of 134.1 ppm. Toxic effects were observed in histopathological examinations and total or partial cell destruction, high cytoplasmic vacuolization, epithelial cell hypersecretion and epithelial paving were verified.
In the pursuit of improving global human health, there is a lack of new strategies to fight against the larvae of this mosquito to reduce spread of deadly viruses, which is of great economic importance. The results presented here show a promising strategy for the biocontrol of Ae. aegypti as a possible alternative to the use of conventional chemical insecticides, reducing environmental contamination and toxic effects for animals and humans.
Conclusion
The use of crude extracts of endophytic microorganisms Curvularia sp. G6-32 and Phyllosticta capitalensis Tg06 show evidence to be a promising biocontrol strategy against Ae. aegypti and constitute an alternative to the use of conventional chemical insecticides, and consequently a reduction in environmental contamination and toxic effects for animals, plants and humans.
Conflicts of Interest
The authors declare that they do not have any conflicts of interest to disclose.
References
-
Andrade JN, Costa Neto EM, Brandão HN, Lucchese AM, Nascimento Neto EB, et al (2021) Evaluation of Phyllanthus acuminatus Vahl (Phyllantaceae) extracts in the mortality of Aedes aegypti Linnaeus larvae, 1762 Culicidae. Brazilian Journal of Development 7(1): 5278- 5295.
-
Santos DC (2018) Avaliação da atividade larvicida de extratos vegetais do cerrado sobre Aedes aegypti**.** Trabalho de Conclusão de Curso (Bacharelado em Farmácia), Universidade de Brasília, Brasília pp: 53.
-
PAHO (2020) Ippc, International Plant Protection Convention. Overview**.**
-
De Oliveira SLA (2021) Prospecção de metabólitos secundários produzidos por fungos marinhos com atividade antifúngica. Dissertation, Biochemistry and Molecular Biology, Federal University of Alagoas, pp: 1-94.
-
Garcez WS, Garcez FR, Silva LMGE, Sarmento UC (2013) Naturally Occurring Plant Compounds with Larvicidal Activity Against Aedes aegypti. Rev. Virtual Quim 5(3): 363-393
-
Manganyi MC, Ateba CN (2020) Untapped Potentials of Endophytic Fungi: A Review of Novel Bioactive Compounds with Biological Applications. Microorganisms 8(12): 1934.
-
Dantas SBS, Alves FAM, Chapla VM (2021) Chemical, Diversity and Biotechnological Potential of Endophytic Fungi Isolated From Brazilian Cerrado Plants. Biota Neotropica 21(2): e20201069.
-
Moura PF, Betim FCM, Oliveira CF de, Dias JFG, Montruchio DP, et al. (2021) Atividade larvicida de extratos de Diplodia pinea frente à Aedes aegypti. Research, Society and Development 10(2): e6710212295.
-
Silva IP (2014) Endophytic fungi: alternative source to secondary plant metabolites**.** Enciclopédia Biosfera, Centro Científico Conhecer.
-
Pamphile JA, Costa AT, Rosseto P, Polonio JC, Pereira JO, et al. (2017) Biotechnological Applications of Secondary Metabolites Extracted from endophytic fungi: the case of Colletotrichum sp Revista Uningá 53(1): 113-119.
-
Golias HC, Polonio JC, Dos Santos Ribeiro MA, Polli AD, da Silva AA, et al. (2020). Tibouchina granulosa (Vell.) Cogn (Melastomataceae) as source of endophytic fungi: Isolation, identification, and antiprotozoal activity of metabolites from Phyllosticta capitalensis. Braz J Microbiol 51(2): 557-569.
-
Polli AD, Ribeiro MAS, Garcia A, Polonio JC, Santos CM, et al. (2020) Secondary metabolites of Curvularia sp. G6-32, an endophyte of Sapindus saponaria, with antioxidant and anticholinesterasic properties. Natural Product Research 16: 1-6.
-
Ramsewak RS, Nair MG, Murugesan S, Mattson WJ, Zasada J (2001) Insecticidal fatty acids and triglycerides from Dirca palustris. Journal of Agricultural and Food Chemistry 49(12): 5852-5856.
-
Kato M, Sakai K, Endo A (1992) Koningic acid (heptelidic acid) inhibition of glyceraldehyde 3-phosphate dehydrogenases from various sources. Biochem Biophys Acta 1120(1): 113-116.
-
Calhoun LA, Findaly JA, Miller JD, Whitney NJ (1992) Metabolites toxic to spruce budworm from balsam fir needle endophytes. Mycol Res 96: 281-286.
-
Ratnaweera PB, Jayasundara JMNM, Herath HHMSD, Williams DE, Rajapaksha SU, et al. (2020) Antifeedant, contact toxicity and oviposition deterrent effects of phyllostine acetate and phyllostine isolated from the endophytic fungus Diaporthe miriciae against Plutella xylostella larvae. Pest Manag Sci 76: 1541-1548.
-
Ragavendran C, Manigandan V, Kamaraj C, Balasubramani G, Prakash JS, et al. (2019) Larvicidal, Histopathological, Antibacterial Activity of Indigenous Fungus Penicillium sp. Against Aedes aegypti L and Culex quinquefasciatus (Say) (Diptera: Culicidae) and Its Acetylcholinesterase Inhibition and Toxicity Assessment of Zebrafish (Danio rerio). Frontiers in Microbiology pp: 10.
-
Ragavendran C, Srinivasan R, Kim M, Natarajan D (2018) Aspergillus terreus (Trichocomaceae): A Natural, Eco-Friendly Mycoinsecticide for Control of Malaria, Filariasis, Dengue Vectors and Its Toxicity Assessment Against an Aquatic Model Organism Artemia nauplii. Frontiers in Pharmacology 9: 1355.
-
Abutaha N, Mashaly AMA, Al Mekhlafi FA, Farooq M, Al Shami M, et al. (2015) Larvicidal activity of endophytic fungal extract of Cochliobolus spicifer (Pleosporales: Pleosporaceae) on Aedes caspius and Culex pipiens (Diptera: Culicidae). Applied entomology and Zoology 50(3): 405-414.
-
Brodgon WC, McAllister JC (1998) Insecticide Resistance and Vector Control. Emerging Infectious Diseases 4(4): 605-613.
-
Baskar K, Chinnasamy R, Pandy K, Venkatesan M, Sebastian PJ, et al. (2020) Larvicidal and histopathology effect of endophytic fungal extracts of Aspergillus tamarii against Aedes aegypti and Culex quinquefasciatus. Heliyon 6(10): e05331.
-
Zhang YL, Kong LC, Jiang DH, Yin CP, Cai QM, et al. (2011) Phytotoxic and antifungal metabolites from Curvularia sp. FH01 isolated from the gut of Atractomorpha sinensis. Bioresource Technology102(3): 3575-3577.
-
Yurchenko AN, Smetanina OF, Khudyakova YV, Kirichuk NN, Yurchenko EA, et al. (2013) Metabolites of the marine isolate of the fungus Curvularia inaequalis. Chemistry of Natural Compounds 49(1).
-
Garcia A, Rhoden SA, Bernardi Wenzel J, Orlandelli RC, Azevedo JL, et al. (2012) Antimicrobial activity of crude extracts of endophytic fungi isolated from medicinal plant Sapindus saponaria L. Journal of Applied Pharmaceutical Science 2(10): 35-40.
-
Rhoden SA, Garcia A, Bongiorno VA, Azevedo JL, Pamphile JA (2012) Antimicrobial activity of crude extracts of endophytic fungi isolated from medicinal plant Trichilia elegans A Juss. Journal of Applied Pharmaceutical Science 2(08): 57-59.
-
Nisa J, Kamili AN, Nawchoo IA, Shafi S, Shameem N, et al. (2015) Fungal endophytes as prolific source of phytochemicals and other bioactive natural products: A review. Microbial Pathogenesis 82: 50-59.
-
Barreto CF, Cavasin GM, Silva HHG, Silva IG (2006) Study of morphohistological changes in Aedes aegypti larvae (Diptera, culicidae) subjected to crude ethanol extract of Sapindus saponaria Lin (SAPINDACEAE). Revista de Patologia Tropical 35: 37-57.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework