Evident Sexual Dimorphism in the Shape of Subadults of Astyanax altiparanae (Teleostei, Characidae)
The fish species known as “lambari” inhabit small streams, lakes, and large rivers throughout the entire tropical environment of the New World; however, there is limited quantitative information regarding its morphological variations. Our investigation was aimed at studying the sexual dimorphism in subadults of Astyanax altiparanae, using geometric morphometrics to compare sexes in terms of not only size (height and weight) but also shape. A total of 47 specimens (34 males and 13 females) was randomly collected. Photographs of the lateral plane were obtained without sacrificing the specimens, and eight landmarks were placed in the images. The results showed statistically significant differences between sexes. The length of the snout and thickness of the abdomen and caudal peduncle were greater in females, whereas males presented greater dorsal curvature. Females had tall and robust bodies, whereas males were longer and thinner. The difference in shape is probably associated with the species' defensive or survival strategies. However, whether there are ecological advantages or behavioral aspects in sexual dimorphism is still poorly understood, and further research is required to provide data that can explain the causes.
Pere M Parés-Casanova*, Daniel Leonardo Cala1 and Arcesio Salamanca Carreño2
Introduction
Morphological variations make it possible to distinguish the structures that represent divergences, provided that they are measurable and/or quantifiable. Numerous investigations have focused on identifying traits associated with habits, physiology, and even reproductive behavior, at the individual, population, or species levels [1]. The classification of morphometric variability is based on the function of adaptive mechanisms, assuming that the shape of organisms is not only determined by their genetics, but also by their ecology and environmental pressures, which causes them to differ markedly in their morphology within and between populations [2]. Complex patterns of morphometric variation in organisms can be discerned through statistical analyses, simultaneously considering the variation of quantitative traits [3].
Geometric morphometrics (GM) is used to compare sizes and quantify profiles, geometries, conformations, and shapes, regardless of the variation that size may represent, and to study correlations between variation and environmental and morphological parameters [4, 5, 6]. The technique makes it possible to evaluate differences between morphometric conformations through statistical procedures and facilitates the graphical representation of results [7, 8].
There are many species of fish, commonly known as “lambari”, genus Astyanax, which inhabit small streams, lakes, and large rivers in the watersheds throughout the entire tropical zone of the New World [9, 10]. Astyanax belongs to the family Characidae, the largest group of the Tetragonopterinae and constitutes one of the dominant genera throughout South America. There are approximately 100 nominal species and subspecies described in this genus, even though many aspects of its taxonomy are still unknown [9]. The yellow-tailed lambari, Astyanax altiparanae, is a small species that reaches 10–15 cm in length and can weigh up to 60 g. It is an opportunistic generalist species [11] with rapid growth rate, reaching sexual maturity at 4 months of age under farming conditions, when it has a length of 7–9 cm for males and 12–15 cm for females [12]. Although multiple studies on this genus have been conducted [13], there are limited reports on GM, especially on Astyanax altiparanae.
This investigation is aimed at studying sexual dimorphism on Astyanax altiparanae using GM, making possible the detection of possible sexual dimorphism in a pure way, not only in terms of size (height and weight).
Materials and Methods
This study was conducted at the Jaboticabal Campus of Universidad Estadual Paulista (Brazil). A total of 47 subadult specimens (34 males and 13 females) of “lambari” or “river sardine” (Astyanax altiparanae) were randomly collected from water bodies within the university. The animals were transported to the UNESP Aquaculture Center (CAUNESP) located at 21° 14’ 05” latitude, 48° 17’ 09” W longitude, and at an altitude of 615.01 m. (Table 1) presents the height and weight values of both sexes. After capture, the fish were transferred to a water/ice tub to take photographs of the lateral plane (left or right, equally) (Figure 1); of each fish with a digital camera, without sacrificing them. The lateral plane was oriented as parallel to the camera as possible, and the camera was placed far enough to ensure that the fish took up only part of the visual field to avoid distortions of the relative positions of the angles due to parallax error. A 50-mm pattern was included in each shot. The second author, Cala, was responsible for this part. The images were saved in the .jpg format and then transferred to the computer. An image file was created with the tpsUtil program v. 1.70 Rohlf, et al. [14] to digitize the homologous points, and the tpsDig2 program v. 1.40 Rohlf, et al. [14] was used to place each of the homologous points (landmarks) in the images. A total of eight landmarks were subsequently placed in the images as follows: 1: tip of the upper jaw; 2: middle insertion of the caudal fin; 3: dorsal projection of the anterior margin of the cleithrum; 4: anterior margin of the cleithrum; 5: most dorsal part of the curvature of the back; 6: most ventral part of the belly; 7: dorsal insertion of the caudal fin; and 8: ventral insertion of the caudal fin. The first author, Parés, was responsible for the placement of these landmarks. The landmarks were aligned and rescaled using Generalized Procrustes Analysis, by which all information not related to the shape, such as the variation in the position of the specimens, was removed.
| Height (cm) | Weight (g) | |
|---|---|---|
| Males (n = 34) | 5.45 ± 0.49 | 4.01 ± 1.50 |
| Females (n = 13) | 5.54 ± 0.89 | 5.10 ± 2.50 |
Table 1: Body size and weight values of Astyanax altiparanae (n = 47).

A repeatability analysis was carried out to determine whether the homologous points in the specimens were marked correctly. The analysis consisted of randomly selecting a subsample of 25 individuals whose homologous points had been digitized and digitizing them again. Using the PAST application, both replicates were compared using a Multivariate ANalysis Of VAriance (MANOVA) test, and the repeatability values for the X and Y components of each homologous point were obtained.
The Kolmogorov–Smirnov D test was used to compare distribution by sex, and the shape distribution between sexes was compared using MANOVA analysis. Weight and height comparisons were conducted with the Mann–Whitney U test. The data were processed with the MorphoJ v. 1.06c and PAST v. 2.17c statistical packages. The data were tested at a level α = 0.05 to find significant differences [15].
Results
The two replicate subsamples did not demonstrate statistically significant differences between them (P = 0.899). The precision at which the seven homologous points were placed in the specimens was 99.2%. Table 2 shows the repeatability value (between 0 and 1) for the X and Y components of the eight landmarks, average repeatability for each landmark, and total repeatability [16, 17].
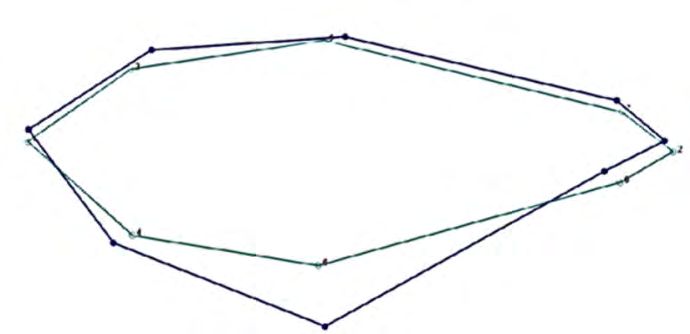
The Kolmogorov–Smirnov test did not demonstrate differences in the distribution between sexes regarding length (D = 0.325, P = 0.220). There were no differences between sexes in terms of size (U = 183.5, P = 0.377; U = 151, P = 0.098 for height and weight, respectively). The MANOVA test revealed statistically significant differences between sexes (P = 0.007). Discriminant analysis showed that the first two axes were significantly different (Wilk’s lambda = 0.345; F16.30 = 3.547; P = 0.0013) (Figure 2). The first canonical function, which accounted for 92.09% of the variance, showed that both sexes were clearly differentiated. Females appeared at the positive end and were basically differentiated by a longer snout and thicker abdomen and caudal peduncle. Males, on the other hand, showed higher dorsal curvature (Tables 2&3). The deformation grid shows these variations with respect to the average configuration (Figure 3). The classification percentage was 89.36% because some females occurred mixed among males. Moreover, the quadratic Mahalanobis distance indicated that there were significant differences between sexes (P < 0.0001).
| Homologous point | X-coordinate | Y-coordinate | Average |
|---|---|---|---|
| 1 | 0.988 | 0.996 | 0.992 |
| 2 | 0.994 | 0.996 | 0.995 |
| 3 | 0.989 | 0.997 | 0.993 |
| 4 | 0.986 | 0.993 | 0.99 |
| 5 | 0.987 | 0.997 | 0.992 |
| 6 | 0.983 | 0.996 | 0.989 |
| 7 | 0.993 | 0.996 | 0.995 |
| 8 | 0.993 | 0.995 | 0.994 |
| Total repeatability | 0.993 |
Table 2: Repeatability value for each of the X and Y components of the eight homologous points placed in the digital images of a
| Coordinate | Eigenvalues |
|---|---|
| x1 | 339.754 |
| y1 | 121.671 |
| x2 | −0.0004 |
| y2 | 369.192 |
| x3 | −121.438 |
| y3 | 574.95 |
| x4 | −50.105 |
| y4 | −91.223 |
| x5 | −30.825 |
| y5 | −210.460 |
| x6 | 60.991 |
| y6 | −796.805 |
| x7 | 283.91 |
| y7 | 53.202 |
| x8 | −482.282 |
| y8 | −20.527 |
Table 3: Coefficient matrix of the discriminant analyses in the study of Astyanax altiparanae (n = 47). The eigenvalues of the mo
Discussion
Fish measurements and weights are used for the characterization of growth, sexual maturation, maximum size, and population structure and additionally, for the specific identification and differentiation between sexes of a species. The morphometric information related to A. altiparanae is not widely documented in the literature.
Our results demonstrate that the sexes in A. altiparanae could be well differentiated by the shape of the body. Females had tall and robust bodies, and males had longer and thinner bodies due to morphological deformations showing variations especially in body height and caudal peduncle. Differences in shape are probably associated with the species’ defensive or survival strategies. For example, females with “deeper” bellies could harbor larger or more numerous eggs and guarantee better defense during the most vulnerable times such as laying. Moreover, more “elongated” males could imply a lower energy investment in growth and an increase in their mobility capacity, a condition that would increase their ability to locate females, ease of courtship, and forced insemination.
It is worth noting that the morphometric analysis was limited to subadult organisms. Discriminant measures are likely to change in juveniles, since fish show body changes during growth related to habitat and diet. In view of the aforementioned factors, it is necessary to understand how the morphometric characteristics change in A. altiparanae, if the differences are maintained. Therefore, these results should be considered with caution. In order to know if the distribution of resources can be a determining factor that facilitates their coexistence, it is also advisable to associate the morphological and osteological characteristics with the species’ feeding habits.
Subadult males and females of A. altiparanae could therefore be differentiated by body shape, i.e., females had tall and robust bodies, and males had longer and thinner bodies. The difference in shape is probably associated with the species’ defensive or survival strategies. However, the existence of ecological advantages or behavioral aspects in sexual dimorphism remains poorly understood, and further research is necessary to establish causal relationships.
Acknowledgement
We/The authors thank Crimson Interactive Pvt. Ltd. (Enago)-https://www.enago.com/es/ for their assistance in manuscript translation and editing.
Authors Contributions
Pere Miquel Parés-Casanova: Writing-original draft preparation, Methodology, Formal analysis, Writing- review and editing, Writing-original draft preparation.
Daniel Leonardo Cala: Photograph’s acquisition, Data collection, Writing-original draft preparation, Arcesio Salamanca-Carreño: Writing- review and editing, Writing- original draft preparation, Conceptualization.
Ethical Statement
The approval of the Animal Ethics Committee was not required, as we used the animals only to take the photograph and then they were released to their natural habitat.
Conflicts of Interest
The authors declare that they have not had any type of conflicts of interest, commercial or otherwise, throughout the preparation and subsequent writing of this article.
References
-
Souza MA, Fagundes DC, Leal CG, Pompeu PS (2014) Ecomorphology of Astyanax Species in streams with different substrates. Zoología (Curitiba) 31(1): 42-50.
-
McCoy M, Bolker BM, Osenberg CW, Miner BG, Vonesh JR (2006) Size correction: comparing morphological traits among populations and environments. Oecología 148: 547-554.
-
Galvão GA (2011) Comparação morfométrica de machos e fêmeas de Astyanax bimaculatus (Linnaeus, 1758) capturados em dois açudes da Bacia do Rio Moxotó (PE) sob a influência do Projeto São Francisco. (Graduação em Medicina Veterinária). Universidade Federal do Vale do São Francisco, Campus de Ciências Agrárias.
-
Bookstein FL, Schäfer K, Prossinger H, Seidler H, Fieder M, et al. (1999) Comparing frontal cranial profiles in archaic and modern Homo by morphometric analysis. Anat Rec 257(6): 217-224.
-
Toro IM, Manriquez SG, Suazo GI (2010) Geometric Morphometry and the Study of Biological Forms: From Descriptive Morphology to Quantitative Morphology. International Journal of Morphology 28(4): 977-990.
-
Ruiz CR, Román Valencia C, Herrera MB, Peláez O, Ermakova AA (2011) Variación morfológica de las especies de Astyanax, subgénero Zygogaster (Teleostei, Characidae). Animal Biodiversity and Conservation 34(1): 47-66.
-
Bookstein FL (1992) Morphometric Tools for Landmark Data: Geometry and Biology. Cambridge University Press.
-
Rohlf F, Marcus LF (1993) A revolution in morphometrics. Trends in Ecology & Evolution 8(4): 129-132.
-
Prioli SM, Prioli AJ, Júlio HF, Pavanelli CS, Oliveira AV, et al. (2002) Identification of Astyanax altiparanae (Teleostei, Characidae) in the Iguaçu River, Brazil , Based on Mitochondrial DNA and RAPD Markers. Genet. Mol. Biol 25(4): 421-430.
-
Catilho Almeida RB (2007) Astyanax altiparanae (Pisces, Characiformes) como modelo biológico de espécie de peixe para exploração zootécnica e biomanipulação. Tese Doutorado, Universidade Estadual Paulista - UNESP, Brazil, pp: 1-119.
-
Almeida FS, Santana CA, Lima DM, Andrade LH, Súarez YR, et al. (2016) Discrimination of Astyanax altiparanae (Characiformes, Characidae) populations by applying Fourier transform-infrared photoacoustic spectroscopy in the fish scales. Infrared Physics & Technology 76: 303- 307.
-
Castro JP, Moura MO, Moreira-Filho O, Shibatta OA, Santos MH, (2014) Evidence of incipient speciation in Astyanax scabripinnis species complex (Teleostei: Characidae). Neotropical Ichthyology 12(2): 429-438.
-
Stevanato DJ, Ostrensky A (2018) Ontogenetic development of tetra Astyanax lacustris (Characiformes: Characidae). Neotrop Ichthyol 16(2): 1-10.
-
Rohlf FJ (2015) The tps series of software. Hystrix the Italic Jouyrnal of Mammalogy 26(1): 9-12.
-
Klingenberg CP (2011) MorphoJ: An integrated software package for geometric morphometrics. Mol Ecol Resour 11(2): 353-357.
-
Hammer Ø, Harper DA, Ryan PD (2001) PAST: Paleontological Statisticts Software Package for education and data analysis. Palaeontological Association 4(1): 1-229.
-
Ruíz RI, Cipriani R (2007) Geometric morphometry of Astyanax siape (Pisces:Characidae). Dahlia (Rev. Asoc. Colomb. Ictiol.) 9: 63-75.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind