Some Differences in Subspecies Morphology of Zanclorhynchus spinifer (Pisces: Zanclorhynchidae)
Additional data, calculation and statistics results of diagnostic parameters of the four subspecies of Zanclorhynchus spinifer Günther, 1880 are presented. Living for a long time in mostly geographically isolated populations within Antarctic Circumpolar Current (Marion-Crozet district, Kerguelen-Heard district, Macquarie Island, and Ridge of Hercules) leads to radiation into subspecies with significant morphological differences. The diagnostic importance of body depth, antedorsal distance, depth of gill opening, position of paired fins, proportions of the gill opening, heights of dorsal fin spines, and index of arming is confirmed. Of the 66 calculated pairs of parameters, 47 of them belongs to a different sets with statistical significance p<0.05. Antarctic horsefish differs from geographically closest Armed horsefish in maximum body depth, body depth at the origin of the anal fin, antedorsal distance, and index of arming with p-values lower than 0.000001. The index of arming also differentiates with p<0.001 Antarctic horsefish and Macquarian horsefish, Hercules horsefish and Armed horsefish, Macquarian horsefish and Armed horsefish. The difference in the sum of the lengths (heights) of the second, the third, and the fourth spines of first dorsal fin between all subspecies have statistical significance p<0.012. Differences in longitudinal orbit diameter have p-value <0.0001 in pairs Antarctic horsefish and Armed horsefish, Antarctic horsefish and Hercules horsefish. Listed parameters are sufficient for the identification of taxa.
Introduction
Family Zanclorhynchidae Andriashev, 1993 compares of two genera: Alertichthys Moreland, 1960 and Zanclorhynchus Günther, 1880 [1]. Both Zanclorhynchus species inhabit island shelves and seamounts in Indian and Pacific sectors of the Southern Ocean: Z. chereshnevi Balushkin et Zhukov, 2016 [2, 3] was caught in the waters of Marion- Crozet zoogeographic district, Z. spinifer Günther, 1880 [1] is distributed much wider – this species was described from the waters off Kerguelen Islands [4], Macquarie Island [5], Crozet Islands [6], Heard and McDonald Islands [6], Ob’ and Lena seamounts (Conrad Rise) [6], Prince Edward Islands [7], and Ridge of Hercules (Pacific-Antarctic Ridge) [8]. All these locations are within the Antarctic Circumpolar Current, which high velocity successfully isolate western localities from the others downstream. For example, for a horsefishes, Marion-Crozet district is fully isolated from Kerguelen-Heard district ~ 1500 km to the East. Benthic fishes, which prefer to walk slowly on four fins on the bottom rather than to swim [9], couldn’t overcome that distance upstream. From the other side horsefish from Crozet may occasionally reach the Kerguelen with the floating islands of macroalgae, so-called kelps. It is not surprising that fishes of different populations in such conditions were radiated into the rank of subspecies. Armed horsefish Z. spinifer armatus Zhukov, 2019 is the westernmost subspecies, ~ 1500 km downstream is distributed Antarctic horsefish Z. spinifer spinifer Günther, 1880 within the Kerguelen Plateau, than after ~ 5300 km to the east lies Macquarie Island with Macquarian horsefish Z. spinifer macquariensis Zhukov, 2019 [10], and at last ~ 4000 km eastward distributed the easternmost subspecies Hercules horsefish Z. spinifer heracleus [8].
In 2020, the impossibility of using the parameter of the distance between dorsal fins to determine the subspecies was shown. Despite the high statistical significance (p-value 0.000086 between Armed horsefish and Antarctic horsefish) it is impossible to use it because of individual variability [11]. Thus, it became necessary to confirm the operability of the main diagnostic parameters. An explanation of the measurement of the lengths of head and fin spines is given due to their high importance in diagnosis.
Materials and Methods
Four subspecies of Zanclorhynchus spinifer Günther, 1880 with total amount of 167 specimens from the collection of Zoological Institute RAS were examined from following locations: Z. spinifer spinifer (56 specimens): ZIN 39410 – 58 mm TL_, 48 mm SL, RV _Ob’, st. 121, near Kerguelen Island, 49°39′S-70°43′E, 140 m depth, 20 May 1956, coll. Andriashev AP and Tokarev AK; ZIN 40508 – 10 specimens, 123−270 mm TL, 96−220 mm SL, RV Skif, cruise 3, st. 1125/214, Kerguelen Island, 48°22′S-68°57′E, 131 m depth, 11 Jan. 1971, coll. Pushkin AF; ZIN 45673 – 2 specimens, 245 and 280 mm, 194 and 231 mm SL, RV Skif, tr. 377, Kerguelen Island, 19 Jan. 1970, coll. Kononov NV; ZIN 45684 – 2 specimens, 134 and 174 mm TL, 105 and 138 mm SL, RV Skif, cruise 3, tr. 38, о. Kerguelen Island, 48°57’S-67°27’E, 195– 207 m depth, 10 Dec. 1970, coll. Pushkin AF; ZIN 45685 – 142 mm TL, 114 mm SL, RV Skif, cruise 3, tr. 44, Kerguelen Island, 49°51’S-70°31’E, 220 m depth, 12 Dec. 1970, coll. Pushkin AF; ZIN 45686 – 155 mm TL, 124 mm SL, RV Skif, cruise 3, tr. 54, Kerguelen Island, 49°55’S-70°33’E, 215–225 m depth, 13 Dec. 1970, coll. Pushkin AF; ZIN 45687 – 228 mm TL, 185 mm SL, RV Skif, cruise 3, tr. 83, Kerguelen Island, 47°52’S-70°44’E, 153 m depth, 22 Dec. 1970, coll. Pushkin AF; ZIN 45688 – 221 mm TL, 181 mm SL, RV Skif, cruise 3, tr. 97, Kerguelen Island, 48°01’S-70°27’E, 146 m depth, 25 Dec. 1970, coll. Pushkin AF; ZIN 45689 – 216 mm TL, 178 mm SL, RV Skif, cruise 3, tr. 104, Kerguelen Island,
48°00’S-70°31’E, 148 m depth, 26 Dec. 1970, coll. Pushkin AF; ZIN 45690 – 146 mm TL, 116 mm SL, RV Skif, cruise 3, tr. 109, Kerguelen Island, 48°03’S-70°34’E, 147–149 m depth, 26 Dec. 1970, coll. Pushkin AF; ZIN 45691 – 26 specimens, 122–257 mm TL, 97–204 mm SL, RV Skif, cruise 3, st. 1072/161, tr. 113, Kerguelen Island, 47°25’S-68°36’E, 189 m depth, 02 Jan. 1971, coll. Pushkin AF; ZIN 45692 – 4 specimens, 143–161 mm TL, 112–130 mm SL, RV Skif, cruise 3, tr. 123, Kerguelen Island, 50°12’S-69°18’E, 197 m depth, 05 Jan. 1971, coll. Pushkin AF; ZIN 45693 – 247 mm TL, 201 mm SL, RV Skif, cruise 3, tr. 138, Kerguelen Island, 48°00′S-70°29′E, 152 m depth, 11 Jan. 1971, coll. Pushkin AF; ZIN 45694 –240 mm TL, 188 mm SL, RV Skif, cruise 3, tr. 139, Kerguelen Island, 47°11′S-70°16′E, 135 m depth, 11 Jan. 1971, coll. Pushkin AF; ZIN 45695 – 261 mm TL, 206 mm SL_, RV _Skif, cruise 3, tr. 140, Kerguelen Island, 47°59’S-70°16’E, 150 m depth, 11 Jan. 1971, coll. Pushkin AF; ZIN 45696 – 195 mm TL_, 156 mm SL, RV _Skif, cruise 3, tr. 141, Kerguelen Island, 48°21’S-70°02’E, 130 m depth, 11 Jan. 1971, coll. Pushkin AF; ZIN 45697 – 91 mm TL_, 73 mm SL, RV _Skif, cruise 3, tr. 162, Kerguelen Island, 48°22’S-70°01’E, 134–130 m depth, 15 Jan. 1971, coll. Pushkin AF; ZIN 45699 –122 mm TL_, 95 mm SL, RV _Skif, cruise 5, tr. 47, North off Kerguelen Island, 14 Feb. 1972, coll. Pushkin AF; Z. spinifer armatus (90 specimens): ZIN 56039 (Holotype) – 210 mm TL, 167 mm SL, FT Aelita, st. 17, Prince Edward Islands, 46°55′S-37°58′E, 170 m depth, 18 Jan. 1968, bottom trawl, coll. Karpenko AI and Volya GS; ZIN 40248 (Paratypes) – 26 specimens, 130– 205 mm TL_, 102–159 mm SL , RV _Skif, cruise 3, st. 944/33, tr. 18, Crozet Islands, 45°51’S-49°47’E, 235–260 m depth, 04 Dec. 1970, coll. Pushkin AF; ZIN 45541 (Paratypes) – 3 specimens, 156−212 mm TL, 124−172 mm SL, FT Chatyr- Dag, tr. 140, North off Crozet Island, 185 m depth, 17 Mar. 1973, coll. Tankevitch PB; ZIN 45682 (Paratypes) – 2 specimens, 101 and 131 mm TL, 79 and 103 mm SL, RV Skif, cruise 3, tr. 14, Crozet Islands, 46°13’S-49°34’E, 250–260 m depth, 04 Dec. 1970, coll. Pushkin AF; ZIN 45683 (Paratypes) – 248 mm TL_, 202 mm SL, RV _Skif, cruise 3, tr. 24, Crozet Islands, 45°43’S-50°15’E, 173 m depth, 05 Dec. 1970, coll. Pushkin AF; ZIN 45700 (Paratypes) – 2 specimens, 77–163 mm TL_, 64–129 mm SL, FT Aelita, Marion Island, 46°55’S-37°58’E, 170 m depth, 18 Jan. 1968, coll. Orlov VN; ZIN 56040 (Paratypes) –133 mm TL, 102 mm SL, FT Aelita, Prince Edward Islands, st. 17, 46°55′S-37°58′E, 170 m depth, 18 Jan. 1968, bottom trawl, coll. Karpenko AI and Volya GS; ZIN 45671 – 5 specimens, 132−186 mm TL, 103−153 mm SL, RV _Skif, cruise 3, st. 934/23, tr. 8, Crozet Islands, 46°15’S-49°31’E, 03 Dec. 1970, coll. Pushkin AF; ZIN 45672 – 6 specimens, 155−190 mm TL, 123−154 mm SL, RV Skif, cruise 3, st. 942/31, tr. 16, Crozet Islands, 45°56’S-49°28’E,310 m depth, 04 Dec. 1970, coll. Pushkin AF; ZIN 45675 – 3 specimens, 143−268 mm TL, 113−213 mm SL, FT Aelita, st. 41, Crozet Islands, 45°52’D-49°54’E, 230 m depth, 25 Jan. 1968, coll. Karpenko AI and Volya GS; ZIN
45676 – 118 mm TL_, 89 mm SL, RV _Skif, cruise 3, st. 917/6, tr. 1, Crozet Islands, 46°43›S-51°35›E, 375 m depth, 30 Nov. 1970, coll. Pushkin AF; ZIN 45677 – 122 mm TL_, 94 mm SL, RV _Skif, cruise 3, st. 920/9, tr. 4, Crozet Islands, 46°43›S-51°19›E, 320 m depth, 30 Nov. 1970, coll. Pushkin AF; ZIN 45678 – 176 mm TL_, 140 mm SL, RV _Skif, cruise 3, tr. 5, Crozet Islands, 46°38’S-51°03’E, 320 m depth, 01 Dec. 1970, coll. Pushkin AF; ZIN 45679 – 145 mm TL_, 112 mm SL, RV _Skif, cruise 3, st. 922/11, tr. 6, Crozet Islands, 46°35’S-50°42’E, 410 m depth, 01 Dec. 1970, coll. Pushkin AF; ZIN 45680 – 18 specimens, 146−186 mm TL_, 114−145 mm SL, RV _Skif, cruise 3, tr. 10, Crozet Islands, 46°02’S-49°24’E, 255−270 m depth, 03 Dec. 1970, coll. Pushkin AF; ZIN 45681 – 204 mm TL_, 166 mm SL, RV _Skif, cruise 3, tr. 13, Crozet Islands, 45°58’S-49°38’E, 254 m depth, 04 Dec. 1970, coll. Pushkin AF; ZIN 45698 – 2 specimens, 141 and 153 mm TL_, 112 and 124 mm SL, RV _Skif, cruise 3, Crozet Islands, coll. Pushkin AF; ZIN 56301 – 14 specimens, 136−187 mm TL_, 105−147 mm SL, RV _Skif, cruise 3, st. 944/33, tr. 18, Crozet Islands, 45°51’S-49°47’E, 235–260 m depth, 04 Dec. 1970, coll. Pushkin AF; ZIN 56516 – 181 mm TL, 143 mm SL, RV Skif, cruise 3, st. 923/12, tr. 7, Crozet Islands, 01 Dec. 1970, coll. Pushkin AF; Z. spinifer heracleus (7 specimens): ZIN 56110 (Holotype) – 140 mm TL, 109 mm SL, SSS Mys Tikhiy, tr. 57, Ridge of Hercules, Southern Bank, 53°44’S-140°19’W, 260–380 m depth, 05 Oct. 1977, coll. Pavlova TP; ZIN 45701 (Paratypes) – 2 specimens, 126 and 127 mm TL, 101 and 102 mm SL, FRV Gerakl, tr. 114, Ridge of Hercules, Central (Umanova) Bank, 53°33’S-140°41’W, 170–260 m depth, 20 Dec. 1975, coll. Pavlova TP; ZIN 45702 (Paratypes) – 2 specimens, 127 and 136 mm TL, 100 and 108 mm SL, SSS Mys Tikhiy, tr. 57, Ridge of Hercules, Southern Bank, 53°44’S-140°19’W, 260–380 m depth, 05 Oct. 1977, coll. Pavlova TP; ZIN 45706 (Paratype) – 260 mm TL, 215 mm SL, FRV Kamenskoe, Ridge of Hercules, Central (Umanova) Bank, 53°32’S-140°40’W, 250 m depth, 05 Aug. 1978, coll. TINRO; ZIN 45703 – 274 mm TL, 224 mm SL, SSS Mys Tikhiy, tr. 29, Ridge of Hercules, Central (Umanova) Bank, 53°30’S-140°42’W, 270–400 m depth, 26 Sep. 1977, coll. Pavlova TP; Z. spinifer macquariensis (14 specimens): ZIN 56401 (Holotype) – 230 mm TL, 180 mm SL, SRV Dmitriy Mendeleev, Macquarie Island, Buckles Bay, 21–27 m depth, cruise 16, fishing rod, 22 Jan. 1976, coll. Andriashev AP and Prirodina VP; ZIN 45705 (Paratype) – 3 specimens, 91–104 mm TL, 72–82 mm SL, SRV Dmitriy Mendeleev, cruise 16, tr. 1293, South off Macquarie Island, 54°53′S-158°47′E, 80–84 m depth, 25 Jan. 1976, coll. Andriashev AP and Prirodina VP; ZIN 56402 (Paratypes) – 2 specimens, 229 and 259 mm TL, 182 and 204 mm SL, SRV Dmitriy Mendeleev, Macquarie Island, Buckles Bay, 21–27 m depth, cruise 16, fishing rod, 22 Jan. 1976, coll. Andriashev AP and Prirodina VP; ZIN 45704 – 6 specimens, 206–265 mm TL, 161–210 mm SL, SRV Dmitriy Mendeleev, Macquarie Island, Buckles Bay, 21–27 m depth, cruise 16, fishing rod, 22 Jan. 1976, coll. Andriashev AP and Prirodina VP; ZIN 56403 – 2 specimens, 57 and 60 mm TL, 46 and 48 mm SL, SRV Dmitriy Mendeleev, cruise 16, tr. 1293 [10, 11] South off Macquarie Island, 54°53′S-158°47′E, 80–84 m depth, 25 Jan. 1976, coll. Andriashev AP and Prirodina VP. Standard measurements followed Hubbs, et al. [12], were made with a vernier caliper with an accuracy of 0.1 mm. Every head spine belongs to the segment of the sensory system [2]. These spines are measured from the tip to the point of the sharpest bend towards the base of the bone. Posttemporal, suborbital, lacrimal, and supraorbital spines are measured from above, cleithrum spine anteriorly. Spines of the first dorsal fin were measured from the tip to the upper contour of the body anteriorly. Formula of the arming index is: _I_ar = _lsT_II + 0.1_hD_12–4 [8] where _lsT_II is a length of the posttemporal spine, _hD_12–4 – a sum of the heights of the second, third, and fourth spine of the first dorsal fin. Length of the posttemporal spine calculated as the mean of the spines lengths from both sides. In case of a broken spine, its length was taken as mean in subspecies sample.
All samples were tested for normal distribution. After correcting the outliers the distributions are normal, p>0.20 using Kolmogorov–Smirnov test. Outliers changed to the nearest μ±2ϭ, where μ – mean and ϭ – standart deviation. The number of outliers in any sample did not exceed 1%. The samples were compared using the Student’s t-test in Statistica 8.0 software package.
Results
Results of the measurements and calculations are shown in Table 1.
| Parameter | Z. spinifer armatus | Z. spinifer spinifer | Z. spinifer heracleus | Z. spinifer macquariensis |
|---|---|---|---|---|
| in % SL | ||||
| h | 15.9–23.0 (19.7) | 18.3–24.5 (21.2) | 17.2–20.6 (19.0) | 16.0-21.3 (19.4) |
| H | 23.9–32.8 (27.5) | 25.7–32.1 (29.3) | 26.5–29.9 (27.8) | 27.3-32.4 (29.8) |
| aD | 29.9–39.5 (34.4) | 28.8–38.0 (32.6) | 33.7–38.7 (35.8) | 30.5-38.5 (35.4) |
| hBr | 6.1–11.4 (8.5) | 6.0–10.6 (8.2) | 7.0–8.3 (7.7) | 7.7-11.3 (9.6) |
| P–V | 13.8–20.1 (17.0) | 14.3–19.8 (17.3) | 13.4–17.8 (15.6) | 12.7-21.0 (17.9) |
| c | 32.2–39.4 (35.3) | 31.4–38.9 (34.5) | 36.4–39.9 (38.1) | 33.4-38.7 (36.6) |
| ao | 10.9–16.8 (13.4) | 11.0–16.1 (12.9) | 12.8–16.1 (14.5) | 13.0-16.2 (14.3) |
| o | 7.6–11.3 (9.2) | 6.5–12.7 (8.4) | 7.7–11.2 (9.7) | 7.5-11.9 (9.1) |
| hD1 1 | 6.6–20.2 (12.2) | 6.3–14.7 (11.2) | 7.4–11.4 (8.9) | 6.8-13.7 (10.6) |
| hD1 2–4 | 59.8–109.7 (88.1) | 62.9–98.9 (82.6) | 52.4–82.2 (64.1) | 59.1-88.6 (73.9) |
| Index | ||||
| I ar | 16.7–29.7 (23.2) | 11.3–21.0 (16.1) | 12.5–20.1 (16.8) | 9.6–24.7 (18.5) |
Table 1: Morphometric parameters of four subspecies _Zanclorhynchus spinifer_: body depth at the origin of the anal fin (_h_), ma
Table 1: Morphometric parameters of four subspecies Zanclorhynchus spinifer: body depth at the origin of the anal fin (h), maximal body depth (H), antedorsal distance (aD), depth of gill opening (hBr), distance between paired fins (P–V), length of the head (c), snout length (ao), longitudinal orbit diameter (o), length (height) of the first spine of first dorsal fin (hD_11), sum of the lengths (heights) of the second, the third, and the fourth spines of first dorsal fin (_hD_12–4), index of arming (_Iar).
Diagnostic [10] and additional parameters of subspecies mostly differs with high statistical significance. The statistical analysis results are shown below:
- Body depth at the origin of the anal fin (Figure 1A): Samples h_spinife and _h_armatus belong to a different sets (_t-value 6.83, p-value 0.000000), h_spinifer and _h_heracleus belong to a different sets (_t=4.25, p=0.000076), h_spinifer and _h_macquariensis belong to a different sets (_t=4.76, p=0. 000011), no statistically significant difference in samples h_heracleus and _h_macquariensis (_t=0.27, p=0.790087), no statistically significant difference in samples h_heracleus and _h_armatus (_t=1.31, p=0.193873), no statistically significant difference in samples h_armatus and _h_macquariensis (_t=1.18, p=0.239484).
- Maximal body depth (Figure 1B): Samples H_spinifer and _H_armatus belong to a different sets (_t=6.38, p=0.000000), samples H_spinifer and _H_heracleus belong to a different sets (_t=2.31, p=0.024088), samples H_heracleus and _H_macquariensis belong to a different sets (_t=2.95, p=0.008912), samples H_armatus and _H_macquariensis belong to a different sets (_t=4.27, p=0.000045), no statistically significant difference in samples H_spinifer and _H_macquariensis (_t=0.69, p=0.491915), no statistically significant difference in samples H_heracleus and _H_armatus (_t=0.51, p=0.609662).
- Antedorsal distance (Figure 1C): Samples aD_1spinifer and _aD_1armatus belong to a different sets (_t=6.13, p=0.000000), samples aD_1spinifer and _aD_1heracleus belong to a different sets (_t=4.42, p=0.000043), samples aD_1spinifer and _aD_1macquariensis belong to a different sets (_t=4.24, p=0.000071), no statistically significant difference in samples aD_1heracleus and _aD_1armatus (_t=1.90, p=0.061029), no statistically significant difference in samples aD_1heracleus and _aD_1macquariensis (_t=0.64, p=0.533597), no statistically significant difference in samples aD_1armatus and _aD_1macquariensis (_t=1.20, p=0.231720).
- Depth of gill opening (Figure 1D): Samples hBr_spinifer and _hBr_macquariensis belong to a different sets (_t=4.77, p=0.000011), samples hBr_heracleus and _hBr_macquariensis belong to a different sets (_t=4.02, p=0.000895), samples hBr_armatus and _hBr_macquariensis belong to a different sets (_t=3.31, p=0.001301), no statistically significant difference in samples hBr_spinifer and _hBr_armatus (_t=1.65, p=0.101494), no statistically significant difference in samples hBr_spinifer and _hBr_heracleus (_t=1.39, p=0.169527), no statistically significant difference in samples hBr_heracleus and _hBr_armatus (_t=1.80, p=0.075109).
- Distance between paired fins (Figure 1E). Samples P– V_spinifer and _P–V_macquariensis belong to a different sets (_t=2.61, p=0.011306), samples P–V_spinifer and _P–V_heracleus belong to a different sets (_t=3.37, p=0.001322), samples P–V_heracleus and _P–V_macquariensis belong to a different sets (_t=3.80, p=0.001422), samples P–V_heracleus и _P–V_armatus belong to a different sets (_t=2.68, p=0.008529), samples P–V_armatus and _P–V_macquariensis belong to a different sets (_t=2.72, p=0.007480), no statistically significant difference in samples P–V_spinifer and _P–V_armatus (_t=0.70, p=0.488347).
- Length of the head (Figure 1F): Samples c_spinifer and _c_macquariensis belong to a different sets (_t=3.72, p=0.000420), samples c_spinifer and _c_armatus belong to a different sets (_t=2.78, p=0.006199), samples c_spinifer and _c_heracleus belong to a different sets (_t=6.02, p=0.000000), samples c_heracleus and _c_macquariensis belong to a different sets (_t=2.33, p=0.032161), samples c_heracleus and _c_armatus belong to a different sets (_t=4.52, p=0.000018), samples c_armatus and _c_macquariensis belong to a different sets (_t=2.17, p=0.032247).
- Snout length (Figure 1G): Samples ao_spinifer and _ao_macquariensis belong to a different sets (_t=4.46, p=0.000033), samples ao_spinifer and _ao_armatus belong to a different sets (_t=2.24, p=0.026591), samples ao_spinifer and _ao_heracleus belong to a different sets (_t=3.67, p=0.000511), samples ao_heracleus and _ao_armatus belong to a different sets (_t=2.39, p=0.018837), samples ao_armatus and _ao_macquariensis belong to a different sets (_t=2.86, p=0.005128), no statistically significant difference in samples ao_heracleus и _ao_macquariensis (_t=0.19, p=0,853760).
- Longitudinal orbit diameter (Figure 1H): Samples o_spinifer and _o_armatus belong to a different sets (_t=4.17, p=0.000046), samples o_spinifer and _o_heracleus belong to a different sets (_t=4.35, p=0.000053), samples o_heracleus and _o_armatus belong to a different sets (_t=2.72, p=0.007347), samples o_heracleus and _o_macquariensis belong to a different sets (_t=2.26, p=0.037155), no statistically significant difference in samples o_spinifer and _o_macquariensis (_t=1.27, p=0.208855), no statistically significant difference in samples o_armatus and _o_macquariensis (_t=0.92, p=0.358313).
- Length (height) of the first spine of first dorsal fin (Figure 1I): Samples hD_11spinifer and _hD_11armatus belong to a different sets (_t=2.38, p=0.018594), samples hD_11spinifer and _hD_11heracleus belong to a different sets (_t=2.75, p=0.007870), samples hD_11heracleus and _hD_11armatus belong to a different sets (_t=3.40, p=0.000994), samples hD_11heracleus and _hD_11macquariensis belong to a different sets (_t=2.36, p=0.030271), no statistically significant difference in samples hD_11spinifer and _hD_11macquariensis (_t=0.42, p=0.674648), no statistically significant difference in samples hD_11armatus and _hD_11macquariensis (_t=1.66, p=0.099587).
- Sum of the lengths (heights) of the second, the third, and the fourth spines of first dorsal fin (Figure 1J): Samples hD_12–4spinifer and _hD_12–4heracleus belong to a different sets (_t=5.78, p=0.000000), samples hD_12–4spinifer and _hD_12–4macquariensis belong to a different sets (_t=2.65, p=0.010077), samples hD_12–4spinifer and _hD_12–4armatus belong to a different sets (_t=3.36, p=0.000988), samples hD_12–4heracleus and _hD_12–4macquariensis belong to a different sets (_t=2.85, p=0.011038), samples _hD_12–4heracleus and _hD_12–
4armatus belong to a different sets (t=6.26, p=0.000000), samples hD_12–4armatus and _hD_12–4macquariensis belong to a different sets (_t=4.04, p=0.000107). • Index of arming (Figure 1K): Samples Iar_spinifer and _Iar_armatus belong to a different sets (_t=17.30, p=0.000000), samples Iar_spinifer and _Iar_macquariensis belong to a different sets (_t=6.16, p=0.000000), samples Iar_heracleus and _Iar_armatus belong to a different sets (_t=5.83, p=0.0000000), samples Iar_armatus and _Iar_macquariensis belong to a different sets (_t=3.86, p=0.000204), no statistically significant difference in samples Iar_spinifer and _Iar_heracleus (_t=0.96, p=0.341917), no statistically significant difference in samples Iar_heracleus and _Iar_macquariensis (_t=2.07, p=0.053833).
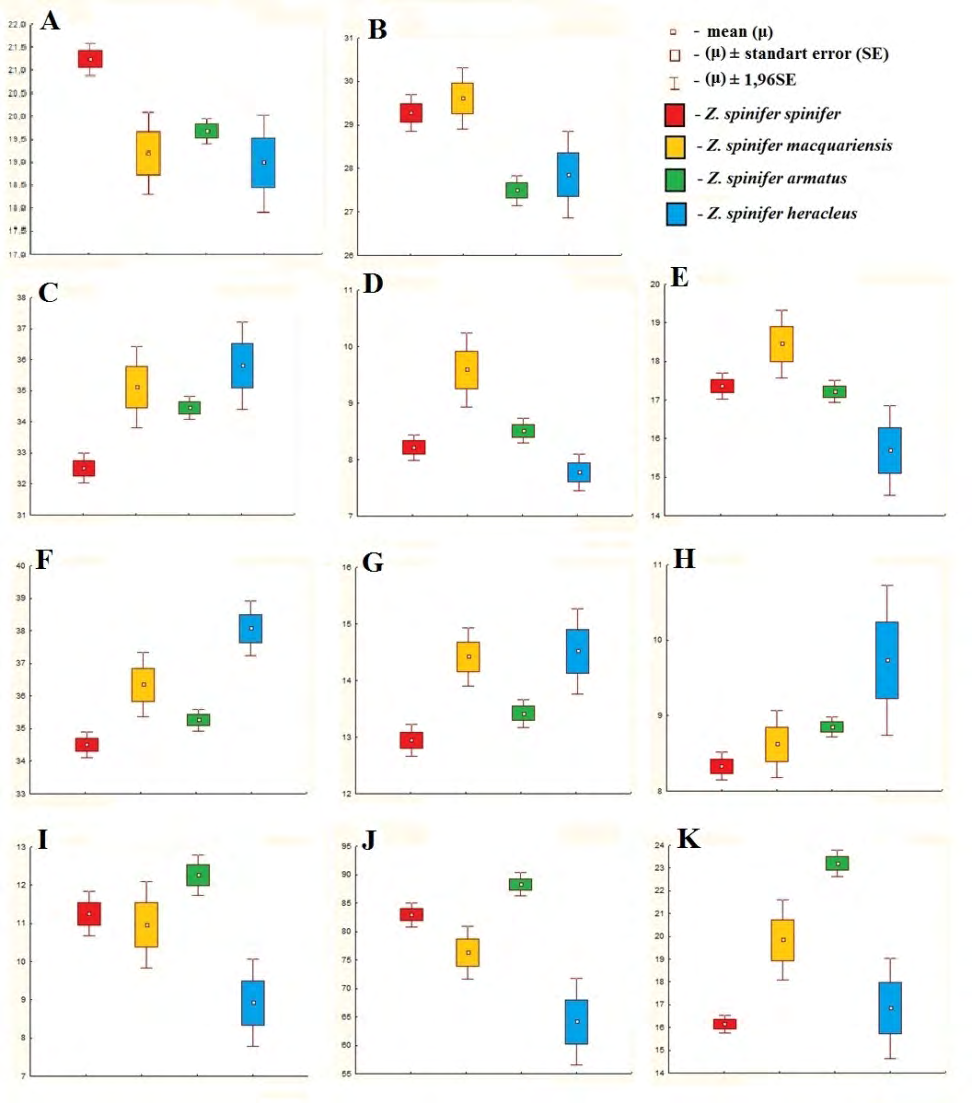
Figure 1: Parameter statistics: A – body depth at the origin of the anal fin, B – maximal body depth, C – antedorsal distance, D – depth of gill opening, E – distance between paired fins, F – length of the head, G – snout length, H – longitudinal orbit diameter, I – length (height) of the first spine of first dorsal fin, J – sum of the lengths (heights) of the second, the third, and the fourth spines of first dorsal fin, K – index of arming.
Discussion
There are no paleontological data on the time of the appearance of horsefishes in the Kerguelen zoogeographic subregion. The starting point may be the age of the lava on Marion Island of the Prince Edward Archipelago, the westernmost point of Zanclorhynchus distribution area. Thus, the archipelago could begin to form 500,000 years ago [13]. As mentioned above, it is difficult to imagine the colonizing of this archipelago from the Crozet Islands located to the East, downstream the Antarctic Circumpolar Current. Prince Edward is separated by depths of more than 3 km [14], insurmountable for these species under the condition of a strong countercurrent, even from the Del Cano Rise, located approximately in the middle between these archipelagos, where horsefishes are also noted.
It can be assumed that horsefishes entered the Kerguelen subregion not earlier than 500,000 years ago. Further expansion to the east allowed the horsefishes to create several practically isolated populations. Staying in such isolation for a long time allowed the populations to acquire a large number of differing morphometric parameters. However, the isolation time was not enough for the formation of the strong hiatuses using which the discussed four taxa could be simply identified.
Antarctic horsefish Zanclorhynchus spinifer spinifer have a smallest head in the group, what also leads to small snout and antedorsal distance, the body depth at the origin of the anal fin is the largest, but maximal body depth of this subspecies is close to Macquarian horsefish Zanclorhynchus spinifer macquariensis. Macquarian horsefish has the largest values of depth of gill opening and distance between paired fins. Armed horsefish Zanclorhynchus spinifer armatus have the greatest values in index of armory, maximal body depth is similar to that of Hercules horsefish Zanclorhynchus spinifer heracleus. These two subspecies form a group opposite to Macquarian and Antarctic horsefishes with higher bodies [15]. The heights of the anterior profile of the first dorsal fin are different in all subspecies, the highest of Armed horsefish and the lowest of Hercules horsefish. Using a series of the listed parameters helps to firmly determine subspecies.
Of the above measurements, the distance between paired fins and arming parameters are clearly adaptive apomorphies. A greater distance between paired fins than of the fish in sister family Cоngiopodidae is not a plesiomorphic [16], but an apomorphic [9] state. The pelvic girdle shifted back is more convenient for the way of locomotion formed in Zanclorhynchus genus, slow movement on four fins along the bottom. It is clear that being armed is an adaptation feature as a defense against predators. The development of spines on the head and the first spines of the dorsal fin led to the fact that the only source of danger are predators of the highest trophic level with the most developed fishing techniques: cormorants, penguins, some mammals of the Otariidae family [17, 18, 19]. The greatest index of arming have Armed horsefish (23.2% SL mean) and Macquarian horsefish (18.5). It remains unclear why Zanclorhynchus spinifer spinifer (16.1) is so much less armed than Z. spinifer macquariensis. In both cases, these fishes are food for Pigoscelis papua [17, 20]. Perhaps, fishes from Macquarie Island have better developed cranial spines due to greater pressure from other predators – Phocarctos hookeri and Eudyptes chrysocome [21, 22]. Or this can be explained by different defense strategy: Z. spinifer spinifer have a well-developed spines of the dorsal fin, 82.6% SL mean versus 73.9% SL of Z. spinifer macquariensis. The lack of pressure of the predators explains well the low index of arming in Hercules horsefish (16.8). Here, the minimum depth in the habitat is 170 m [23], and the nearest land is ~ 2300 km south (Siple Island) or ~ 2750 km west (Chatham Islands), what sharply reduces the ability of birds and mammals to prey in this region.
Funding
The study was supported by the Russian State Research Program number AAAA-A19-119020790033-9.
Conflict of Interest
The author declare that he have no conflict of interest.
References
-
Voskoboinikova OS, Zhukov MYu (2021) Comparative osteology of the superfamily Congiopodoidea (Pisces: Scorpaenoidei). Zoomorphology 140: 373-385.
-
Balushkin AV, Zhukov MYu (2016) Polytypy of the genus Zanclorhynchus (Zanclorhynchinae: Congiopodidae): description of the new species _Z. chereshnevi_ sp. n. from the Indian sector of the Southern Ocean. J Ichthyol 56: 791-798.
-
Zhukov MYu (2020) First record of _Zanclorhynchus_ _chereshnevi_ (Congiopodidae) off the Crozet Archipelago, Southern Ocean. J Ichthyol 60: 786-790.
-
Günther A (1880) Report on the shore fishes. Scientific results of the voyage of H.M.S. Challenger during the years 1873–1876. Challenger Reports. Zoology 1(6): 1-82.
-
Waite ER (1916) Fishes. Australasian Antarctic Expedition. Scientific reports. Series C 3(1): 1-93.
-
Meisner EE, Kratkii VE, Tot VS (1977) Species composition of ichthyofauna of the island shelves of the Indian Sector of the Antarctic. Proceedings of VNIRO 120: 55-62.
-
Permitin YuE (1987) To the study of the fauna and distribution of benthic fishes of the Antarctic. Biological resources of the Arctic and Antarctic. Nauka, Moscow: 258-296.
-
Zhukov MYu, Balushkin AV (2018) Description of a New Subspecies _Zanclorhynchus spinifer heracleus_ subsp. nov. from the Ridge of Hercules (Pacific-Antarctic Ridge). J Ichthyol 58: 104-112.
-
Andriashev AP (1993) Quadrupedal locomotion of Kerguelen Antarctic horsefish _Zanclorhynchus spinifer_ (Scorpaeniformes, Congiopodidae). Zoologicheskiy Zhurnal 72(9): 130-136.
-
Zhukov MYu (2019) Two new subspecies of Antarctic horsefish (_Zanclorhynchus_, Scorpaeniformes: Congiopodidae) from Indian Sector of Southern Ocean. Proceedings of the Zoological Institute RAS 323(4): 541- 557.
-
Zhukov MYu (2020) Statistical analysis of the interdorsal distances of subspecies _Zanclorhynchus spinifer_ Günther 1880. International scientific conference “Scientific forum: medicine, biology and chemistry 34: 5-10.
-
Hubbs CL, Lagler KF (1947) Fishes of the Great Lakes Region. Bulletin Cranbrook Institute of Science (Bloomfield Hills, Michigan) 26: 186.
-
McDougall I (1971) Geochronology. Marion and Prince Edward Islands. Balkema, Cape Town, South Africa, pp: 72-77.
-
Ansorge IJ, Froneman PW, Pakhomov EA, Lutjeharms JRE, Perissinotto R, et al. (1999) Physical-biological coupling in the waters surrounding the Prince Edward Islands (Southern Ocean). Polar Biology 21: 135-145.
-
Zhukov MYu (2020) Determination of the two groups different in body profile within the species _Zanclorhynchus_ _spinifer_ (Scorpaeniformes: Congiopodidae). International scientific conference Biodiversity protection and environmental problems of nature management 1: 113-116.
-
Moreland JM (1960) A new genus and species of congiopodid fish from southern New Zealand. Records of The Dominion Museum 3: 241-246.
-
Lescroel A, Bost CA (2004) Spatial and temporal variation in the diet of the Gentoo penguin (_Pygoscelis_ _papua_) at Kerguelen Islands. Polar Bioljgy 27: 206-216.
-
Duhamel G, Gasco N, Davaine P (2005) Poissons des îles Kerguelen et Crozet, guide régional de l’océan Austral. Patrimoines naturels 63: 419.
-
Green K, Burton HR, Williams R (1989) The diet of Antarctic fur seals _Arctocephalus gazella_ (Peters) during the breeding season at Heard Island. Antarctic Science 1: 317-324.
-
Robinson SA, Hindell MA (1996) Foraging ecology of gentoo penguins _Pygoscelis papua_ at Macquarie Island during the period of chick care. IBIS 138(4): 722-731.
-
McCahon CR, Holley D, Robinson S (1999) The diet of itinerant male Hooker’s sea lions, _Phocarctos hookeri_, at sub-Antarctic Macquarie Island. Wildlife Research 26: 839-846.
-
Hindell MA (1988) The diet of the Rockhopper penguin Eudyptes chrysocome at Macquarie island. Emu 88: 227- 233.
-
Boldyrev VZ, Darnitskii VB (1991) Specific distribution of fishes o the submerged mountains of the Eltanin Fracture Zone. In: Biological Resources of the Thalassobathyal Zone of the World Ocean. VNIRO, Moscow, Russia, pp: 258-285.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework