“Proof of Concept” Rabbit Experiments Demonstrating GHR106 (hIgG4) as GnRH Antagonist
“Proof of Concept” (P-o-C) experiments in rabbits were conducted to demonstrate reversible suppressions of serum reproductive hormones (LH, testosterone or E2) upon a single injection of a humanized monoclonal antibody against human GnRH receptor, GHR106 (hIgG4). In the case of a male rabbit, a single subcutaneous injection of either 1 mg/kg or 3 mg/kg of the antibody was shown to reduce concurrently the serum LH and testosterone concentrations by 60 to 80% for a period of one week or longer. The reproductive hormone levels returned to the normal range after next two weeks. Similarly, the suppressions of LH and estradiol (E2) were also observed within two days upon a single injection of GHR106 (hIgG4) in the female. The E2 levels can decrease to 30-60% of the normal over a period of one week followed by full recovery of both hormones during the next 5-10 days. P-o-C experiments were expanded to a scale of more than 30 rabbits with identical protocols to those of a single rabbit, in addition to the comparisons with those of the negative control. Reversible suppressions of reproductive hormones were observed consistently with all rabbits in the antibody injection groups, and were shown to be statistically significant. The circulation half-life of GHR106 (hIgG4) in rabbits was estimated to be 4 ± 1 day. Therefore, the results of this study revealed that GHR106(hIgG4) can be used as antibody-based long- acting GnRH antagonist with similar biological actions to those of decapeptide GnRH antagonist such as Cetrorelix or Antide. The relatively long half-life of the antibody-based GnRH antagonist, however, may be more beneficial to the treatment of gynecological and fertility-related diseases which may be caused by modulations of human GnRH receptor in humans, and also in other animal species for fertility controls.
Introduction
GRH106 is a monoclonal antibody generated from mouse against N1-29 oligopeptide located in the extracellular domains of human GnRH (Gonadotropin releasing hormone) receptor [1]. Due to a high degree of amino acid sequence homology (>90 -95%), GHR106 cross-reacts with those of monkey, rabbit, dog or cat, but not with those of mouse and rat. GHR106 and its humanized forms were shown to react specifically with human GnRH receptor either in cancer cells or in anterior pituitary [1, 2, 3, 4].
In humans, there is only one type of functional GnRH receptor gene [2, 3]. The main action site of GnRH receptor located in the anterior pituitary is responsible for the release of gonadotropin hormones, LH and FSH upon pulsatile stimulation of GnRH released from hypothalamus.
The roles of GnRH and GHR106 as GnRH antagonist in hypothalamus-pituitary-gonadal axis of mammalian reproductive system need to be briefly explained [1, 2, 3, 4], and is illustrated in Figure 1. GnRH is a decapeptide hormone released from hypothalamus and is essential for male and female reproductive health. Upon release from hypothalamus through blood stream, GnRH interacts with GnRH receptor in the anterior pituitary and stimulates the release of gonadotropins including LH and FSH.
Numerous derivatives, or analogs of GnRH are available and act as agonist or antagonist depending on their respective actions to stimulate or inhibit the release of gonadotropin. Based on previous experimental data, GHR106 was considered as long-acting antibody-based GnRH antagonist [1, 2, 3, 4].
In case of GnRH or its agonists, the binding to GnRH receptor in anterior pituitary will stimulate release of LH and FSH into circulation. Gonadotropins interact with female ovaries to secrete estradiol (E2) and progesterone, whereas testosterone in the male testis can be produced upon interactions with gonadotropins [1, 2, 3, 4]. As demonstrated in Figure 1, reproductive hormones are produced for various reproductive functions in both males and females in humans and several other animal species such as rabbit, dog and cat. In contrast, if GnRH antagonist is present in blood steam to compete with GnRH for the pituitary GnRH receptor, the release of gonadotropins will be inhibited to result in reduction of the gonadal productions of testosterone in the male, or estradiol and progesterone in the female. The objective of our studies is to show that GHR106 acts as GnRH antagonist, similar to that of decapeptide GnRH antagonists such as Cetrorelix or Antide (Figure 1) [3, 4, 5, 6].
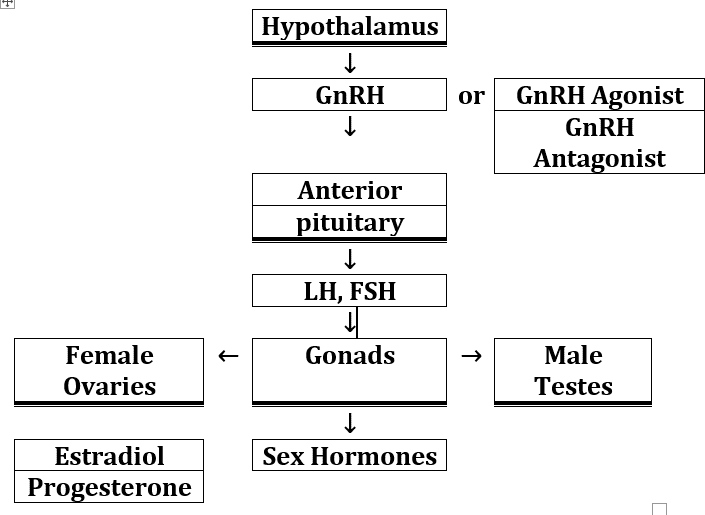
Thick double bottom lines box means organs or tissues.
Single bottom line box means hormones or analogs.
GnRH receptor is also expressed in gonad-related tissues as well as cancer cells [2, 3]. The extra- pituitary GnRH receptor may serve to induce cellular apoptosis upon interactions with GnRH or its analogs through the mechanism of autocrine/ paracrine regulations [2, 3]. GHR106 will also act on GnRH receptor either in anterior pituitary or extra- pituitary tissues including cancer cells, similar to other decapeptide GnRH antagonists.
In this study, the rabbit was selected as animal model to demonstrate that GHR106 monoclonal antibody is GnRH antagonist to act on the anterior pituitary GnRH receptor. Reversible suppressions of reproductive hormones through pituitary-gonadal axis can be induced upon a single injection of this GnRH antagonist with similar action to that of decapeptide GnRH antagonist.
Materials and Methods
Rabbit Experiments Performed by CRO
Multi-rabbit experiments were performed and maintained mainly through CRO, Pocono Rabbit Farm and Laboratory (PRFAL) (Philadelphia, PA). The 30-day experimental protocols were designed by Vancouver Biotech Ltd. New Zealand white rabbits of either sex were employed for all experiments carried out by PRFAL for the injections and scheduled bleedings. Rules and protocols of animal experiments were approved by animal experimentation committee of PRFAL. Mature rabbits (four months or older) were evenly divided into three study groups, each of which were injected subcutaneously on Day 1 with either high dose (3 mg/kg GHR106), low dose (1 mg/kg GHR106) or no injection as the negative control. Following injections, the rabbits were bled every two days at the same time of the day (8-10 AM) until day 29.
The serum specimens of each rabbit were collected and frozen at -20 °C until used later for hormonal assays of LH and Estradiol (female) or testosterone (male).
A Separate Study of Single Rabbit
Experiments were performed in house at Vancouver Biotech Ltd. The study protocols were identical to those of multi-rabbit study performed by PRFAL. Data collected were also included in this report for comparative purposes.
Chemicals
The enzyme immunoassay (EIA) kits for serum testosterone or estradiol concentrations were a generous gift of Perkin Elmer Health Science Center (Hayward, CA}. The LH EIA kits for the rabbit were obtained from Cusabio, USA. The immunoassays of these EIA kits were performed according to instructions specified by the supplier’s package inserts. N
1-29 oligopeptides of GnRH receptors from human, dog and rabbit were custom-synthesized by GenScript (Piscataway, NJ). They were coated separately on microwells for binding assays with GHRl06 (hlgG4) and its N1-29 oligopeptide according to the established procedure [1, 2, 3, 4, 5, 6, 7, 8].
GHR106 Monoclonal Antibody Used in Experiments
GHR106 monoclonal antibody of mouse origin was humanized through LakePharma CRO services [9]. Through the co-development agreement with IO Biosciences Inc. GHR106 (hIgG4) was produced in large quantity for the CRO studies. The quality of the newly produced GHR106 (hIgG4) has been shown to be identical to that obtained from LabePharma Inc. (Belmont, CA) in 2018. Through biological studies, GHR106 (hIgG1) and GHR106 (hIgG4) isoforms have been shown to have comparable binding affinity to human GnRH receptor or its N1-29 oligopeptide. The affinity constants of GHR106 (hIgG4) to human GnRH receptor were shown to be in the range of 1-4 nM [9].
Results
Summarized Results of Proof-of-Concept Studies with GHR106 (hIgG4)
Selection of Rabbit for Proof-of-Concept Experiments
GHR106 (hIgG4) is a monoclonal antibody derived from immunizations of mouse against N1-29 oligopeptide of human GnRH receptor. An animal model is required to demonstrate that GHR106 (hIgG4) interacts with pituitary GnRH receptor which can lead to the reversible suppression of reproductive hormones. Therefore, N1-29 oligopeptides from different animal species including human, monkey, dog, cat, rabbit and mouse are compared and shown in Table 1.
| Human | MANSASPEQNQNHCSAINNSIPLMQGNLPT |
|---|---|
| Rabbit | MENSASPEQNQNHCSAINNSIPLTQGNLNT |
| Monkey | MANSALPEQNQNHCSVINNSIPLMQGNLPT |
| Cat | MA-SAPPEQNQNHCSAINNSIPLMQGNLPT |
| Dog | MA-SASPEQNQNHCSAVNNSNMLMQGNLPT |
| Mouse | MANNASLEQDPNHCSAINNSIPLIQGKLPT |
Table 1: Comparisons of GnRH receptor (GnRHR) amino acid sequences (N1-30) in human, rabbit, monkey, cat, dog and mouse. High deg
It was apparent that N1-29 oligopeptides between human and rabbit have a high degree of sequence homology and should show comparable binding affinity to GHR106 (hIgG4) and comparable biological activities to their respective GnRH receptors (1-4). Based on this assumption, the rabbit may be selected as a suitable animal model for proof- of-concept experiments in this study.
Additional experiments are essential to demonstrate comparable binding affinity of GHRl06 (hlgG4) to N1- 29 oligopeptides derived from human, dog and rabbit, respectively. Therefore, comparative binding ELISA studies were performed to estimate the relative binding affinity between GHR106 (hIgG4) and microwell-coated N1- 29 oligopeptides derived from human, dog and rabbit, respectively. The results of such binding studies are presented and compared in Figure 2. It was clearly demonstrated that the binding affinities between GHR106 (hIgG4) and N1-29 peptides derived from human, dog, and rabbit, respectively are comparable to one another, when compared with that of unrelated RP215 used as the negative control [6, 7].
![Figure 2: It was clearly demonstrated that the binding affinities between GHR106 (hIgG4) and N1-29 peptides derived from human, dog, and rabbit, respectively are comparable to one another, when compared with that of unrelated RP215 used as the negative control [6,7].](/fulltextimages/8625/fig_2.png)
Figure 2: Binding ELISA to demonstrate GHR106 to N1- 29 oligopeptides derived from GnRH receptor of human, rabbit and dog, respectively. The double log plot of the binding OD vs. GHR106(hIgG4) concentration indicates the similarity of respective bindings of GHR106(hIgG4) to GnRH receptors derived from three different animal species such as human ( ● ), dog ( ▲ ) and rabbit ( ■ ).
In previous studies, we also showed that the three isoforms of GHR106 isoforms including murine GHR106, humanized GHRl06 (hlgG1) and humanized GHR106(hIgG4) are essentially identical in their respective binding affinity and specificity to human GnRH receptor as well as its N1-29 oligopeptide with dissociation constants on the order of 1-4
In the binding ELISA study presented in Figure 2. It has been demonstrated that GHR106 (hIgG4) revealed comparable binding to GnRH receptor or its N1-29 oligopeptides of either human or rabbit.
Therefore, the rabbit was selected for proof-of-concept experiments to demonstrate the reversible suppressions of reproductive hormones upon a single treatment with GHR106 (hIgG4) [9].
Hormonal Effects of Single Rabbit Injections with GHR106 (hIgG4)
Single Rabbit Experiments: Attempts were made to demonstrate that GHR106 interacts with pituitary GnRH receptor, similar to that of decapeptide GnRH antagonist. P-o-C experiments were initiated with the study of hormonal effects to a single rabbit (male or female) upon one injection of GHR106 (hIgG4) on day 1 over a period of two to three weeks.
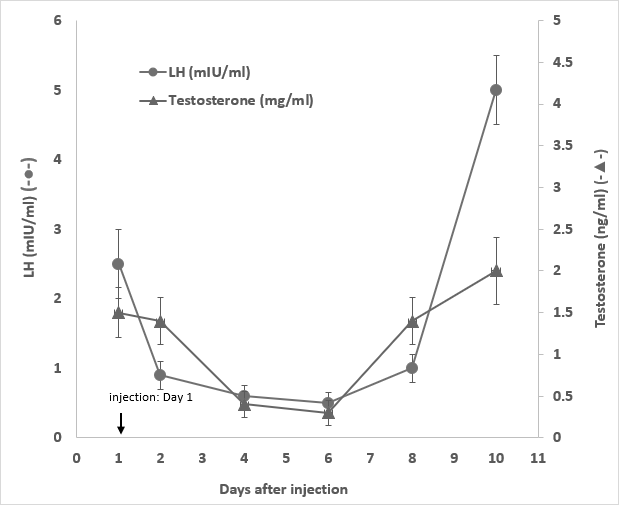
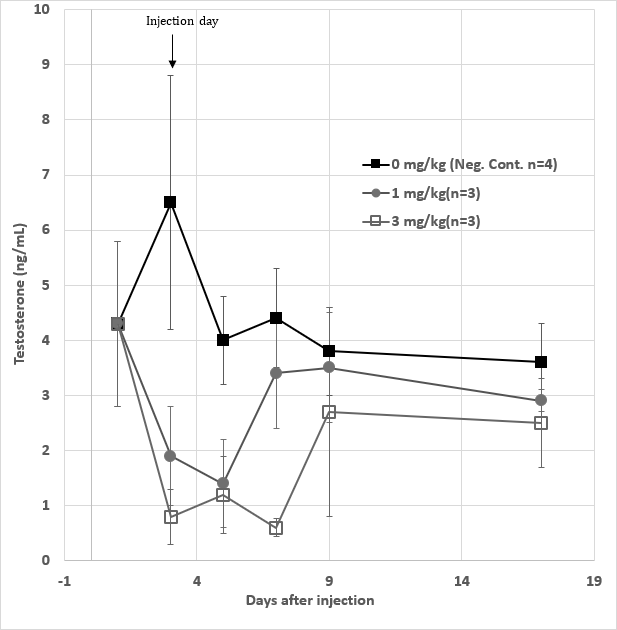
To a given male rabbit, upon a single injection with 1 mg/ kg GHR106 (hIgG4) on day 1, both serum LH and testosterone levels decreased to 60 to 80% of the normal levels from day 4 to day 6. The serum LH and testosterone levels returned to the normal range on day 8. The hormonal profiles are shown in Figure 3.
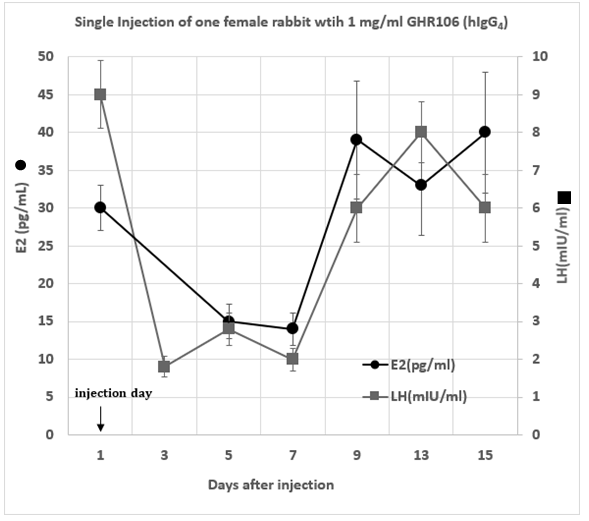
Figure 3: Serum LH (●) and testosterone ( ▲ ) levels of one single male rabbit injected with 1 mg/kg of GHR106(hIgG4) on day 1. They were monitored until day 10. LH and Testosterone serum concentration were plotted as a function of time after the day 1 injection for comparative purpose. (Data presented are means and error bars of three determinations).
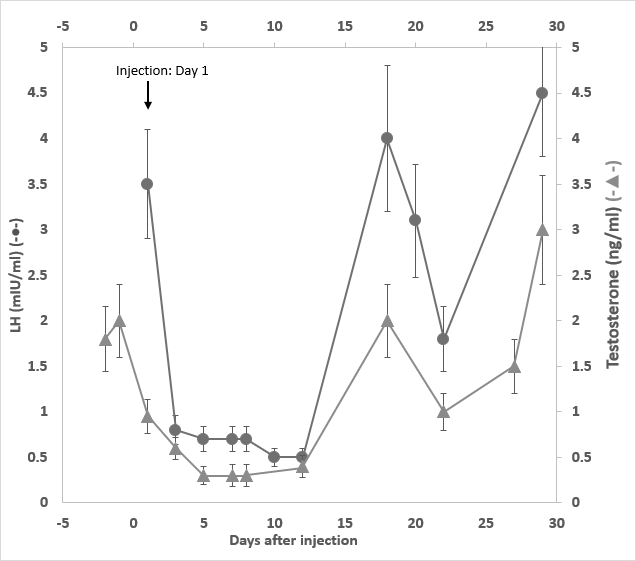
In a separate study, one male rabbit was subject to a single injection with 3 mg/kg of GHR106 (hIgG4) on day 1. Immediate suppressions of serum LH and testosterone were observed on day 3 until day 11. Both reproductive hormones returned to the normal range (2-5 mIU/ml) after day 15 as presented in Figure 4.
The hormonal profiles of serum LH and estradiol (E2) were also monitored with one single subcutaneous injection of one female rabbit either with 1 mg/kg or 3 mg/kg doses of GHR106 (hIgG4). They are demonstrated in Figure 5 and Figure 6, respectively.
A single injection with 1 mg/kg of GHR106 (hIgG4) to one female rabbit on day 1 was performed.
Both serum levels of LH and estradiol decreased significantly by as much as 50 to 70% after day 3. The hormone profiles of both returned to normal ranges after day 9 until day 15.
In a separate study, one female rabbit was injected with 3 mg/kg of GHR106 (hIgG4) on day 1. The hormone profiles including serum LH and E2 were determined until day 20. Both LH and E2 levels decreased immediately by as much as 60 - 80% on day 3 until day 9. The hormonal levels of LH and E2 slowly returned to normal ranges after day 13. The hormone level changes upon a high dose injection (3 mg/kg) were monitored every two days and are presented in Figure 6 for companions.
Average Hormone Profiling from Groups of Rabbits upon Single Injection with GHR106 (hIgG4) • Male Rabbits Experiments Ten male rabbits were evenly divided into three study groups. The negative control groups (n=4) received no injection and serum specimens collected from day 1 and every other two days until day 29.
The low dose group (n=3) received 1 mg/kg injection of GHR106 (hIgG4) on day 1. After pre-injection bleeding and serum specimens were collected according to the same schedules. Similar protocols were applied to the high dose group with 3 mg/kg injection on day 1.
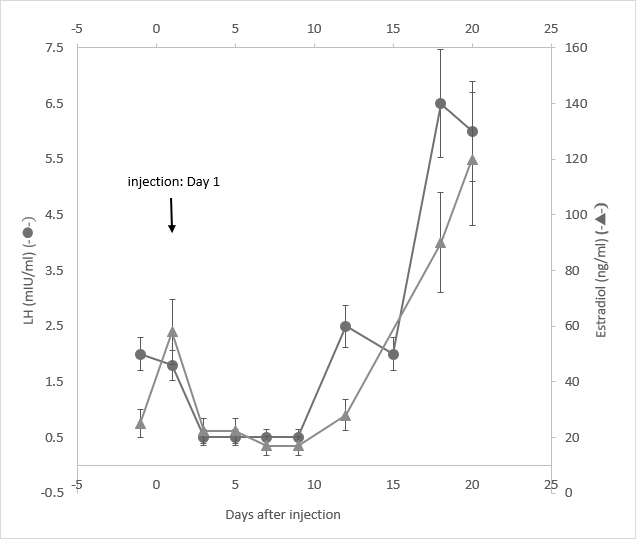
Figure 7: Serum testosterone levels of a total ten male rabbits divided into three experimental groups. The negative control group (n=4) (■) received no injection on day 1, whereas the low dose (●) (n=3) and high dose group (◻) (n=3) received 1 mg/kg and 3 mg/kg injection to rabbits in each group respectively on day 1. The serum testosterone levels were determined and the means plus derivations were determined and presented from day1 to day 17. Data presented are means and standard deviations of the negative control group (n=4) vs. the experimental groups (n=3 for each).
Serum levels of testosterone were determined for rabbits in the same group and mean values plus standard deviations were calculated. They are presented in Figure 7 including those of the negative control, high dose as well as low dose groups. Following day 1 of injection, the low dose groups showed significant decrease by 50%-70% in serum testosterone levels on day 3 and day 5, after which the testosterone levels returned to the normal on day 7. In the case of high dose group, significant decreases (70-80%) of the testosterone levels were observed between day 3 and day 7, after which the levels returned to the normal ranges. In the negative control group, the testosterone levels were maintained within the normal ranges during the period. The LH levels of male rabbits were also determined with the same specimens collected from each group, the negative control group were found to stay more or less within normal ranges during the study period. The experimental groups were found to drop significantly by 40-80% from day 5 to day 9, upon the antibody injection.
In the case of high dose group, low LH levels were found between day 5 and day 9, whereas in the low dose group the suppressed LH levels were observed between day 3 and day 9. Compared to those of the negative control, the LH levels returned to normal ranges after day13 in all these study groups. Results of these comparative group studies were graphed with deviations and presented in Figure 8, including the statical analysis with typical student t-test methods.
| t-test values and p-values | ||||
|---|---|---|---|---|
| Day | neg. ctrl to 1 mg/kg | neg. ctrl to 3 mg/kg | ||
| t test | P value | t test | P value | |
| 3 | 2.965789 | 0.003072 | 4.2 | 0.0001 |
| 5 | 3.352632 | 0.000842 | 3.851228 | 0.000132 |
| 7 | 1.085873 | 0.281089 | 7.396226 | 0.0001 |
| 9 | 0.34386 | 0.743985 | 0.840546 | 0.407961 |
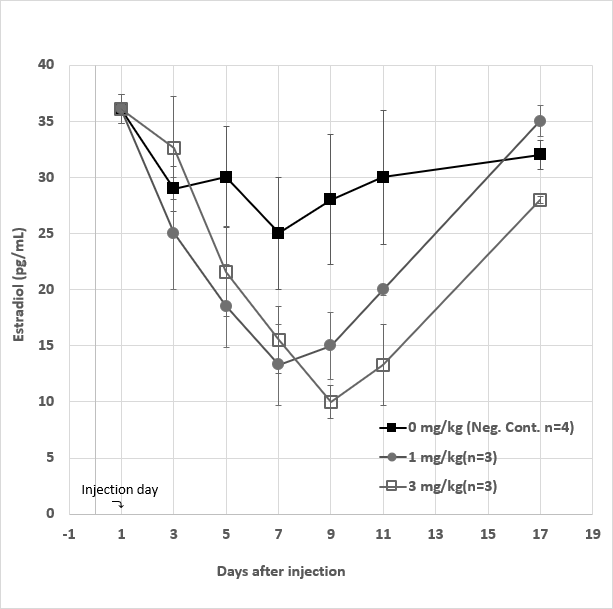
Figure 8: Serum LH levels from a total of 10 male rabbits divided into three experimental groups. The negative control groups (■) (n=4) received no injection on day 1, whereas each of the low dose (●) (n=3) group and high dose group (◻) (n=3) received 1 mg/kg and 3 mg/kg of GHR106 (hIgG4) injection subcutaneously, respectively. Serum LH levels was determined in each group starting on day 1 and ending on day 13. They were presented in each experimental group as means and deviations on the day indicated. Data presented are means and standard deviations of the negative control group (n=4) vs. the experimental groups (n=3 for each).
| t-test values and p-values | ||||
|---|---|---|---|---|
| Day | neg. ctrl to 1 mg/kg | neg. ctrl to 3 mg/kg | ||
| t test | P value | t test | P value | |
| 3 | 3.578947 | 0.000373 | 1.473684 | 0.140847 |
| 5 | 4.69828 | 0.0001 | 3.59633 | 0.00035 |
| 7 | 4.126316 | 0.0001 | 3.619575 | 0.000321 |
| 9 | 2.93633 | 0.003372 | 3.559959 | 0.0004 |
• Female Rabbits Experiments Ten female rabbits were also employed for “proof-of- concept” experiments. The experimental protocols including two doses of GHR106 injections and bleeding schedules are the same as those of the male rabbits. The group averages plus deviations of serum E2 levels from day 1 to day 17 are presented in Figure 9 for close comparisons. In the case of low dose group, the serum E2 levels decreased significantly by 40-60% from day 3 to day 9 after injection. The hormone levels Returned to the normal range after day 11.
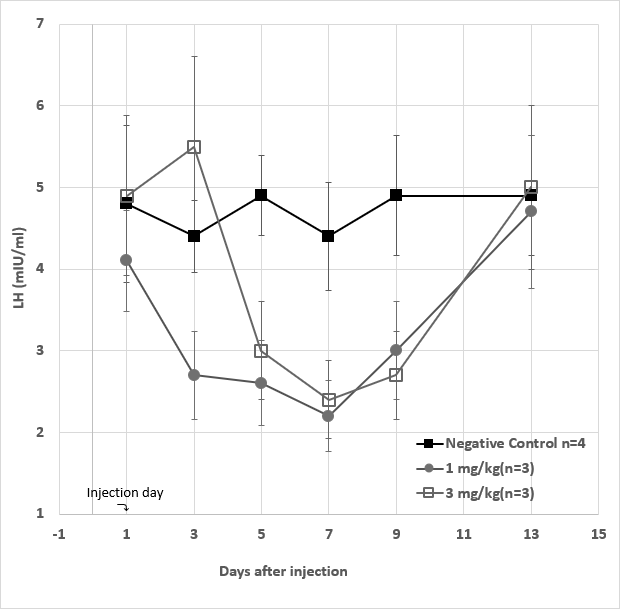
Figure 9: Serum estradiol (E2) levels for a total of 10 female rabbits divided into three experimental groups, which were maintained from day1 to day17. The negative control group (■) (n=4) received no injection on day 1, whereas each of the high dose group (◻) (n=3) and the low dose group (●) (n=3) received 3 mg/kg and 1 mg/kg injection of GHR106 (hIgG4), respectively on day 1. The serum E2 levels were determined starting with day 1 and ending on day 17, and presented at different time intervals during the study period. Data presented are means and standard deviations of the negative control group (n=4) vs. the experimental groups (n=3 for each).
In the case of high dose group, similar decreases in serum E2 levels (40-70%) were observed from day 3 to day 11 following antibody injection on day 1. On day 17, the serum E2 levels were shown to be in the normal range to all three experimental groups. By comparison, the average daily changes of serum E2 levels among the experiment groups are considered to be acceptable, including those of the negative control group. Therefore, among the injection groups, the decreases in serum E2 levels upon injection during the first week are considered statistically significant.
| t-test values and p-values | ||||
|---|---|---|---|---|
| Day | neg. ctrl to 1 mg/kg | neg. ctrl to 3 mg/kg | ||
| t test | P value | t test | P value | |
| 3 | 1.178947 | 0.240867 | 1.125359 | 0.263496 |
| 5 | 2.893453 | 0.003859 | 2.038885 | 0.041122 |
| 7 | 2.806854 | 0.005042 | 2.45 | 0.014212 |
| 9 | 3.047847 | 0.002359 | 5.087239 | 0.0001 |
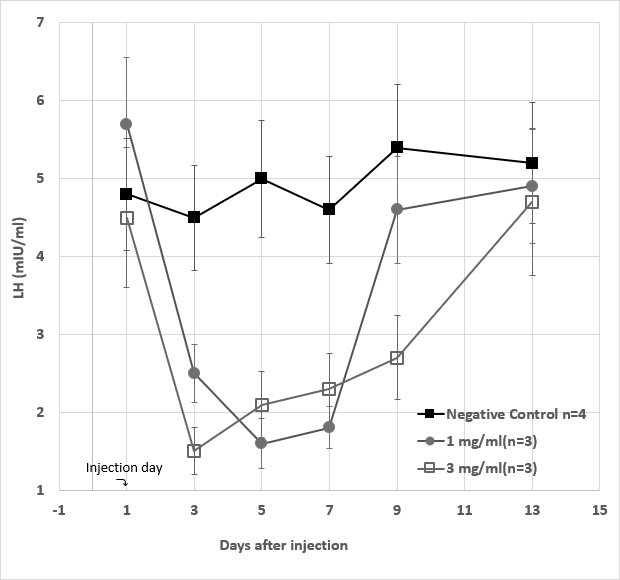
Figure 10: Serum LH levels from a total of 10 female rabbits divided into three experimental groups during the study period. The negative control groups (■) (n=3) received no injection on day 1. In the high dose (◻) (n=3) and the low dose (●) (n=3) groups, the rabbits in each group received 3 mg/kg and 1 mg/kg of GHR106 (hIgG4), respectively. On day 1, the serum LH levels of individual rabbit were determined with means plus deviations from each group are presented for comparisons. Data presented are means and standard deviations of the negative control group (n=4) vs. the experimental groups (n=3 for each).
| t-test values and p-values | ||||
|---|---|---|---|---|
| Day | neg. ctrl to 1 mg/kg | neg. ctrl to 3 mg/kg | ||
| t test | P value | t test | P value | |
| 3 | 3.929825 | 0.0001 | 6.348178 | 0.0001 |
| 5 | 6.555829 | 0.0001 | 5.11381 | 0.0001 |
| 7 | 6.017544 | 0.0001 | 4.126316 | 0.0001 |
| 9 | 1.100351 | 0.274547 | 4.126316 | 0.0001 |
To the same serum specimens collected from female rabbits in the experimental groups, LH levels were analyzed and presented in Figure 9 with standard deviations. Although LH levels in the negative control group are relatively constant during the study period, those in the injection groups are significantly reduced during the first week after day 1 antibody injection. In the low dose group, LH levels decreased to 50% and 30% of the normal control on day 3 and day 5, serum LH levels returned to normal after day 9. Both in the high dose and low dose groups, the time- dependent decreases in the hormone levels of treated rabbits are considered statically significant (Figures 7-10).
Estimation of Circulation Half-life of GHR106 (hlgG4) in Rabbits
Following the initial injection of GHR106 (hlgG4), the serum human lgG concentrations in rabbits were determined over a two-week time period. By using enzyme immunoassays specific to human lgG, the human lgG concentrations were determined. Based on the results of these analysis, it was estimated that the average circulation half-life of GHR106 (hlgG4) in rabbits is about 3 to 5 days.
Discussion
General assessment of Proof-of-Concept (P-o-C) experiments was performed in rabbits to demonstrate the time-dependant reversible suppressions and recoveries of reproductive hormones upon a single injection of GHR106 (hIgG4). As show in Table 1, there is a high degree of sequence homology between N1-29 oligopeptide of human GnRH receptor and that of the rabbit. Therefore, a humanized GHR106, GHR106 (hIgG4) generated against human GnRH receptor should cross-react with that of the rabbit, through its binding with N1-29 oligopeptide located in the extra cellular domains of GnRH receptor. This was demonstrated in binding ELISA studies as presented in Figure 2.
Single Rabbit Experiment
P-o-C studies were conducted in two stages. Initially, mature male and female rabbits were used to test if a single injection of GHR106 (hIgG4) in two different doses (high dose 3 mg/kg and low dose 1 mg/kg) to each rabbit, can cause reversible suppressions of reproductive hormones. Compared to pre-injections on day 1, serum levels of LH and testosterone of ale rabbits decreased significantly (60- 80%) starting on day 3 and returned to the normal ranges after a week or longer. Both high dose or low dose injected male rabbits revealed similar profiling changes of reproductive hormones (Figures 3 & 4).
In the case of female rabbits, a single GHR106 injection to a give one in either doses can also result in significant decreases in serum concentrations of LH and E2 during the initial one week to ten days after injections. The hormone levels returned to normal ranges when the drug effect is over after two weeks (Figures 5 & 6).
Multi-Rabbit Experiments
With these preliminary observations in single rabbit data, long scale rabbit experiments (n≥30) by including the negative controls were conducted with identical protocols. The data generated in each experimental group were analyzed statistically. Averages and deviations of hormone levels from rabbits in each experimental group are presented in Figures 7-10 for parallel comparisons. In general, both single rabbit and multi-rabbit experiments revealed reversible suppressions of reproductive hormones upon the single injection of GHR106 (hIgG4) with either 1 mg/kg or 3 mg/kg dose during the initial one- or two-weeks’ period. The hormone levels return to normal ranges as in the negative control group. In spite of significant variations of individual hormone data for individual rabbits in the experiment groups, reversible suppressions of reproductive hormone were clearly observed. Multi-rabbit experiments and single individual rabbit experiments led to the same conclusion in terms of time-dependent reversible hormones suppression upon single injection with GHHR16 (hIgG4). Therefore, it can be demonstrated that GHR106 (hIgG4) is an antibody-based long- acting GnRH antagonist when compared with GnRH antagonist such as Cetrorelix currently used clinically.
Conclusion
GHR106 monoclonal antibody was initially generated against N1-29 oligopeptide corresponding to the extracellular domains of human GnRH receptor [1, 2, 3, 4, 5, 6]. Human GnRH receptor is mainly located in the anterior pituitary and functions to regulate the release of gonadotropins including LH and FSH upon pulsatile stimulations of GnRH from hypothalamus [2, 3, 4].
High degrees of N1-29 oligopeptide sequence homology exist among humans and other animal species such as dog, cat, rabbit and monkey (Tables 1). One would expect that GHR106 and its antibody isoforms can interact with GnRH receptors from these animal species to induce reversible suppressions of reproductive hormones as reported in the present study [10, 11]. Therefore, GHR106 and its related antibody isoforms may have potential clinical applications in humans and other animal species in the areas of fertility regulation/control or diseases [12, 13, 14] including uterine fibroids endometrioses and female infertility [14, 15, 16, 17]. Human GnRH receptor was also found in other tissues such as gonads and cancer cells, with different mechanisms of actions from that of the pituitary receptor. Through autocrine/paracrine regulations, GnRH receptor in extra-pituitary tissues serves to induce apoptosis upon interactions with GnRH or its peptide derivatives and GHR106 [4, 5, 6, 7]. These characteristic interactions may suggest potential clinical applications of GHR106 similar to those of analogue of the known GnRH antagonists [14, 15].
Based on previous extensive investigations [1, 6, 7, 8, 9], GHR106 monoclonal antibody or its antibody isoforms should be considered as first-in-class of GnRH antagonist and is quite different from the other two classes of known GnRH antagonists [14, 15, 16], not only in molecular size, but also in structures [4]. Since there are numerous clinical applications of GnRH antagonists such as Elagolix [17] and decapeptide Cetrorelix [14, 17], additional further studies of GHR106- related GnRH antagonists are required to explore their full potential clinical applications [17, 18] not only in humans but also served domestic animal species [19, 20, 21, 22] such as dog and cat.
Disclosure Statement
The author reports no conflict of interest.
References
-
Lee CYG, Ho J, Chow SN, Yosojima K, Schwarz C, et al. (2000) Immunoidentification of gonadotropin-releasing hormone receptor in human sperm, pituitary and cancer cells. Am J Reprod Immunol 44(3): 170-177.
-
Williams BL, Akazome Y, Oka Y, Eisthen HL (2014) Dynamic evolution of the GnRH receptor gene family in Vertebrates. BMC Evol Biol 14: 215.
-
Millar RP (2005) GnRHs and GnRH receptors. Anim Reprod Sci 88: 5-28.
-
Limonta P, Moretti RM, Montagnani M, Morta M (2003) The biology of gonadotropin hormone- releasing hormone: role in control of tumor growth and progression in humans. Front Neuroendocrinol 24(4): 279-295.
-
Kakar SS, Jennes L (2005) Expression of gonadotropin- releasing hormone and gonadotropin- releasing hormone receptor mRNA in various non-reproductive tissues. Cancer Lett 98(1): 57-62.
-
Lee G, Ge B (2010) Growth inhibition of tumor cells in vitro by using monoclonal antibodies against gonadotropin-releasing hormone receptor. Cancer Immunol Immunothera 59(7): 1011-1019.
-
Chien CH, Chen CH, Lee CYG, Chang TC, Chen RT, et al. (2004) Detection of gonadotropin- releasing hormone receptor and its mRNA in primary epithelial ovarian cancer. Int J Gynecol Cancer 14: 451-458.
-
Weiss JM, Diedrich K, Ludwig M (2002) Gonadotropin- releasing hormone antagonists: Pharmacology and clinical use. Women Treat Endocrinology 4: 281-291.
-
Lee G, Huang CY, Ge B (2012) Two distinct humanized monoclonal antibodies for immunotherapy of ovarian cancer. J. Cancer Sci Thera 6: 110-116.
-
Crescloi S, Correa I, Karagiannis P, Davis AM, Sutten BJ, et al. (2016) IgG4 characteristics and functions in cancer immunology. Current Allergy Athma Res 16: 7-26.
-
Lee G, Chow SN, Chien CH and Liu S (2015) Anti-GnRH receptor monoclonal antibodies, First- in-class GnRH analog. J. Gynecol Research 1 (1): 102-111.
-
Engel JB, Anderbert A, Frydam R (2007) Presurgical Short terms treatment of uterine fibroids with different doses of Cetrorelix acetate: a double blind, placebo-controlled multicenter study. Eur J Obstet Gynecol Reprod Biol 134: 225-232.
-
Klotz L, Boccon-Gilbod L, Shore ND, Andreou C, Persson BE, et al. (2008) The efficacy and safety of degarelix: a 12-month, comparative, randomized open-label, parallel group phase III study in patients with prostate cancer. BJU Int 102(11): 1531-1538.
-
Reissmann T, Sehall AV, Bouchard Po, Riethmiller H, Engel J (2000) The LHRH antagonist Cetrorelix: a review. Hum Reprod Update 6(4): 322-331.
-
Copperman AB, Brnadifa C (2013) Optimal usage of GnRH antagonists: a revies of literature. Reprod Biol Endocrinol 110(20): 1-23.
-
Ezzatio MC, Bruce R (2015) Elagolia, a novel orally available GnRH antagonist under investigations for the treatment of endometriosis-related pain. Woman’s Health 11(1): 11-28.
-
Shore ND (2013) Enperience with degarlix in the treatment of prostate cancer. Ther Adv Urol 5(1): 11-24.
-
Coccia ME, Compuretto C, Braco GL, Scarcelli G (2004) Eur J Obs Gyn and Reprod Biol 1155: 544-556.
-
Corrada Y, Hermo D, Tortora M, Esquirel C, Gobello C (2005) Use of GnRH antagonist, Acyline, on pregnancy termination in the bitch, in “World small animal veterinary association world congress proceedings.
-
Gobello C (2012) Effects of GnRH antagonists vs. agonists in domestic carnivores, a review; Reprod. In domestic animals 47(36): 373-376.
-
Lucas X (2004) Clinical use of Deslorelin in companion animals, a review: Reprod in Domestic Animals 49(54): 64-71.
-
Herbert CA, Trigg TE (2005) Application of GnRH in the control and management of fertility animals. Animal Reprod. Science 88(1-2): 141-153.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework