Vibrio Species of Cultured Marine Fishes Importance: A Comprehensive Review
Aquaculture is the fastest-growing high-protein resource market worldwide. Increased production in aquaculture brings along epidemics, which negatively affect the profitability and sustainability of production. In particular, vibriosis is responsible for the important diseases that can cause significant economic losses in cultured shellfish, shrimp, and fish. It may be extremely damaging to aquaculture industries, and preventative and treatment techniques are typically highly costly. Besides, the studies are limited for virulence, toxin genes, effects on host cells, and socio-economic effects of species that cause vibriosis in aquaculture. In addition, Vibrio species includes important pathogens that can especially induce zoonotic diseases, which are significant for public health. This review emphasizes various updated basic information about aquaculture disease including the etiology, epidemiology, pathogenesis, clinical signs, pathological findings, diagnosis, transmission, risk factors, and control measures. The study will also highlight any possible knowledge gaps in the literature, which will serve as a foundation for future research goals.
Introduction
Aquaculture is the fastest-growing high-protein resource market worldwide. It promotes food values and alleviates hunger in certain developing countries, contributing to global economic prosperity and social equilibrium. Aquaculture is described as the development of aquatic organisms along with finfish and shellfish by communities, individuals, or companies through strategies such as drugs, feed, regulated farming, and containment. The aquaculture industry is also important for the human diets that provide more than 15% of animal protein and other essential micronutrients, including calcium, iron, vitamin A, and zinc. However, infections in aquaculture are known to have an economic and social impact worldwide [1]. It has been reported that 66.7% of infections recorded in groupers Epinephelus spp. were vibriosis, which affects all stages of development and can kill up to half of the fish [2]. Several Vibrionaceae species have been connected to health issues in humans and aquatic animals [1]. In 1965, Veron named non-enteric, fermentative, ram-negative rod-shaped bacteria Vibrionaceae, which are motile and oxidase-positive using polar flagella. A new batch has been made to differentiate Vibrionaceae from Enterobacteriaceae. According to Bergey’s Manual of Determinative Bacteriology, Vibrioneacae is a member of the Gammaproteobacteria class that divides into four main genera including Vibrio, Aeromonas, Photobacterium, and Plesiomonas. However, two new genera, Listonella and Shewanella, and a new family Aeromonadaceae have been established developing sequencing analysis techniques in recent years [3].
Vibrio Species
Vibrio species have importance in both historical and contemporary periods. Vibrio cholerae is the causative bacteria of Asiatic cholera, which has been a serious problem for humanity for centuries [4, 5, 6, 7]. The bacteria was first described and named by Italian Filippo Pacini in 1854. After that German scientist Robert Koch described it as a curved or comma-shaped bacteria in pure cultures and named them “Kommabacillus” [8, 9].
Several Vibrionaceae species are associated with infection in marine animals. In recent studies, V. parahaemolyticus, V. anguillarum (formerly named Listonella anguillarum), V. ordalii, V. salmonicida, V. vulnificus, V. alginolyticus, V. harveyi, V. owensii, and V. campbelli were described as the most common species affecting aquatic animals. In addition, some Photobacterium species such as Photobacterium damselae subsp. piscicida (formerly described by Janssen and Surgalla in 1968 and known as Pasteurella piscicida), P. damselae subsp. damselae (previously known as V. damselae), and P. toruni isolated from sand steenbras have been recently described. Vibrio species can be found in large amounts in marine coastal waters and most of the aquatic environments in the world. In addition, it has been found that they are placed in large amounts in marine creatures such as corals, fish, mollusks, sea grass, sponges, shrimp, and zooplankton [10].
Vibriosis
Vibriosis is an important disease that causes by different Vibrio species and fish losses in the wild and aquaculture. Vibrio ordalii, which is one of the bacteria in that group, is a gram-negative bacteria, curved rod with polar flagella. Vibrio salmonicida, which affects Atlantic salmonids (Salmo salar) and has cold water vibriosis, is a facultative anaerobe, motile and club-shaped bacterium. It has active polar flagella and shows a high degree of pleomorphism between strains when isolated from fish. Vibrio vulnificus, also known as Vibrio anguillicida, causes disease in cultivated Japanese eels (Anguilla japonica) [6, 8, 10].
Vibrio species geographically can cause a wide range of infections in wild and aquaculture. They are halophilic bacteria that can grow in the presence of 0.5-4% salt concentration. Bacteria are psychrophiles although their optimum growing temperature is 12-15°C. Among the Vibrio species that can be excreted with feces in fish, especially V. salmonicida can be isolated from the sediment in the pond even after seven months in infected farms. V. salmonicida is also described as the etiological agent of cold-water vibriosis affecting aquaculture in Norway. The agent emerged in 1977, first causing a large-scale economic loss infection in fish farms on the Norwegian island of Hitra in 1979. For this reason, the disease caused by the agent is also called Hitra disease. After that, the disease caused serious economic losses on the western and northern coasts of Norway. However, Vibrio- borne infections mostly occur in autumn, winter, or early spring, as well as affecting Atlantic salmonids, rainbow trout, silverfish, king salmon, and eels [10, 11].
Although Vibrio species infect mainly the muscle and skin of the fish, they can also be isolated from different tissues. The clinical findings primarily characterize necrosis and hemorrhages in the tissues where it is localized. Besides, infected animals can demonstrate extensive anemia and internal hemorrhages in visceral organs. The bacteria can be isolated from the blood of newly dead fish. Cataracts, cranial hemorrhages, and splenomegaly may happen in the infected small fish. The bacteria are abundant in the connective tissue of the gill, the digestive tract, and the pyloric stomach of salmonids [10, 11].
The typical non-hemolytic and grayish pigment-free colonies are defined as Vibrio species on blood agar. The bacteria can be isolated from the intestinal content of the fish that is experimentally infected. Hence, besides being isolated from healthy animals, it is possible to isolate them from marine sediments after being scattered with feces in infected animals. Among the Vibrio species, Vibrio salmonicida is serologically separated from Vibrio anguillarum that grow at 30°C on tryptic soy broth or brain-heart infusion agar including 1.0% NaCl. ELISA is used for rapid detection of the disease. Even though V. vulnificus experimentally infect mice, it was defined that the strain isolated from eels, which is called V. vulnificus biogroup 2, causes infection only in eels [8].
There are many antibiotic options in the treatment of vibriosis. The treatment strategies primarily use for aquaculture instead of the natural environment. Florfenicol, oxanilic acid, and quinolone are common antibiotics used in the treatment of disease in aquaculture. Marine sediments both close to and far from aquacultural activities contain residual antibiotics and may be sensitive to the development of antibiotic resistance due to heavily used antibiotics. It is very important to monitor the water and feed quality in aquaculture, reduce the stress factors, and take effective biosecurity measures in preventing infection. The study of vaccine development to protect against vibriosis still needs further studies [10, 11].
Photobacterium Species
The genus of Photobacterium, which was first discovered by Beijerinck in 1889, is a member of the Vibrionaceae family in the class Gammaproteobacteria. They are gram- negative, facultative aerobic, thick, club-shaped, having a 0.8–1.3 µm diameter and 1.8-2.4 µm length, with incubation at 18–25°C for 48-72h. Although motile by one or three unsheathed polar flagella, some of them do not. The member of Photobacterium species is mostly oxidase and catalase- positive, nitrite produced from nitrate. Some strains grow more easily on the media containing amino acids and sodium chloride (0.5–3.0%, w/v). Although Photobacterium species produce acid and gas from glucose, they do not utilize lactose. Involuted forms may occur under adverse conditions of cultivation and old cultures. While most strains can grow in seawater-based environments with D-glucose and NH4Cl, some strains require L-methionine [3, 12, 13]. Another characteristic of bacteria in these species is that they produce polyunsaturated fatty acids, such as lipase, asparaginase, esterase, and antimicrobial components for adaptation to cold. Hence, they can be used as biosensors for food monitoring. In addition, they produce some unsaturated fatty acids that are used in the pharmaceutical industry, food additives, and supplements. They can also be used in different areas, such as in the production of detergent, food, biological drugs, biodiesel, anticancer drugs, and in various enzymes, such as cold-resistant lipase, esterase, and asparaginase [14, 15].
The common feature of Vibrio species is that their metabolites mostly show antimicrobial properties. Similarly, Photobacterium species also exhibit antibacterial properties via various metabolites (e.g., pyrrothine-homolycin, and korormicin) against several bacteria, such as Escherichia coli, Staphylococcus aureus, and V. parahaemolyticus, which are clinically important pathogens [16].
Genomically, it has been determined that the genetic structure of Photobacterium species is of importance, which includes two chromosomes, one big and the other small, in the large chromosome. In addition, bacteria show the presence of different plasmids. It has been reported that these plasmids are associated with properties of conjugation, antibiotic resistance, and virulence in Photobacterium species, while some encode toxins [17, 18]. It has also been found to be responsible for hemolysis and virulence in fish and mice. A different plasmid (Phdp) has been observed to encode an exotoxin that affects the macrophages and neutrophils of fish, and this plasmid is also involved in the transfer of virulent genes and antibiotic resistance (e.g., beta-lactamase and chloramphenicol resistance) to V. Cholerae [17, 19].
Photobacterium Damselea Subsp. Damselae (Photobacteriosis)
Photobacterium damselea subsp. damselae is a species of Photobacterium that can cause pathogenesis in fish and marine mammals. The bacterium was firstly reported in white sea bass. Extracellular structures secreted by Photobacterium species have an important place in the pathogenesis of the infection. The extracellular products containing proteases, hemolysins, and iron-degrading proteins associated with siderophores, enable invasion and colonization in the hosts besides facilitating nutrient uptake. Photobacterium damselea subsp. damselae has a phospholipase toxin with hemolytic and cytotoxic activities. It also produces a thermostable extracellular cytotoxin of 69 kDa named damselysin that is a phospholipase D enzyme, which acts against the structure of sphingomyelin in the erythrocyte membrane and shows hemolysis activity in many fish species [20, 21]. It has been determined that the Photobacterium psicicida also causes infections with high mortality in fish. The polysaccharide capsular structure causing serum resistance in fish is important in the pathogenesis for both species, whereas this structure increases the mortality of the bacteria. The combination of phospholipase effect, cytotoxicity, and hemolytic activities cause damage to the infected cells and the development of the disease by aiding bacterial invasion into the host tissue. P. psicicida produces an apoptosis-inducing exotoxin (AIP56), which is encoded by a 56kDa plasmid. The exotoxin that activates apoptosis but inhibits the host immune response, synthesized by virulent strains increases apoptotic activity, especially in sea bass. Another important virulence factor is the production of iron-binding siderophores that can bind to iron with high affinity in host cells. These siderophores are similar to the pathogenicity islands of Yersinia species [22].
In addition to the pathogenic character of the bacteria belonging to the genus Photobacterium, some species are symbiotic microorganisms that reside outside the host in marine fish. Although fish provide nutrients and oxygen to fulfill the vital needs of the bacteria, the host structures where the bacteria are localized serve as the light-producing organ. For example, luciferin, which is synthesized by bacteria, shows luminescence properties, and this helps animals in their behaviors such as mating, getting away from prey, and catching prey. At the same time, bacteria that grow on the host tissue localize both in sea water and in other sea creatures sharing the same environment. Photobacterium species can live in various ecological environments, such as various animal tissues, surfaces of fish, and intestines [23].
Recently, Photobacterium species have been identified in frozen ready-to-eat seafood. These Photobacterium species have been reported to be resistant to freezing and posing a human health hazard [24].
Photobacterium has similar characteristics to Vibrio as a member of the Vibrionaceae family. There is a controversy regarding whether some species should be included in the genus Phobacterium or Vibrio. However, in recent studies, it has been accepted that V. damselea, which was previously considered to be in the genus Vibrio, is included in the genus Photobacterium. P. damselea species is classified under two subspecies as P. damselea subsp. piscicida and P.
damselea subsp. damselea. Apart from this, P. leiognathi, P. phosphoreum, and P. profundum are also recently included in the genus Photobacterium [25].
P. damselea subsp. damselea that can live in high salt concentrations (12%) cause symptoms, such as latergia, mucus production, hemorrhagic, and enlarged liver, bleeding abdominal cavity, and acidity in infected fish. Hence, the bacterium harms fish farms. Fish pasteurellosis is an infection caused by the bacterium, which is considered one of the most dangerous diseases in aquaculture due to its high mortality and morbidity. While the species of this genus are widely found in the marine ecosystem and the digestive system of marine creatures, some species are found as symbiotic bacteria that provide specialized light production in marine fish [26].
Immediate delivery of the samples to the laboratory is essential for the diagnosis of the disease. P.damselea subsp. damselea starts to decrease in number after 24 hours when cultured in Stuart’s transport medium from the samples collected from fish and loses its viability after 48-72 hours at 4°C. Therefore, sampling should be done on Amies transport medium. In general, two methods are used in the isolation of Photobacterium species. One of them is by seeding of sea water directly into the medium, the other is the cultivation of fish skin into the medium by enrichment. The sea water taken by the direct seeding method is spread on various media, such as marinated agar, TCBS, TSA, luminous agar, and artificial seawater agar in an amount of 10–100 ml. The agar plates are incubated at 15-20°C. In cases where a small number of bacteria are expected, 5-300 ml of sea water is passed through 0.22-0.45 µm filters to collect bacteria. From these filters, seeding is carried out on media, such as TCBS, TSA, blood agar, luminous agar, marine agar, Photobacterium agar, and artificial sea water agar. Media generally contains 0.5-5% salt. Furthermore, the temperature is an important factor in multiplication, and 5-25°C is generally the preferred incubation temperature. Optimal pH varies between 6-9. In addition to using many sugars other than lactose as a source of carbon, some species also use amino acids as a source of carbon. P. damselea subsp. damselea is sensitive to 500 µg that the vibrio static agent O/129 used in the distinguishing of species in the genus Vibrio [27, 28, 29].
Although various antibiotics can be used in the treatment of the disease, antibiotics should be selected based on the antibiogram results because of the rapid development of antibiotic resistance and the ability to transfer it between bacteria. Florfenicol, enrofloxacin, and oxytetracycline can be used for the treatment of the disease. For protection from infection, vaccination is generally applied by dipping the fish (in the larval stage) twice in one month, and oral inoculation when the weight of the fish reaches 50 mg or
2 g. Inactive bacterial vaccines can usually be applied by immersion in water. In addition, recombinant, subunit, and live vaccines are applied in various forms, such as injection, oral administration, and dipping for research purposes [30, 31, 32].
Listonella Species
The Listonella genus was differentiated from the Vibrio species known to affect both culture and natural fisheries since 1817 by using 5S rRNA gene sequences for phylogenetic analysis of bacteria in 1985. It has been reported that the Listonella genus includes L. anguillarum, L. damselae, and L. pelagia [33]. The species L. damselae was subsequently reclassified as Photobacterium damselae as a result of 16S rRNA gene sequence analysis [34].
L. pelagia (V. pelagia) was first isolated from sea water in 1971. It has been defined as the most frequently isolated Vibrio species from sea waters in Britain [35]. This species, which has been defined as non-pathogenic for many years, has been isolated from epidemics in turbot fish and mussels in various countries, especially after 2000 [36].
L. anguillarum, which is defined as the most common bacteria, was isolated from ell in the Baltic sea and named Vibrio anguillarum [37]. The Listonella genus is examined as an opportunistic pathogen of various fish and shellfish in aquatic environments nowadays. L. anguillarum has 23 different O serotypes (O1-O23), and O1 and O2 are known as the most virulent serotypes [38]. O1, O2, and O3 serotypes are frequently detected serotypes in fish mortalities worldwide [39]. O1 and O2, and O1 serotype are the main ones responsible for the infection of the perch and trout, respectively [38]. The most common serotype isolated from aquatic environments is the O1 in Turkey [40, 41].
Listonella spp. is gram-negative, straight or comma- shaped rod bacteria with peritrichous or monotrichous flagella, oxidase-positive, chemoorganotrophic, and able to grow at 25-35°C. They grow best around neutral pH values (6.5 - 7.0), but they can mostly tolerate a pH of 6 to 9. These species with halophilic properties can easily grow in environments containing 1-2% NaCl. There is also a direct relationship between the salt content in the medium and the motility of L. anguillarum. The growing is suppressed at temperatures below 5°C and above 41°C in environments where includes 7% and over NaCl [10].
Listonella Anguillarum (Vibriosis)
The causative agent is Listonella anguillarum (Vibrio anguillarum) which is gram-negative, comma-shaped, motile, facultative anaerobe, club-shaped bacterium non-spore.
Listonella species are found in large amounts in marine environments with a high organic load, soft ground, and no waves. It can grow easily at 25-30°C in media containing 1.5% sodium chloride (NaCl) such as brain-heart infusion, tryptic soy broth, or agar. It produces round, cream-colored colonies on the agar surface. L. anguillarum is halophilic and can survive in different salt concentrations. According to L. anguillarum sequence analysis, it consists of two chromosomes, eight genomic regions on chromosome I, and two genomic regions on chromosome II. Catalase, arginine dihydrolase, ß-galactosidase, and indole produce but not H2S, lysine, and ornithine decarboxylase, phenylalanine deaminase, and urease. L. anguillarum is positive for Voges Proskauer test but not for the methyl red test. Chitin, gelatin, DNA, lipid, and starch are degraded but not esculin. It reduces nitrate and grows in media with 0.3-3% NaCl. It utulises citrate, tartrate, and malonate. Acid is produced from arabinose, cellobiose, galactose, glycerol, maltose, mannitol, sorbitol, sucrose, and trehalose, but not adonitol, dulcitol, erythritol, inocytol, lactose, melibiose, raffinose, rhamnose, salicin, and xylose [11].
Listonella anguillarum (V. anguillarum) is an infectious bacterium that causes significant economic losses in both aquaculture and larviculture worldwide. It is associated with fatal hemorrhagic septicemia in many marine and freshwater fish, especially eels, sole fish, trout, mackerel, cod, salmon and similar fish species, and bivalves and arthropods. The infection is enzootic in eels, which also act as the reservoir of the bacteria [1]. The source of infection is fish excrement, dead fish, and contaminated feed, and it can spread through feces and open wounds. It is transmitted through digestion and skin. Damaged areas on the skin and portals caused by ectoparasites are important sources of contamination. Outbreaks are common during summer when the water temperature is high and the fish’s skin gets damaged due to heavy metals and parasitic infestations. It is associated with opportunistic infections when it is taken place in the intestinal flora [11, 42].
Although the incubation period varies based on virulence, it is 3 days on average. The infection causes mortality between 40-60%. The infection in peracute cases could lead to death without any symptoms, especially in eels. Typically, infected fish demonstrate clinical features such as lack of appetite, lethargy, swimming close to the water surface, and balance disorders. The petechiae in the mouth, gills, abdomen, and anus, ulceration on the skin surface, hemorrhages, and erosions on the fin tips in infected animals occur and necrotize over time (Figure 1). Exophthalmus and corneal opacification may be observed. There are petechial hemorrhages in the subcutaneous, internal organs and muscles, inflammation of the intestine, splenomegaly and necrosis in the spleen, necrosis foci in the kidney and liver, and distinctly anemia. The gills are pale while the liver appears to be hyperemic [11, 42].

Figure 1: External view of fish with L. anguillarum infection. Large hemorrhagic foci the pelvic, pectoral, and anal fins of the rainbow trout (A), Haemorrhaging on the fins and around the operculum of rainbow trout caused by L. anguillarum (B) (permission for the pictures were obtained from Prof. Dr. Ertan Emek Onuk and Assoc. Prof. Dr. Secil Metin).
Nutrient agar containing 1.5–3.5% NaCl, trypticase soy agar (TSA) containing 1–2% NaCl, thiosulfate citrate bile salt sucrose agar (TCBS), salt-starch agar, and bromthymol blue teepole-salt agar can be used for the isolation of the bacterium. L. anguillarum produces bright yellow colonies with yellow haloes on Vibrio anguillarum medium (VAM), which is the selective media (Figure 2). Identification of the bacterium can be made easily using the API 20E system and MALDI-TOF MS. Besides culture-dependent methods, serological methods and molecular techniques such as polymerase chain reaction commonly practice in vibriosis diagnosis. Specific a fluorescently labeled monoclonal antibody test is used in the diagnosis of serotypes 1, 2, and 3 and 4, 5, and 7 which are the environmental serotypes. In addition, multiplex assays based on DNA array technology have high specificity, sensitivity, and throughput [43, 44, 45, 46].
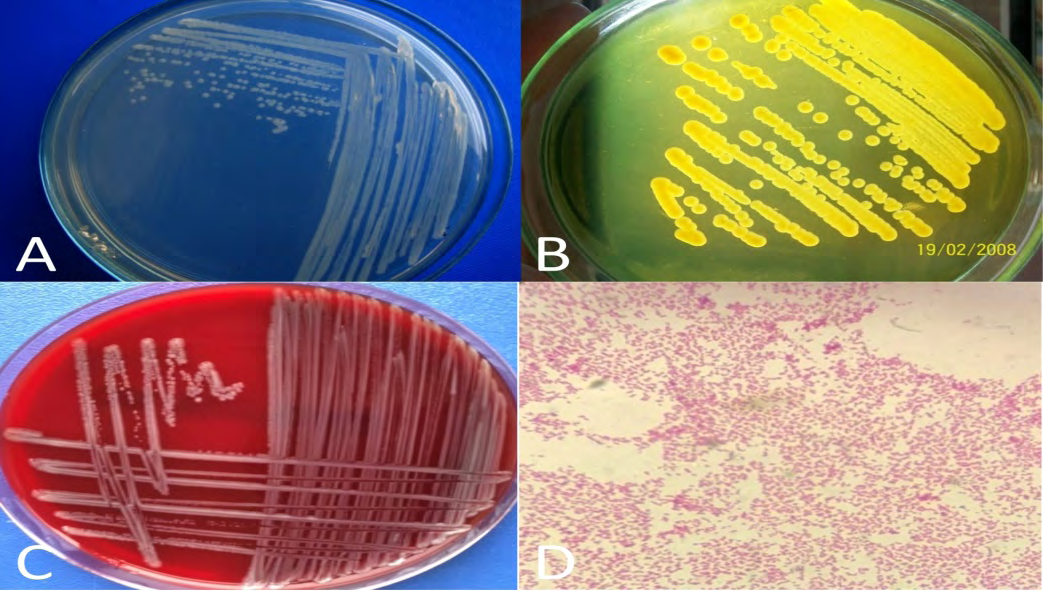
Figure 2: Cream-colored 24-hours pure subculture colonies of L. anguillarum re-isolate on TSA containing 1% NaCl (A), L. anguillarum isolate on TCBS agar (B), L. anguillarum isolate on blood agar (C), a microscopic image of L. anguillarum to gram staining (D) (permission for the pictures were obtained from Prof. Dr. Ertan Emek Onuk and Assoc. Prof. Dr. Secil Metin).
Protecting or treating fish living in nature from infection is practically impossible. However, antibiotics are added to the feed according to the antibiogram test results of the cultured fish. Oxytetracycline, sulfamethaxole-metoprine, erythromycin, and oxanilic acid can be used in the treatment. Vitamin C, selenium, and probiotics can be specifically added to feeds to stimulate the non-specific immune system. Studies have shown that probiotics contribute significantly to decreasing fish mortality [11, 47, 48].
It is very difficult to eliminate L. anguillarum, one of the most persistent infectious agents known, after it settles in the tanks. Sterilization and disinfection procedures should be applied after the tanks have been emptied. L. anguillarum is inactivated for 3 minutes at 44°C and 2 minutes at 47.5°C. Hence, the pool temperature should be increased to protect fishes from infection [11].
Concentrated vaccine administration orally or via the parenteral route for 4 weeks provides protective immunity to the aquaculture. Vaccination should be done by the bathing method for fish weighing between 1-4 g and by injection method for fish weighing greater than 15 g. Sodium salicylate, sodium carpath, and vitamin E supplements have been shown to increase absorption in oral vaccines [30, 31, 32].
Conclusion
Problems with vibriosis are continuously being faced in the aquaculture industries due to various factors such as the vibrio’s variability and adaptive potential, as well as its widespread presence in the marine ecosystem. Despite many studies focusing on the global impact of vibriosis and the causative agent, the appearance of new Vibrio organisms or species creates a considerable threat to human and animal health. Good aquaculture practices, selective use of control agents, and adequate vaccine systems all contribute to improving fish welfare, preventing epidemics, and reducing the destructive economic effects on aquaculture. With the emerging antimicrobial resistance, it should be focused on the search for more environmentally friendly approaches and it should be kept in mind that it is possible to control and prevent diseases with biosecurity applications.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
-
Salwany MYI, Saari NA, Mohamad A, Mursidi FA, Aris AM, et al. (2019) Vibriosis in fish: a review on disease development and prevention. J Aquat Anim Health 31(1): 3-22.
-
Chong R, Bousfield B, Brown R (2011) Fish disease management. Veterinary Bulletin–Agriculture, Fisheries, and Conservation Depart-ment Newsletter 1(8): 1-12.
-
Brenner DJ, Krieg NR, Staley JT (2007) The proteobacteria, Part B: The gammaproteobacteria. Garrity G (Ed.), Bergey’s Manual® of Systematic Bacteriology. Springer Science & Business Media, New York, USA, pp: 491-555.
-
Satitsri S, Pongkorpsakol P, Srimanote P, Chatsudthipong V, Muanprasat C (2016) Pathophysiological mechanisms of diarrhea caused by the Vibrio cholerae O1 El Tor variant: an in vivo study in mice. Virulence 7(7): 789- 805.
-
Begum YA, Rydberg HA, Thorell K, Kwak YK, Sun L, et al. (2018) In situ analyses directly in diarrheal stool reveal large variations in bacterial load and active toxin expression of enterotoxigenic Escherichia coli and Vibrio cholerae. Msphere 3(1): e00517-17.
-
Mohamad N, Mustafa M, Amal MN, Saad MZ, Yasin IS, et al. (2019) Environmental factors associated with the presence of Vibrionaceae in tropical cage‐cultured marine fishes. J Aquat Anim Health 31(2): 154-167.
-
Balasubramanian D, Murcia S, Ogbunugafor CB, Gavilan R, Moreno SA (2021) Cholera dynamics: lessons from an epidemic. J Med Microbiol 70(2): 1-11.
-
Jusserand MB, Copin S, Bris CL, Brauge T, Gay M, et al. (2019) Vibrio species involved in seafood-borne outbreaks (Vibrio cholerae, V. parahaemolyticus and V. vulnificus): Review of microbiological versus recent molecular detection methods in seafood products. Crit Rev Food Sci Nutr 59(4): 597-610.
-
Davenport RJ, Satchell M, Taylor LMWS (2019) Cholera as a ‘sanitary test’of British cities, 1831–1866. Hist Fam 24(2): 404-438.
-
Procop GW, Church DL, Hall GS, Janda WM, Koneman EW, et al. (2017) Koneman’s Color Atlas and Textbook of Diagnostic Microbiology. Philedelphia: Wolters Kluwer Health.
-
Austin B, Austin DA (2016) Bacterial Fish Pathogens: Disease of Farmed and Wild Fish. 5th (Edn.), Switzerland: Springer International Publishing.
-
Zhang Z, Yu YX, Wang K, Wang YG, Jiang Y, et al. (2019) First report of skin ulceration caused by Photobacterium damselae subsp. damselae in net-cage cultured black rockfish (Sebastes schlegeli). Aquac 503: 1-7.
-
Xie J, Mei H, Jin S, Bu L, Wang X, et al. (2021) First report of Photobacterium damselae subsp. damselae infection in the mud crab Scylla paramamosain cultured in China. Aquac 530: 1-8.
-
Perez SF, Hauschild P, Hilgarth M, Vogel RF (2019) Biodiversity of Photobacterium spp. isolated from meats. Front Microbiol 10: 2399.
-
Yildiz FH, Visick KL (2009) Vibrio biofilms: so much the same yet so different. Trends Microbiol 17(3): 109-118.
-
Zhang SD, Isbrandt T, Lindqvist LL, Larsen TO, Gram L (2021) Holomycin, an antibiotic secondary metabolite, is required for biofilm formation of the native producer Photobacterium galatheae S2753. Appl Environ Microbiol 87(11): e00169-21.
-
Vences A, Abushattal S, Matanza XM, Dubert J, Uzun E, et al. (2020) Highly transferable pAQU-related plasmids encoding multidrug resistance are widespread in the human and fish pathogen Photobacterium damselae subsp. damselae in aquaculture areas in the Black Sea. Microb Ecol 80(3): 507-518.
-
Valderrama K, Balado M, Varela DR, Rodríguez J, Vila- Sanjurjo A, et al. (2019) Outer membrane protein FrpA, the siderophore piscibactin receptor of Photobacterium damselae subsp. piscicida, as a subunit vaccine against photobacteriosis in sole (Solea senegalensis). Fish Shellfish Immunol 94: 723-729.
-
Abushattal S, Vences A, Osorio CR (2020) A virulence gene typing scheme for Photobacterium damselae subsp. piscicida, the causative agent of fish photobacteriosis, reveals a high prevalence of plasmid-encoded virulence factors and of type III secretion system genes. Aquac 521: 735057.
-
Terceti MS, Vences A, Matanza XM, Barca AV, Noia M, et al. (2019) The RstAB system impacts virulence, motility, cell morphology, penicillin tolerance and production of type II secretion system-dependent factors in the fish and human pathogen Photobacterium damselae subsp. damselae. Front Microbiol 10: 897.
-
Matanza XM, Osorio CR (2020) Exposure of the opportunistic marine pathogen Photobacterium damselae subsp. damselae to human body temperature is a stressful condition that shapes the transcriptome, viability, cell morphology, and virulence. Front Microbiol 11: 1771.
-
Elbahnaswy S, Elshopakey GE (2020) Differential gene expression and immune response of Nile tilapia (Oreochromis niloticus) challenged intraperitoneally with Photobacterium damselae and Aeromonas hydrophila demonstrating immunosuppression. Aquac 526: 735364.
-
Moi IM, Roslan NN, Leow ATC, Ali MSM, Rahman RNZRA, et al. (2017) The biology and the importance of Photobacterium species. Appl Microbiol Biotechnol 101(11): 4371-4385.
-
Sahu KK, Sherif AA, Davaro R (2020) A rare cause of cellulitis: Photobacterium damselae. J Microsc Ultrastruct 8(1): 25-26.
-
Gupta N, Partridge GJ, Buller NB, Pilmer L, Currie A, et al. (2021) Genetic diversity of Australian isolates of Photobacterium damselae subsp. damselae is associated with virulence to yellowtail kingfish (Seriola lalandi). Aquac 538: 736552.
-
Singaravel V, Gopalakrishnan A, Dewangan NK, Kannan D, Shettu N, et al. (2020) Photobacterium damselae subsp. damselae associated with bacterial myonecrosis and hepatopancreatic necrosis in broodstock Pacific white leg shrimp, Litopenaeus vannamei (Boone, 1931). Aquacult Int 28: 1593-1608.
-
Son MAE, Elbahnaswy S, Ibrahim I (2020) Molecular and histopathological characterization of Photobacterium damselae in naturally and experimentally infected Nile tilapia (Oreochromis niloticus). J Fish Dis 43(12): 1505- 1517.
-
Petchimuthu M (2020) Detection of virulence hemolysin gene (HlyA) in Photobacterium damselae subspecies damselae isolates. J Entomol Zool Stud 8(6): 1326-1330.
-
Yardimci R, Turgay E, Steinum SK (2020) Diagnosis of Photobacterium sanguinicancri in Smout-hound Shark (Mustelus mustelus, Linnaeus 1758). Act Aqu Tr 16(3): 338-343.
-
Hickey ME, Lee JL (2018) A comprehensive review of Vibrio (Listonella) anguillarum: ecology, pathology and prevention. Rev Aquac 10(3): 585-610.
-
Sun X, Jin P, Liu Q, Wang Q, Zhang Y, et al. (2020) A CpG- riched plasmid as vaccine adjuvant reduce antigen dose of an inactivated Vibrio anguillarum vaccine in turbot (Scophthalmus maximus L.). Fish Shellfish immunol 98: 312-317.
-
Xing J, Jiang X, Xu H, Sheng X, Tang X, et al. (2021) Local immune responses to VAA DNA vaccine against Listonella anguillarum in flounder (Paralichthys olivaceus). Mol Immunol 134: 141-149.
-
MacDonell MT, Colwell RR (1985) Phylogeny of the Vibrionaceae, and recommendation for two new genera, Listonella and Shewanella. Syst Appl Microbiol 6(2): 171- 182.
-
Smith SK, Sutton DC, Fuerst JA, Reichelt JL (1991) Evaluation of the genus Listonella and reassignment of Listonella damsela (Love et al.) MacDonell and Colwell to the genus Photobacterium as Photobacterium damsela comb. nov. with an emended description. Int J Syst Bacteriol 41(4): 529-534.
-
Baumann P, Baumann L, Mandel M (1971) Taxonomy of marine bacteria: the genus Beneckea. J Bacteriol 107(1): 268-294.
-
Thompson FL, Thompson CC, Dias GM, Naka H, Dubay C, et al. (2011) The genus Listonella MacDonell and Colwell 1986 is a later heterotypic synonym of the genus Vibrio Pacini 1854 (Approved Lists 1980)–a taxonomic opinion. Int J Syst Evol Microbiol 61(Pt 12): 3023-3027.
-
Bergman AM (1909) Die rote beulenkrankheit des aals. Bericht aus der Koniglichen Bayerischen Versuchsstation 2: 10-54.
-
Mohamad N, Amal MNA, Yasin ISM, Saad MZ, Nasruddin NS, et al. (2019) Vibriosis in cultured marine fishes: a review. Aquac 512: 734289.
-
Noga EJ (2010) Fish disease: diagnosis and treatment. Second edition. Missouri: John Wiley & Sons.
-
Tanrıkul TT, Gultepe N (2011) Mix infections in rainbow trout (Oncorhynchus mykiss Walbaum): Lactococcus garvieae and Vibrio anguillarum O1. J Anim Vet Adv 10: 1019-1023.
-
Parin U, Erbas G, Savasan S, Yuksel HT, Gurpinar S, et al. (2019) Antimicrobial resistance of Vibrio (Listonella) anguillarum isolated from rainbow trouts (Oncorhynchus mykiss). Indian J Anim Res 53(11): 1522-1525.
-
Frans I, Michiels CW, Bossier P, Willems KA, Lievens B, et al. (2011) Vibrio anguillarum as a fish pathogen: virulence factors, diagnosis and prevention. J Fish Dis 34(9): 643-661.
-
Balta F, Dengiz ZB (2017) Serotyping, genetic characterization and antimicrobial susceptibility determination of Vibrio anguillarum strains isolated from farmed rainbow trout (Oncorhynchus mykiss) in the eastern Black Sea. Ankara Üniv Vet Fak Derg 64(4): 321- 328.
-
Onuk EE, Altun S, Duman M, Saticioglu İB, Mustak HK (2018) Phenotypic and genotypic characterization of Listonella anguillarum isolates from rainbow trout. Etlik Vet Mikrobiyol Derg 29: 143-150.
-
Akayli T, Aydin B, Urku C, Kayalar O (2018) Diagnosis of Vibrio anguillarum in cultured rainbow trout (Oncorhynchus mykiss) by different methods. Eur J Biol 77(1): 26-31.
-
Kazazić SP, Popović NT, Perović IS, Babić S, Florio D, et al. (2019) Matrix-assisted laser desorption/ionization time of flight mass spectrometry identification of Vibrio (Listonella) anguillarum isolated from sea bass and sea bream. Plos one 14(11): e0225343.
-
Touraki M, Karamanlidou G, Koziotis M, Christidis I (2013) Antibacterial effect of Lactococcus lactis subsp. lactis on Artemia franciscana nauplii and Dicentrarchus labrax larvae against the fish pathogen Vibrio anguillarum. Aquacult Int 21: 481-495.
-
Huang JB, Wu YC, Chi SC (2014) Dietary supplementation of Pediococcus pentosaceus enhances innate immunity, physiological health and resistance to Vibrio anguillarum in orange-spotted grouper (Epinephelus coioides). Fish Shellfish Immunol 39(2): 196-205.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind