Molecular Analysis of Pomphorhynchus kashmirensis based on 18S rDNA and ITS-rDNA
Pomphorhynchus kashmirensis Kaw, 1941 was redescribed to correct inadequacies in the original description. We provide a molecular characterization to qualify its now completed morphological description and to compare with related taxa. Sequences of the 18S rDNA and ITS-rDNA datasets of specimens of P. kashmirensis were generated for assessment of gene diversity and the phylogenetic analyses. Comparative sequence analysis indicated interspecific variation between P. kashmirensis with different species of Pomphorhynchus was 0.2-1.4% and 2.4-36.4% based on 18S rDNA and ITS-rDNA datasets, respectively. The ITS-rDNA region is more variable than 18S rDNA within members of the family Pomphorhynchidae and it appears appropriate for assessment of their biodiversity. Our phylogenetic analyses showed that taxonomic position of the species P. kashmirensis is closely related with P. tereticollis and P. laevis. Also, the systematic status of Tenuiproboscis and Longicollum is uncertain within the family Pomphorhynchidae. Therefore, further molecular investigations will be needed for better understanding of the phylogenetic relationships in this family.
Introduction
Pomphorhynchus kashmirensis Kaw, 1941 is one of 9 species of Pomphorhynchus Monticelli, 1905 known from the Jammu-Kashmir regions of the Northern Indian Subcontinent. The original description from Triplophysa kashmirensis Hora, 1922 (Nemacheilidae) was inadequate as much of its morphological features could not be adequately visualized or confirmed in text or in illustrations [1]. Only one taxonomic treatment by Fotedar and Dhar was reported since its original description and before the redescription from specimens obtained from Schizothorax plagiostomus Heckel, 1838 by Amin et al. In the redescription, comparisons with the Kaw and Fotedar and Dhar accounts were made and 12 SEM images were added presenting new features not previously provided [2, 3]. This study complements the morphological aspects of P. kashmirensis as redescribed by providing its molecular profile for the first time based on of the 18S rDNA and ITS-rDNA datasets. Furthermore, its phylogenetic relationships with other species of the genus Pomphorhynchus and other members of Pomphorhynchidae are analyzed and discussed.
Materials and Methods
DNA Extraction and PCR Amplification
The P. kasmirensis material was collected from Schizothorax plagiostomus in the Jammu-Kashmir regions in the Northern Indian subcontinent. For molecular analysis, the genomic DNA was extracted from ethanol-preserved worms (n=3) using Qiagen DNeasy tissue kit (Qiagen Inc., Valencia, California, USA) according to manufacturer’s instructions. Finally, purified DNA was kept at − 20 °C until use.
The PCR mixture was performed in a final volume 30 μL containing 15 μL of 2X PCR premix with 1.5 mM MgCl2 (Ampliqon, Denmark), 20 ρM of each primer and 2 μL of the extracted DNA. The partial 18S region was amplified using the forward primer (5′-AGATTAAGCCATGCATGCGTAAG-3′) [4] and reverse primer (5′- ACCCACCGAATCAAGAAAGAG-3′) [5]. PCR amplification of the ITS1-5.8S-ITS2 ribosomal RNA gene was performed using the primers BD1 (5′ GTCGTAACAAGGTTTCCGTA-3′) and BD2 (5-TATGCTTAAATTCAGCGGGT-3′) [6].
The temperature profile for the 18S rRNA gene included an initial denaturation step at 95°C for 5 min followed by 35 cycles of denaturation at 95 °C for 30 s, annealing at 61°C for 30 s and extension at 72°C for 60 s, followed by a final extension step at 72°C for 7 min. The PCR conditions of ITS-rDNA region amplification consisted of initial denaturation at 95°C for 5 min, 35 cycles of 95°C for 30 s, 55°C for 30 s, and 72°C for 90 s, followed by a final extension at 72°C for 10 min. The PCR products were separated by electrophoresis on a 1.5% agarose gel and visualized with UV transluminator (Vilber Lourmat, Collégien, France). Finally, PCR products were sequenced by an ABI 3730 DNA Analyzer (Applied Biosystems, Foster City, CA, USA).
The sequence results were edited and trimmed by Chromas software v.2.01 (Technelysium Pty Ltd., Brisbane, Queensland, Australia). Edited sequences were compared with GenBank submitted sequences using the Basic Local Alignment Search Tool (BLAST) [7]. The sequences of 18S rRNA and ITS-rDNA were deposited in GenBank (Accession Numbers: MZ381411-MZ381413 for 18S rRNA and MZ381446-MZ381448 for ITS-rDNA).
Phylogenetic Analysis
Phylogenetic trees were constructed using Maximum- likelihood algorithm and tamura-3- parameter model in MEGA6 software [8]. Also, genetic distances were calculated with p-distance model. The bootstrap value with 1000 replications were used for evaluation of reliability of the tree. The sequences used for the phylogenetic analysis are listed in Tables 1 & 2.
| Species | Host | Location | GenBank accession no. | Reference | |
|---|---|---|---|---|---|
| Pomphorhynchus kashmirensis (Kaw, 1941) | Schizothorax plagiostomus | India | MZ381411-MZ381413 | This study | |
| Pomphorhynchus tereticollis (Rudolphi, 1809) | Gammarus pulex | France | AY423347 | [9] | |
| Pomphorhynchus laevis (Zoega in Müller, 1776) | Squalius cephalus | Croatia | KF559309 | [10] | |
| Pomphorhynchus laevis (Zoega in Müller, 1776) | Rutilus rutilus | Germany | JX014223 | [11] | |
| Pomphorhynchus zhoushanensis (Li, Chen, Amin & Yang, 2017) | Oplegnathus fasciatus Temminck & Schlegel | China | KY490049-59 | [12] | |
| Longicollum pagrosomi (Yamaguti, 1935) | Oplegnathus fasciatus Temminck & Schlegel | China | KY490052 | [12] | |
| Longicollum pagrosomi (Yamaguti, 1935) | Red sea bream | South Korea | KX641270 | [13] | |
| Rhadinorhynchus lintoni (Cable & Linderoth, 1963) | Selar crumenophthalmus | USA | JX014224 | [11] | |
| Rhadinorhynchus pristis (Rudolphi, 1802) | Gempylus serpens | Indonesia | JX014226 | [11] | |
| Echinorhynchus borealis (Linstow, 1901) | Perca fluviatilis | Finland | MW172281 | [14] | |
| Echinorhynchus truttae (Schrank, 1788) | Thymallus thymallus | Na | AY830156 | [15] | |
| Echinorhynchus gadi (Zoega in Müller, 1776) | Macrourus berglax | Greenland | JX014222 | [11] | |
| Echinorhynchus gymnocyprii (Liu, Wang & Yang, 1981) | Ptychobarbus kaznakovi | China | MT162051 | Lei et al. (unpublished) | |
| Echinorhynchus gymnocyprii (Liu, Wang & Yang, 1981) | Gymnocypris przewalskii | China | MT162047 | Lei et al. (unpublished) | |
| Acanthocephaloides propinquus (Dujardin, 1845) | Gobius bucchichii | Na | AY830149 | [15] | |
| Acanthocephalus lucii (Müller, 1776) | Perca fluviatilis | Na | AY830152 | [15] | |
| Acanthocephalus clavula (Dujardin, 1845) | Anguilla anguilla | Ireland | MW172278 | [14] | |
| Acanthocephalus rhinensis (Amin, Thielen, Münderl, Taraschewski & Sures, 2008) | Anguilla anguilla | Germany | MW172279 | [14] | |
| Acanthocephalus dirus (Van Cleave, 1931) | Asellus aquaticus | Na | AY830151 | [15] | |
| Acanthocephalus anguillae (Müller, 1780) | Asellus aquaticus | Slovenia | MN394415 | [16] | |
| Serrasentis sagittifer (Linton, 1889) | Lethrinus laticaudis | Australia | MF426939 | [17] | |
| Gorgorhynchoides bullock (Cable & Mafarachisi, 1970) | Eugerres plumiere | Na | AY830154 | [15] | |
| Outgroup | |||||
| Oligacanthorhynchus tortuosa (Leidy, 1850) | Didelphis virginiana | Na | AF064817 | [18] | |
Table 1: Acanthocephalan species represented in the phylogenetic analysis with their host species, locations, GenBank accession n
| Species | Host | Location | GenBank accession no. | Reference | |
|---|---|---|---|---|---|
| Pomphorhynchus kashmirensis (Kaw, 1941) | Schizothorax plagiostomus | India | MZ381446-MZ381448 | This study | |
| Pomphorhynchus tereticollis (Rudolphi, 1809) | Platichthys flesus | Germany | JF706705 | [19] | |
| Pomphorhynchus tereticollis (Rudolphi, 1809) | Salmonids | Germany | MT216144 | [20] | |
| Pomphorhynchus laevis (Zoega in Müller, 1776) | Phoxinus phoxinus | Slovakia | AY135413 | [21] | |
| Pomphorhynchus bosniacus (Kiskároly & Čanković, 1967) | Alburnus alburnus | Bosnia and Herzegovina | MK157039 | [22] | |
| Pomphorhynchus bosniacus (Kiskároly & Čanković, 1967) | Barbus barbus | Bosnia and Herzegovina | MH319901 | [22] | |
| Tenuiproboscis keralensis (Kaur, Shamal, Chandran, Binesh, Gishnu, Asokan & Sanil, 2017) | Scatophagus argus | India | KU726600 and KU726601 | [23] | |
| Pomphorhynchus lucyi (Williams & Rogers, 1984) | Micropterus salmonoides | USA | AY135418 | [21] | |
| Echinorhynchus gymnocyprii (Liu, Wang & Yang, 1981) | Ptychobarbus kaznakovi | China | MT162085 | [24] | |
| Echinorhynchus gadi (Zoega in Müller, 1776) | Gadus morhua morhua | Barents Sea | EF107648 | [25] | |
| Echinorhynchus truttae (Schrank, 1788) | Na | Germany | MT216137 | [20] | |
| Acanthocephalus nanus (Van Cleave, 1925) | Cynops pyrrhogaster | Japan | LC100043 | [26] | |
| Acanthocephalus anguillae (Müller, 1780) | Proteus anguinus | Slovenia | MN394425 | [16] | |
| Rhadinorhynchus dorsoventrospinosus (Amin, Heckmann & Nguyen Van Ha, 2011) | Decapterus kurroides | Vietnam | MH384822 | [27] | |
| Neoechinorhynchus bullocki (Doolin & Reyda, 2018) | Catostomus commersonii | USA | MK017784 | [28] | |
| Floridosentis mugilis (Machado Filho, 1951) | Mugil sp. | Gulf of Mexico | KC004179 | [29] |
Table 2: Acanthocephalan species represented in the phylogenetic analysis with their host species, locations, GenBank accession n
Results
An approximately 1225 bp fragment of 18S rDNA gene and a 805 bp fragment of ITS-rDNA region were successfully amplified from the specimens of P. kashmirensis. Based on both datasets, there was no intraspecific genetic divergence found among the obtained isolates of P. kashmirensis. The 18S rDNA dataset (1144 nt) included 20 sequences of species representing five families (Pomphorhynchidae, Rhadinorhynchidae, Echinorhynchidae and Arhythmacanthidae) within the Echinorhynchida and the novel sequences of P. kashmirensis. The ITS- rDNA dataset (747 nt) included 13 sequences of species of four families (Pomphorhynchidae, Rhadinorhynchidae and Echinorhynchidae) of Echinorhynchida and the new sequences of P. kashmirensis.
The nucleotide divergence was noted between P. kashmirensis with P. laevis (KF559309 and JX014223), P. zhoushanensis (KY490049 and KY490050), P. tereticollis (AY423347), Longicollum pagrosomi (KY490052 and KX641270), Rhadinorhynchus lintoni (JX014224), R. pristis (JX014226), Acanthocephaloides propinquus (AY830149), Serrasentis sagittifer (MF426939), Gorgorhynchoides bullocki (AY830154), Echinorhynchus spp. and different species Acanthocephalus based on partial 18S rDNA sequence were 0.2-0.3%, 1.4%, 0.2%, 1.4%, 0.8%, 0.6%, 7.6%, 11.5%, 11.6%, 2.1-3.9% and 7.2-7.5%, respectively. Inter-generic differences between P. kashmirensis with P. tereticollis (JF706705 and MT216144), P. laevis (AY135413), P. bosniacus (MK157039 and MH319901), P. lucyi (AY135418), Tenuiproboscis keralensis (KU726600 and KU726601), Rhadinorhynchus dorsoventrospinosus (MH384822), Echinorhynchus spp. and Acanthocephalus spp. based on partial ITS-rDNA ITS-rDNA region were 2.5%, 2.4%, 5.5%, 36.4%, 15.5-15.6%, 49.2%, 36.3-38.2% and 49.6-50.1%, respectively.
The phylogenetic reconstruction based on the partial sequence spanning the 18S rDNA indicated that our sequences of P. kashmirensis (MZ381411-MZ381413) grouped with P. laevis (KF559309 and JX014223), P. tereticollis (AY423347), R. lintoni (JX014224) and R. pristis (JX014226). Additionally, P. zhoushanensis (KY490049 and KY490050) and L. pagrosomi (KY490052 and KX641270) appeared as sister group of mentioned clades (Figure 1).
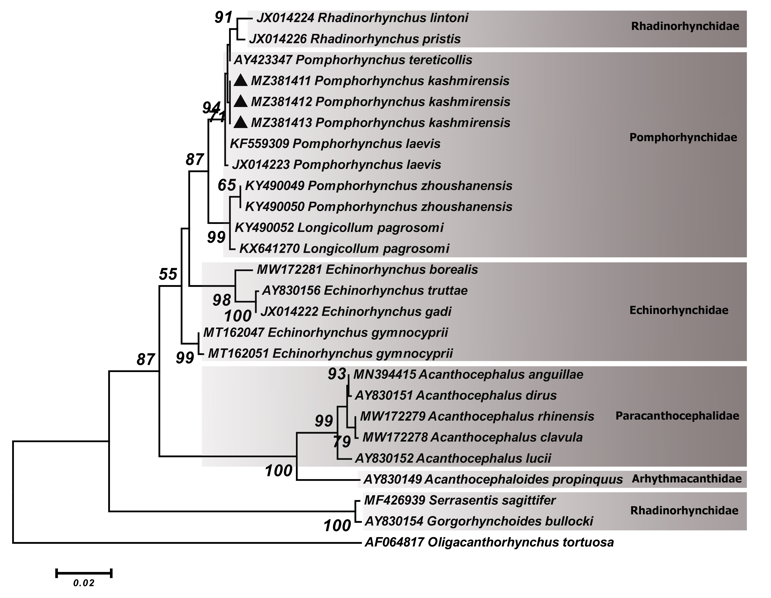
Figure 1: Phylogenetic tree of isolates of the Pomphorhynchus kashmirensis obtained in this study (▲) and other members of the families Pomphorhynchidae, Echinorhynchidae, Arhythmacanthidae and Rhadinorhynchidae retrieved from GenBank based on the partial 18S rDNA gene. The tree was constructed based on the maximum likelihood method and the Tamura 3-parameter model in MEGA6. Oligacanthorhynchus tortuosa sequence was used as the out group. Bootstrap values lower than 50 were omitted.
According to phylogenetic analyses based on the ITS- rDNA ITS-rDNA region, our sequences of P. kashmirensis (MZ381446-MZ381448) clustered with P. tereticollis (JF706705 and MT216144) and P. laevis (AY135413) but with marginal support. Also, P. bosniacus (MK157039 and MH319901) appeared as sister group of mentioned clades with higher bootstrap value. The sequences of T. keralensis (KU726600 and KU726601) as other genus of Pomphorhynchidae family were appeared to be a sister taxon of P. tereticollis, P. laevis and P. bosniacus with 100% of bootstrap support. However, the species of P. lucyi (AY135418) was located at the basal position to the members of the clade of Pomphorhynchidae (Figure 2).
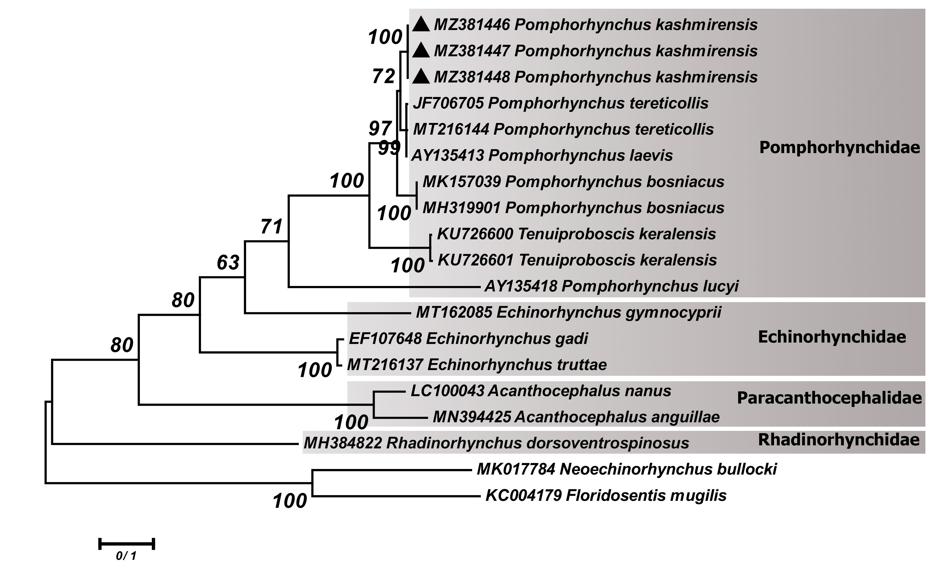
Figure 2: Phylogenetic tree of isolates of the Pomphorhynchus kashmirensis obtained in this study (▲) and other members of the families Pomphorhynchidae, Echinorhynchidae and Rhadinorhynchidae retrieved from GenBank based on the ITS-rDNA region. The tree was constructed based on the maximum likelihood method and the Tamura 3-parameter model in MEGA6. Neoechinorhynchus bullocki and Floridosentis mugilis sequences were used as the out group. Bootstrap values lower than 50 were omitted.
Discussion
The genus Pomphorhynchus presently contains twenty- nine species which are ordinary parasitic worms within the intestinal tract of freshwater fishes, and sometimes marine fishes and amphibians [30]. Until now, only few species of this genus have been characterized using molecular analysis [12, 19, 21, 31, 32, 33]. The validity of P. kashmirensis as redescribed by Amin, et al. is confirmed by the presented molecular analysis. Herein, molecular profile of P. kashmirensis is obtained based on partial 18S rDNA and ITS- rDNA genes for understanding its taxonomic relationships within the family Pomphorhynchidae and with other families of Echinorhynchida.
The level of interspecific nucleotide variation between P. kashmirensis and other species of Pomphorhynchus registered in Gen Bank was 0.2-1.4% and 2.4-36.4% based on 18S rDNA and ITS-rDNA sequences, respectively. These results indicated that ITS-rDNA is more variable between the species than 18S rDNA. Based on the partial 18S rDNA, intergeneric variation between P. kashmirensis and L. pagrosomi another genus of the family Pomphorhynchidae, was 1.4%. Also, based on partial ITS sequences, inter-generic variations between P. kashmirensis and T. keralensis sequences another genus of Pomphorhynchidae family was 15.5-15.6%. Similar to our study, other researchers claimed ITS rDNA sequences has high level of interspecific nucleotide variation in different species of Pomphorhynchus [21, 12]. In addition, some molecular studies on acanthocephalans illustrated that ITS rDNA region is a relatively variable region and has been used as a suitable molecular marker for inferring phylogenetic relationships in some acanthocephalan genera such as Acanthogyrus [34], Corynosoma [35], Neoechinorhynchus [36] and Leptorhynchoides [37].
The small subunit 18S rRNA gene is one of the most frequently used molecular markers to determine genetic diversity and phylogenetic relationships among acanthocephalans. This gene is fairly conserved with a slow evolutionary rate and is usually utilized to infer phylogenetic analysis among the major classes of Acanthocephala [38]. In this study, relatively small genetic variation was found between members of the genus Pomphorhynchus based on18S rRNA gene. Also, there were small genetic differences between species of Pomphorhynchus and Rhadinorhynchus based on this gene. This issue has been investigated and it has been concluded that these two sequences are not really Rhadinorhynchus, but actually Pomphorhynchids [39].
The phylogenetic tree of the 18S rDNA gene (Figure 1) showed that the sequences of P. kashmirensis (MZ381411-MZ381413) obtained in the current study grouped with P. laevis (KF559309 and JX014223) from Croatia and Germany and P. tereticollis (AY423347) from France. However, R. lintoni (JX014224) and R. pristis (JX014226) were nested within this cluster but the genus Rhadinorhynchus was located well apart from the members of the family Pomphorhynchidae in the ITS-rDNA tree. Our 18S rDNA phylogenetic tree is similar to a previously published tree [39, 40], where the genus Rhadinorhynchus was found to be clustered with these species of Pomphorhynchus. In addition, P. zhoushanensis (KY490049 and KY490050) from China and L. pagrosomi (KY490052 and KX641270) from China and Korea were grouped together in one cluster and appeared as a sister group of the mentioned clade with high statistical support. Some authors have also observed a similar situation in L. pagrosomi in Pomphorhynchus clade [12, 40].
Similar to the 18S rDNA tree, the phylogenetic tree of ITS-rDNA (Figure 2) also illustrated that the P. kashmirensis has a close relationship with P. tereticollis (JF706705 and MT216144) from Germany and P. laevis (AY135413) from Slovakia. Additionally, P. bosniacus isolates (MK157039 and MH319901) from Bosnia and Herzegovina clustered as a sister group of the mentioned clade with high statistical support. Similar to our study, Li, et al. showed that the genus Tenuiproboscis was nested within the core of Pomphorhynchus in the phylogenetic trees [12]. Meanwhile, the species of P. lucyi (AY135418) isolated from USA was placed separately from other members of genus Pomphorhynchus. In the current study, the higher level of variation in ITS-rDNA region compared to the 18S rDNA gene improved resolution of the relationships within the family Pomphorhynchidae with high bootstrap support.
Similar to our findings, Li et al. proposed that genus Pomphorhynchus is a polyphyletic taxon based on three different genetic markers (18S rDNA, ITS rDNA and Cox1). It is because members of the Pomphorhynchus were mixed with the genera Longicollum and Tenuiproboscis [12]. The current phylogenetic position of Longicollum and Tenuiproboscis remains questionable and more investigations required to elucidate the phylogenetic relationship between these genera.
Conclusion
The present phylogenetic analyses based on both 18S rDNA and ITS-rDNA sequence data showed the taxonomic position of P. kashmirensis being closely related with P. tereticollis and P. laevis. The ITS-rDNA sequences of members of genus Pomphorhynchus have more variation than 18S rDNA which it can be useful marker for achieving a proper assessment of gene diversity and phylogenetic analyses. In addition, the systematic status of Tenuiproboscis and Longicollum is uncertain within the Pomphorhynchidae. Therefore, obtaining sequence data from other members of the family Pomphorhynchidae and providing additional genetic markers would be useful for better understanding of the phylogenetic relationships in this family.
Acknowledgement
The authors are grateful for the financial support of the Department of Biology at Brigham Young University, Provo, Utah and for the Institutional Grant provided by The Parasitology Center, Scottsdale, Arizona, USA.
Compliance with Ethical Standards
Conflict of Interest: The authors declare no conflict of interest and compliance with all relevant ethical standards.
References
-
Amin OM, Heckmann RA, Zargar UR, Chishti MZ, El Naggar AM (2012) The morphology of the long forgotten _Pomphorhynchus_ _kashmirensis_ (Acanthocephala: Pomphorhynchidae) from freshwater fish in Kashmir using SEM, with notes on histopathology. Sci Parasitol 13(3): 93-99.
-
Kaw B (1941) Studies on the helminth parasites of Kashmir. Part I. Description of some new species of the genus _Pomphorhynchus Monticelli_ (1905). Proc Ind Acad Sci (13): 369-378.
-
Fotedar D, Dhar R (1977) On five new species of the genus _Pomphorhynchus Monticelli_ 1905 from freshwater fishes of Jammu and Kashmir, India. Ind J Zool ( 5): 15- 29.
-
Garey JR, Near TJ, Nonnemacher MR, Nadler SA (1996) Molecular evidence for Acanthocephala as a subtaxon of Rotifera. J Mol Evol 43(3): 287-292.
-
Amin O, Sharifdini M, Heckmann R, Zarean M (2020) New perspectives on _Nephridiacanthus_ _major_ (Acanthocephala: Oligacanthorhynchidae) collected from hedgehogs in Iran. J Helminthol 94: e133-e133.
-
Luton K, Walker D, Blair D (1992) Comparisons of ribosomal internal transcribed spacers from two congeneric species of flukes (Platyhelminthes: Trematoda: Digenea). Mol Biochem Parasitol 56(2): 323- 327.
-
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215(3): 403-410.
-
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30(12): 2725-2729.
-
Perrot Minnot MJ (2004) Larval morphology, genetic divergence, and contrasting levels of host manipulation between forms of _Pomphorhynchus laevis_ (Acanthocephala). Int J Parasitol 34(1): 45-54.
-
Smrzlić IV, Valić D, Kapetanović D, Marijić VF, Gjurčević E, et al. (2015) _Pomphorhynchus laevis_ (Acanthocephala) from the Sava River basin: new insights into strain formation, mtDNA-like sequences and dynamics of infection. Parasitol Int 64 (5): 243-250.
-
Verweyen L, Klimpel S, Palm HW (2011) Molecular phylogeny of the Acanthocephala (class Palaeacanthocephala) with a paraphyletic assemblage of the orders Polymorphida and Echinorhynchida. PLoS One 6(12): e28285.
-
Li L, Chen HX, Amin OM, Yang Y (2017) Morphological variability and molecular characterization of _Pomphorhynchus zhoushanensis_ sp. nov.(Acanthocephala: Pomphorhynchidae), with comments on the systematic status of P_omphorhynchus_ Monticelli, 1905. Parasitol Int 66(5): 693-698.
-
Ha NR, Hong EJ, Ryu SY, Sim CH, Chae JS, et al. (2017) Morphological and molecular finding of _Longicollum_ _pagrosomi_ (Acanthocephala: Pomphorhynchidae) in cultured red sea bream from Korea. Korean J Vet Serv 40(3): 169-175.
-
García Varela M, Andrade Gómez L (2021) First steps to understand the systematics of Echinorhynchidae Cobbold, 1876 (Acanthocephala), inferred through nuclear gene sequences. Parasitol Int 81: 102264.
-
García Varela M, Nadler SA (2005) Phylogenetic relationships of Palaeacanthocephala (Acanthocephala) inferred from SSU and LSU rDNA gene sequences. J Parasitol 91(6): 1401-1409.
-
Amin OM, Heckmann RA, Fišer Ž, Zakšek V, Herlyn H (2019) Description of _Acanthocephalus anguillae_ _balkanicus_ subsp. n.(Acanthocephala: Echinorhynchidae) from Proteus anguinus Laurenti (Amphibia: Proteidae) and the cave ecomorph of _Asellus aquaticus_ (Crustacea: Asellidae) in Slovenia. Folia Parasitol 66: 1-13.
-
Barton DP, Smales L, Morgan JA (2018) A redescription of _Serrasentis_ _sagittifer_ (Rhadinorhynchidae: Serrasentinae) from _Rachycentron_ _canadum_ (Rachycentridae) with comments on its biology and its relationship to other species of _Serrasentis._ J Parasitol 104(2): 117-132.
-
García Varela M, de León GPP, De La Torre P, Cummings MP, Sarma S, et al. (2000) Phylogenetic relationships of Acanthocephala based on analysis of 18S ribosomal RNA gene sequences. J Mol Evol 50(6): 532-540.
-
Špakulová M, Perrot Minnot MJ, Neuhaus B (2011) Resurrection of _Pomphorhynchus tereticollis_ (Rudolphi, 1809)(Acanthocephala: Pomphorhynchidae) based on new morphological and molecular data. Helminthologia 48(4): 268-277.
-
Ros AF, Basen T, Teschner RJ, Brinker A (2020) Morphological and molecular data show no evidence of the proposed replacement of endemic _Pomphorhynchus_ _tereticollis_ by invasive _P. laevis_ in salmonids in southern Germany. PloS one 15(6): e0234116.
-
Král’ová-Hromadová I, Tietz DF, Shinn AP, Špakulová M (2003) ITS rDNA sequences of _Pomphorhynchus laevis_ (Zoega in Müller, 1776) and _P. lucyi_ williams & Rogers, 1984 (Acanthocephala: Palaeacanthocephala). Syst Parasitol 56(2): 141-145.
-
Nedić Z, Smrzlić IV, Paraš S, Nikolić V (2019) _Pomphorhynchus bosniacus_ Kiškarolj & Čanković 1969 (Acanthocephala), intestinal parasite from the Sava River, Bosnia and Herzegovina: new insights on phylogeny, infection dynamics and histopathology. Bull Eur Assoc Fish Pathol 39(3): 93-105.
-
Kaur P, Shamal P, Chandran A, Binesh C, Gishnu M, et al. (2017) Morphometric and molecular characterisation of _Tenuiproboscis keralensis_ n. sp. infecting marine and brackish water fishes from the south-west coast of India with a note on morphological plasticity. Parasitol Res 116(11): 3131-3149.
-
Lei MT, Cai JZ, Li CH, Fu Y, Sun J, et al. (2020) Prevalence and genetic diversity of _Echinorhynchus gymnocyprii_ (Acanthocephala: Echinorhynchidae) in schizothoracine fishes (Cyprinidae: Schizothoracinae) in Qinghai-Tibetan Plateau, China. Parasit Vectors 13(1): 1-11.
-
Sobecka E, Szostakowska B, MacKenzie K, Hemmingsen W, Prajsnar S, et al. (2012) Genetic and morphological variation in _Echinorhynchus gadi_ Zoega in Müller, 1776 (Acanthocephala: Echinorhynchidae) from Atlantic cod Gadus morhua L. J Helminthol 86(1): 16-25.
-
Nakao M (2016) _Pseudoacanthocephalus toshimai_ sp. nov. (Palaeacanthocephala: Echinorhynchidae), a common acanthocephalan of anuran and urodelan amphibians in Hokkaido, Japan, with a finding of its intermediate host. Parasitol Int 65(4): 323-332.
-
Chaudhary A, Amin OM, Heckmann R, Singh HS (2020) The Molecular Profile of _Rhadinorhynchus_ _dorsoventrospinosus_ Amin, Heckmann, and Ha 2011 (Acanthocephala: Rhadinorhynchidae) from Vietnam. J Parasitol 106(3): 418-427.
-
Doolin ML, Reyda FB (2018) A new species of _Neoechinorhynchus_ (Acanthocephala: Neoechinorhynchidae) from white sucker (_Catostomus_ _commersonii_) in New York J Parasitol 104(6): 671-678.
-
Pinacho Pinacho CD, Sereno Uribe AL, Garcia Varela M (2014) Morphological and molecular data reveal a new species of _Neoechinorhynchus_ (Acanthocephala: Neoechinorhynchidae) from _Dormitator maculatus_ in the Gulf of Mexico. Parasitol Int 63(6): 763-771.
-
Amin OM (2013) Classification of the Acanthocephala. Folia Parasitol 60(4): 273-305.
-
García Varela M, Mendoza Garfias B, Choudhury A, de León GPP (2017) Morphological and molecular data for a new species of _Pomphorhynchus Monticelli_, 1905 (Acanthocephala: Pomphorhynchidae) in the Mexican redhorse _Moxostoma austrinum_ Bean (Cypriniformes: Catostomidae) in central Mexico. Syst Parasitol 94(9): 989-1006.
-
Mauer K, Hellmann SL, Groth M, Fröbius AC, Zischler H, et al. (2020) The genome, transcriptome, and proteome of the fish parasite _Pomphorhynchus laevis_ (Acanthocephala). PloS one 15(6): e0232973.
-
Reier S, Sattmann H, Schwaha T, Harl J, Konecny R, et al. (2019) An integrative taxonomic approach to reveal the status of the genus _Pomphorhynchus_ Monticelli, 1905 (Acanthocephala: Pomphorhynchidae) in Austria. Int J Parasitol Parasites Wildl 8: 145-155.
-
Sharifdini M, Amin O, Heckmann R (2021) The molecular profile of _Acanthogyrus_ (_Acanthosentis_) _kashmirensis_ from the Indian subcontinent. Acta Parasitologica pp: 1-8.
-
García Varela M, Aznar FJ, Pérez Ponce de León G, Piñero D, Laclette JP (2005) Molecular phylogeny of _Corynosoma_ Lühe, 1904 (Acanthocephala), based on 5.8 S and internal transcribed spacer sequences. J Parasitol 91(2): 345-352.
-
Martínez Aquino A, Reyna Fabián ME, Rosas Valdez R, Razo Mendivil U, de León GPP, et al. (2009) Detecting a complex of cryptic species within _Neoechinorhynchus_ _golvani_ (Acanthocephala: Neoechinorhynchidae) inferred from ITSs and LSU rDNA gene sequences. J Parasitol 95(5): 1040-1047.
-
Steinauer M, Nickol B, Orti G (2007) Cryptic speciation and patterns of phenotypic variation of a highly variable acanthocephalan parasite. Mol Ecol 16(19): 4097-4109.
-
Garcıa Varela M, de Leon GPP (2015) 9 Advances in the classification of acanthocephalans: evolutionary history and evolution of the parasitism. Parasite Diversity and Diversification: Evolutionary Ecology Meets Phylogenetics pp: 182.
-
Lisitsyna OI, Kudlai O, Smit NJ, Cribb TH (2019) Three new species of acanthocephalans (Palaeacanthocephala) from marine fishes collected off the East Coast of South Africa. Folia Parasitol 66: 012.
-
Chaudhary A, Amin OM, Heckmann R, Singh HS (2020) The Molecular Phylogeny of _Pararhadinorhynchus magnus_ Ha, Amin, Ngo, Heckmann, 2018 (Acanthocephala: Rhadinorhynchidae) from _Scatophagus argus_ (Linn.) (Scatophagidae) in Vietnam. Acta Parasitol 65(3): 610- 619.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework