Morphological Study and Description of Biacetabulum biloculoides Mackiewicz and Mccrae, 1965 (Cestoda: Caryophyllidae) From Catostomid Fishes in Lakes and Rivers of Wisconsin, With Special Notes on Concurrent Infections
Thirteen species in 8 genera of caryophyllid cestodes including Biacetabulum biloculoides Mackiewicz and McCrae, 1965 were found in Catostomus commersoni (Lacépède), Cyprinus carpio (Raf.), and Erimyzon sucetta (Lacépède) in 2 lakes and 2 rivers in SE Wisconsin and/or SW Lake Michigan. The morphology of the Wisconsin populations of B. biloculoides is revisited and missing information from the original description and ours were completed and all measurements compared. The posterior movement of Acanthocephalus dirus (Van Cleave, 1931) Van Cleave and Townsend, 1936 and the anterior movement of B. biloculoides and Glaridacris catostomi Cooper, 1920 in concurrent infections of the intestine of C. commersoni were discussed as a clear case of competitive exclusion. Light microscopy revealed new features especially of the scolex and reproductive structures.
Introduction
Mackiewicz [1] reviewed the morphology, anatomy, development, zoogeography, and systematics of the Caryophyllidea (Cestoidea) in 1972 and indicated the existence of 89 species and 37 genera in these monozoic cestodes. Ten years later, Mackiewicz [2] provided perspectives on the evolution, biology and zoogeography of 128 species in 45 genera of caryophyllids from the Nearctic, Neotropical, Palearctic, Ethiopian, Oriental, and Australian regions. Twelve years later, Mackiewicz [3] recognized about 150 species in 41 genera worldwide (except for the Neotropical Region). Of these, 14 genera and 90 species belonging to three families have been described from the Indomalayan region from catfishes (Siluriformes: Bagridae, Clariidae, Heteropneustidae, Schilbeidae and Siluridae), cyprinid, and cobitid fishes. A large number of inadequately described species from clariid and heteropneustid catfishes in India and neighboring countries have been invalidated [4, 5]. Most recently, however, Scholz, et al. [6] and Scholz and Kuchta [7] recognized only 117 species and 46 genera in order Caryophyllidea van Beneden in Carus, 1863 parasitizing teleost fish (Cypriniformes, Siluriformes, and some catfish). Chubb JC [8] reviewed the seasonal ecology of the Caryophyllidea in the various climate zone of the world.
The order consists of four families, namely the Balanotaeniidae, Capingentidae, Caryophyllaeidae and Lytocestidae [3]. Classification at the family level is based on the position of the inner longitudinal musculature in relation to the testes and vitelline follicles [1, 3]. “The caryophyllideans are exclusively intestinal parasites of freshwater teleosts with 85 species reported from cypriniform and 29 from siluriform fishes. Only few species of the Lytocestus with 2 species and monotypic Lytocestoides are known from mormyrids (Osteoglossiformes), characiform and cichlid (Perciforms) fishes, respectively. Within families, 40 % of caryophyllideans parasitize North American suckers (Catostomidae), 26 % cyprinids, 10 % clariid catfishes and just a few caryophyllideans have been reported from 12 additional fish families” [7]. Freshwater annelids of the family Naididae, especially species previously placed in the Tubificidae (Tubifex tubifex, Limnodrilus spp.), serve as the only intermediate hosts [1].
In Arizona and Wisconsin, Amin OM [9, 10, 11, 12] recognized 14 species in 8 families of caryophyllid cestodes from suckers (Catostomidae) in lake and river systems. These included Biacetabulum biloculoides Mackiewicz, et al. [13] from Catostomus commersoni (Lacépède) in SE Wisconsin. See collections, following, for details. The following is a list of the 14 species of cestodes collected organized by host species from Arizona (A) and Wisconsin (W) based on Amin [9, 10, 11].
Carpoides Cyprinus (LeSueur); quillback Rowardleus pennensis Mackiewicz and Deutsch, 1976 (W) Catostomus clarkii Baird and Girard; desert sucker Isoglaridacris hexacotyle (Linton, 1897) Mackiewicz, 1968 (= Monobothrium hexacotyle (Linton, 1897) Nybelin, 1922, Moghe, 1925; Caryophyllaeus hexacotyle (Linton, 1897) Woodland, 1923, 1926; Glaridacris hexacotyle (Linton, 1897) Hunter, 1927 (A) Catostomus commersoni (Lacépède); white sucker Biacetabulum biloculoides Mackiewicz and McCrae, 1965 (= Biacetabulum biloculoides McCrae, 1961 [nomen nudum]; Glaridacris catostomi Van Cleave and Mueller, 1934; Hugghins, 1958; F. Meyer, 1958) (W) Biacetabulum macrocephalum MacCrae, 1962 (W) Biacetabulum sp. Hunter, 1927 (W) Glaridacris catostomi cooper, 1920 (= Caryophyllaeus catostomi Woodland, 1923; Moghe, 1925; Woodland, 1926) (W) Glaridacris laruei (Lamont, 1921) Hunter, 1927 (Glaridacris intermedius Lyster, 1940 fide Mackiewicz, 1965) (W) Hunterrella nodulosa Mackiewicz and McCrae, 1962 (W) Isoglaridacris folius Fredrickson and Ulmer, 1967 (W) Monobothium hunteri Mackiewicz, 1963 (W) Catostomus insignis Baird and Girard; Sonora sucker Isoglaridacris hexacotyle (Linton, 1897) Mackiewicz, 1968 (A) Cyprinus carpio Linn.; common carp Atractolytocestus huronensis Anthony, 1958 (W) Khawia iowensis Calentine and Ulmer, 1961 (W) Erimyzon sucetta (Lacépède); lake chubsucker Glaridacris laruei (Lamont, 1921) Hunter, 1927 (W) Isoglaridacris agminis Williams and Rogers, 1972 (W) Isoglaridacris multivitellaria Amin, 1986 (W) Amin OM [10] described B. biloculoides from C. commersoni in Wisconsin and noted the effects of C. commersoni from the Pike River on worm burden as well as the effect of the parasite on host and site of infection in stomach pits (Figures 2&3) with up to 25 worms in 4X6X4 mm pits [14] compared to up to 37 and 46 worms per pit in New York and North Carolina [13]. Amin OM [14] also related annual variations in the abundance and development of B. biloculoides in Pike River suckers during the autumn of 1972 and 1973 to seasonal temperature. Additionally, specimens of B. biloculoides and G. catostomi were collected from C. commersoni in the Pike River during 1972 and 1973 for competitive exclusion studies in concurrent infections [15]. Mackiewicz and McCrae [13] originally described B. biloculoides from C. commersoni from Horsetooth Reservoir in Colorado and reported it from other waterbodies in Colorado, Iowa, New York, Pennsylvania, South Carolina, and South Dakota. Mitchum DL [16] briefly described it from the same host in a key to 6 caryophyllid genera from Wyoming. Molnar K [17] reported it from C. commersoni in Ontario and Buckner, et al. [18] added Carpiodes velifer (Rafinesque) as a new host in Indiana.
“Molecular phylogenetic studies confirm monophyly of caryophyllideans, but also reveal the non-monophyly of three of the four families (the family Balanotaeniidae is mono-generic)” [19, 20, 21]. In contrast, most genera seem to be monophyletic, with a few exceptions such as Glaridacris Cooper, 1920” [7] and [22].
Collections
In SE Wisconsin, 51 specimens (92% mature adults) were collected from 20% of 135 individuals of C. commersoni in the Pike River which flows in Lake Michigan, Racine and Kenosha counties (42°38′40″N,87°51′09″W) in September-October of 1972 [10]. Fifty-four additional specimens were similarly collected from C. commersoni in the Pike River in November, 1973 [14]. Twenty-three additional worms similarly infected 3 C. commersoni from the Pike River during July, 1974 [14]. “One juvenile and one gravid adult were recovered from the stomach of a 36-cm- long female C. commersoni collected from Tichigan Lake, a relatively large eutrophic impoundment on the Fox River which is a tributary of the Mississippi River in Racine County (42°48′20″N,88°13′00″W) during the autumn. The same fish also harbored one gravid Hunterella nodulosa Mackiewicz and McCrae, 1962, three-juvenile Glaridacris catostomi Cooper, 1920, and five juvenile Biacetabulum sp. in the stomach and post-gastric region” [11]. A total of 136 specimens of B. biloculoides and G. catostomi were collected from C. commersoni in the Pike River during 1972 and 1973 for competitive exclusion studied in concurrent infections [15]. White suckers from the nearby Root River in Milwaukee and Racine counties (42° 44’ 1” N-87° 46’ 42”
W) were moderately infected with B. biloculoides in the fall of 1974 and some were concurrently infected with specimens of Hunterella nodulosa Mackiewicz and MacCrae, 1962 [13] and Biacetabulum macrocephalum McCrae, 1962 (miscellaneous). Mackiewicz and MacCrae, 1962 [13] and twelve more specimens of 80 newly examined specimens from all collections were used for microscopy.
Methods
Worms were initially stored in cold water then fixed in cold 70% ethanol. They were subsequently punctured with a fine needle then stained in Semichon’s carmine, destained in 4% hydrochloric acid in 70% ethanol, dehydrated in ascending concentrations of ethanol and cleared in 100% xylene. Whole worms were mounted in Canada balsam. Measurements are in micrometers and mm as stated; the range is followed by the mean values between parentheses. Width measurements represent maximum width. Microscope images were created using 10X and 40X objective lenses of a BH2 light Olympus microscope (Olympus Optical Co., Osachi-shibamiya, Okaya, Nagano, Japan) attached to an Am Scope 1000 video camera (United Scope LLC, dba AmScope, Irvine, California), linked to an ASUS lab top equipped with HDMI high-definition multimedia interface system (Taiwan- USA, Fremont, California). Images from the microscope are transferred from the lab top to a USB and stored for subsequent processing on a computer.
Results
Amin OM [10] provided a brief description of B. biloculoides based on 50 adults collected from C. commersoni in the autumn of 1972 from the Pike River, SE Wisconsin for the first time since its original description by Mackiewicz and McCrae [13] from the same host species in Colorado, Iowa, New York, North Carolina, Ohio, Pennsylvania, South Carolina, and South Dakota. We have reexamined an additional sample of 50 adults collected from the same host species in the autumn of 1971 from the Root River in 1971. Measurements from both collections are combined (Table 1).
Specimens of B. biloculoides from C. commersoni were common in the Pike River than the Root River systems, rarely present in Tichigan Lake and totally absent in Silver Lake.
Our new observations, 57 years after the original description of that species, were generally in agreement with our brief description and comparable to that of Mackiewicz and McCrae [13] with some variations. Most worms were recovered from non-perforated stomach pits that appeared as cysts on the serosal surface. Up to 4 worms per stomach pit were noted from suckers in the Pike River. Annual cycles in natural infections appear to be involved in the different proportion of immature worms in the infra-populations and their numbers and distribution in stomach pits from year to year [14].
| Mackiewicz & McCrae (1965) | Amin (1974) & this paper) | |
|---|---|---|
| Character | Colorado & New York | Pike & Root rivers, Wisconsin |
| N | 39 | 50 + 50 |
| Total length (mm) | 3.70-11.00 | 2.00-10.25 (5.20) |
| Max. W at gonopore (mm) | 0.40-1.00 | 0.47-1.76 (0.88) |
| Scolex W (mm) | --- | 0.35-0.84 (0.54) |
| Neck W (mm) | --- | 0.24-0.60 (0.44) |
| Testes no. | 50-90 (61) | 64-95 (80) |
| Testes max. diameter (mm) | 0.08-0.18 corrected | 0.06-0.20 (0.15) |
| Cirrus sac diameter (mm) | 0.14-0.24 | 0.16-0.23 (0.20) |
| Vitellaria diameter (mm) | 0.08-0.17 | 0.13-0.22 (0.15) |
| Ovary arm L (mm) | 0.20-0.50 | 0.23-0.55 (0.39) |
| Ovary arm W (mm) | --- | 0.19-0.38 (0.28) |
| Eggs in utero L X W (μm) | 50-55 X 27-30 | 45-55 (51) X 32-40 (36) |
Table 1: Comparative measurements of mature Biacetabulum biloculoides from Catostomus commersoni collected from the Pike and Root
We provide below a brief revised account of our qualitative morphological findings with emphasis on first reported variations in the scolex and some reproductive structures (Figures 1-12) and a comparative morphometrical data for mature adults (Table 1) high-lightening variations from, and additions to our earlier account Amin OM [10] and that of Mackiewicz and McCrae [13]. Considerably smaller immature worms measuring 1.12-1.88 mm (mean 1.41) long, 0.36-0.0.68 mm (0.48) wide at gonopore, 0.24-0.56 mm (0.36) wide at scolex, and 0.16-0.44 mm (0.28) wide at neck, with undefined scolex are more common especially in younger fish. Worms were more prevalent, more mature, and larger in larger hosts [10, 11]. Observations on new specimens show that testes develop first then vitelline glands appear next in 1 row per each of 2 lateral columns (Figures 1 & 8).
Morphological Description of Adults
Body of small to medium length, dorso-ventrally flattened, with an outer layer of cortical parenchyma surrounding internal medullary parenchyma containing reproductive, muscular, and excretory systems (Figures 1-12). For measurements, see Table 1. Developing adults with unremarkable, lancet-shaped, or elongate-ovoid to bulbous, with 2 shallow depressions (loculi) lacking muscular septa and almost impossible to detect in mounted preparations (Figures 1-7). Neck, narrowest part of body, not especially differentiated. Three pairs of inner longitudinal muscles well developed Figures 4, 6 & 7; arrows); outer longitudinal muscles not evident. Vitelline glands usually round, about as large as testes, in 2 regions; the first beginning just posterior to neck anteriorly (Figure 8) and extending in 2 lateral bands and also sporadically surrounding testes centrally reaching cirrus (Figure 9), and the second region in a separate usually V-shaped cluster posterior to ovary (post ovarian vitellaria) (Figure 9). Less than 100 somewhat rounded testes in medullary region occupy central space beginning posterior to first vitelline gland (Figure 8) and usually develop sooner than vitellaria. Cirrus pouch eversible, round, highly muscular, with external seminal vesicle (Figures 11 & 12). Lobate ovary arms, compact, rounded, H-shaped, with robust short straight commissure Figures 9 & 10). Pre- and post-ovarian uterine coils (Figure 10) prominent leading to vagina (Figures 11, arrow) draining in common gonopore (Figure 11). Excretory system with a number of canals in body leading to terminal pore posteriorly. Operculate eggs with smooth surface (Figure 10). Specimens deposited: US National Museum Helminthol. Coll. No. 78811.
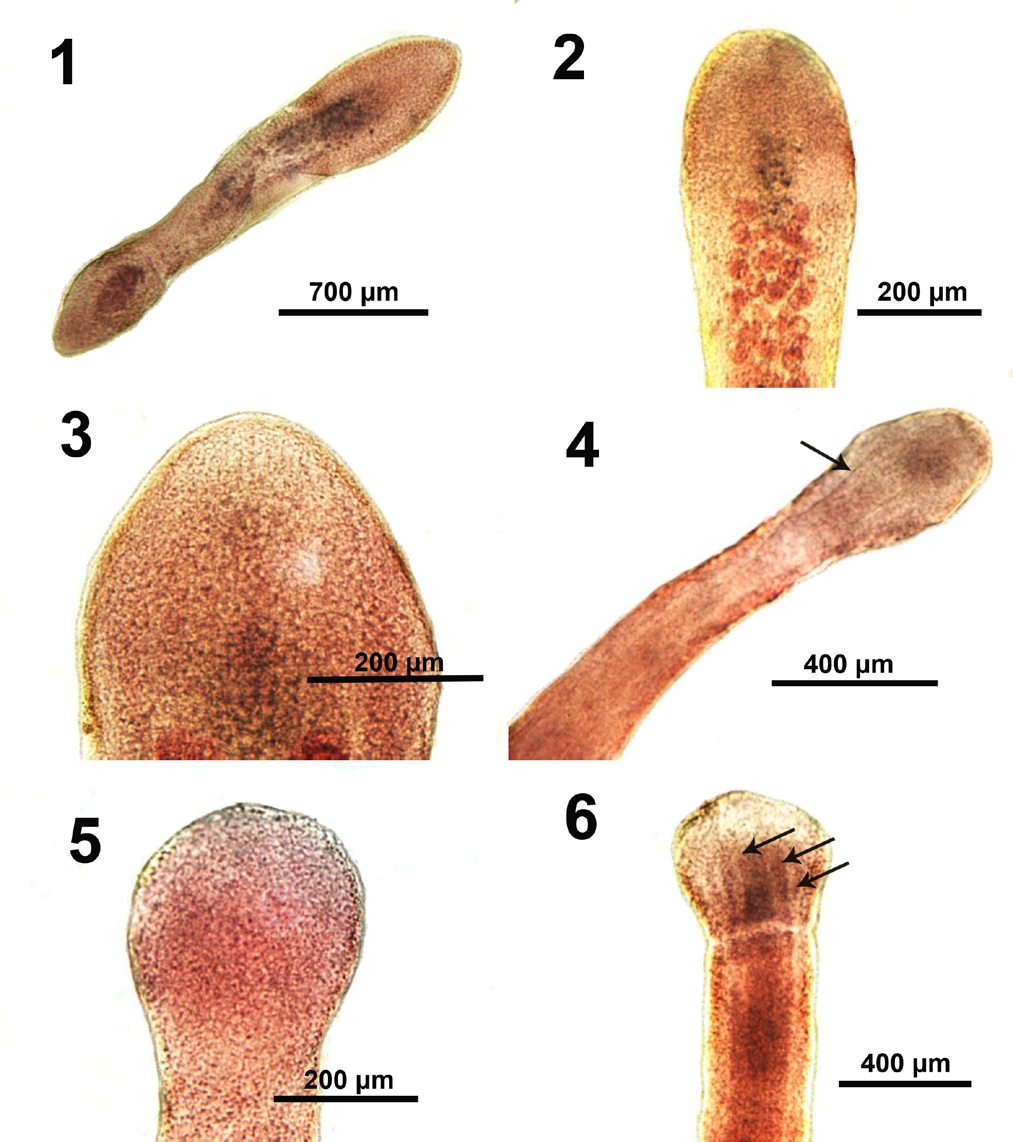
Figures 1-6: Microscope images of immature and adult specimens of Biacetabulum biloculoides in subsequent stages of development from Catostomus commersoni collected from the Pike and Root rivers, Wisconsin.
- An immature specimen with a lancelet-like scolex, incipient testes and vitellaria, and developing reproductive organs.
- The anterior end of a young adult showing a slightly more rounded apical end and the early emergence of only testes; vitellaria are not developed yet at that stage.
- The slightly more pointed scolex of this slightly more mature specimen.
- A slightly more developed specimen showing post-scolex constriction (neck) and inner longitudinal muscle fiber (arrow).
- In the next stage, the scolex becomes more rounded, still with no evidence of development of loculi.
- The scolex and anterior pre-testicular portion of another worm in a more advanced stage of development showing the delineation of a definite neck and 3 pairs of inner longitudinal muscle fibers (arrows).
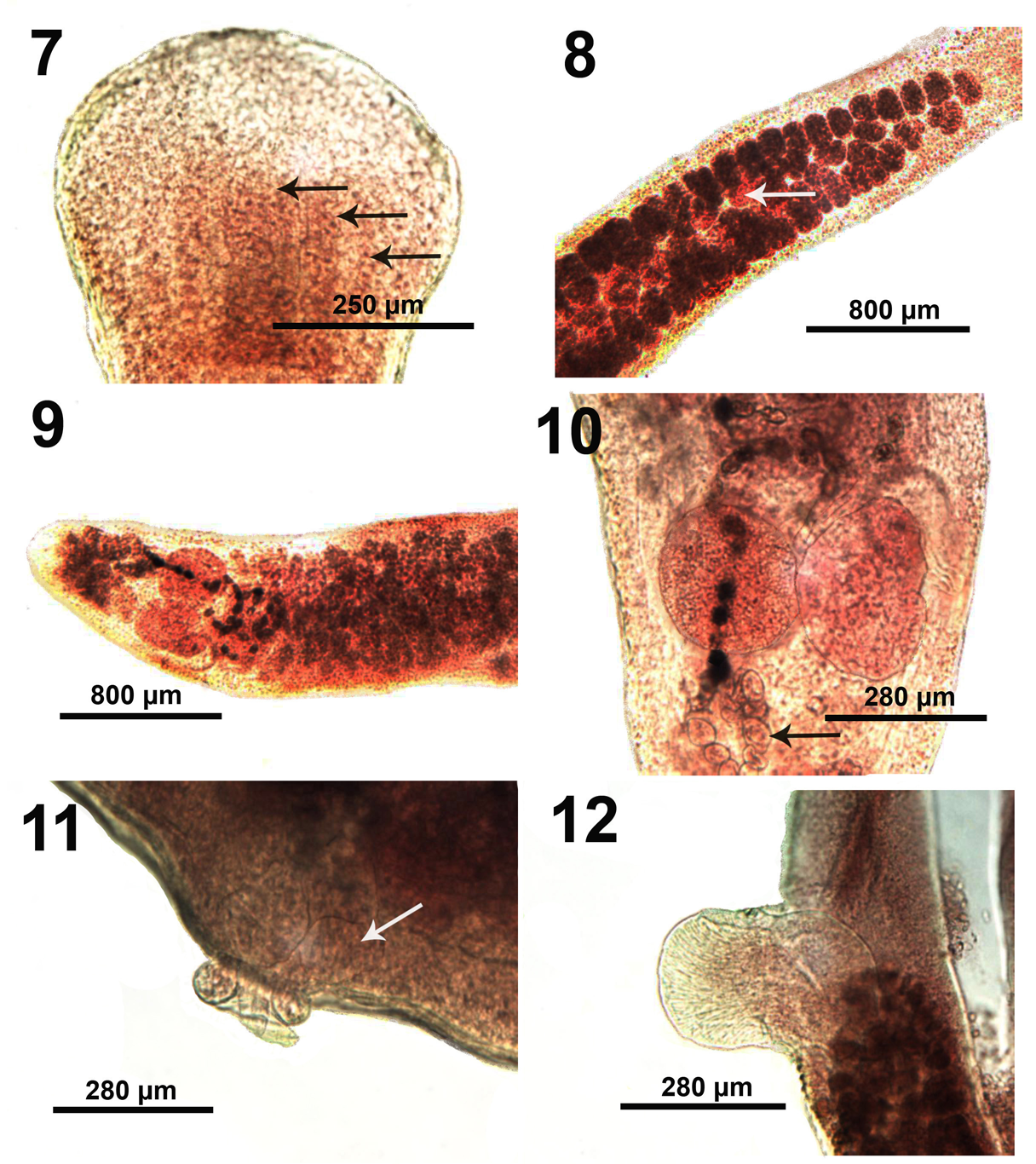
Figures 7-12: Microscope images of adult specimens of Biacetabulum biloculoides from Catostomus commersoni collected from the Pike and Root rivers, Wisconsin. 7. A near fully developed scolex showing the anterior reach of the inner longitudinal muscle fibers (arrows). 8. The anterior post-scolex half of a mature specimen showing the central lighter colored testes (arrow) encircled by the darker vitelline glands, especially laterally. 9. The posterior part of the same specimens in Figure 8 showing the shape and size of the ovarian lobes and the post-ovarian vitellaria. 10. A higher magnification of the ovarian lobes of the specimens in Figure 9 showing eggs in the pre- and post-ovarian uterus (arrow). 11. Ejaculatory duct extruded from the partially evaginated cirrus sac in a specimen also showing the vagina (arrow) joining the male system in common gonopore. 12. An extruded highly muscular cirrus sac about as wide as the diameter of the posterior body of this specimen.
Concurrent Infections
Amin OM [14] demonstrated the effect of concurrent infections on habitat isolation in the gut of 25 breeding C. commersoni from the Pike River in the spring of 1972 and 1973. This information has been corroborated and expanded from later collections (unpublished). Initially, 14 (56%) and 11 (44%) of suckers were infected with Acanthocephalus dirus (Van Cleave, 1931) Van Cleave and Townsend, 1936 singly and with A. dirus and B. biloculoides and G. catostomi concurrently, respectively. No fish was singly infected with cestodes but in a later collection in November, 3 of 45 examined suckers were singly infected with 29 specimens of B. biloculoides with site preference similar to that in concurrent infections. Competitive exclusion was demonstrated by the posterior movement of acanthocephalans and anterior movement of the cestodes in concurrent infections.
Misidentifications
Biacetabulum biloculoides can be readily misidentified because of similarities to other closely related species. It is closely related to other species of Biacetabulum in North America except for the absence of acetabular suckers. Mackiewicz and McCrae [13] resisted the temptation of erecting a new genus of caryophyllids based on this character alone. The identity of B. biloculoides has occasionally been confused with that of G. catostomi. Whole or parts of 18 specimens originally found in mucosal pits of C. commersoni from Oneida Lake were originally determined to be G. catostomi by Van Cleave and Mueller [23] but were later re-identified as B. biloculoides based on the examination of the original slides by Mackiewicz and McCrae. “Similarly, the G. catostomi records of Hugghins EJ [24] and Meyer FP [25] (were) redetermined as B. biloculoides on the basis of a restudy of the original material”. It is understandable that these misidentifications were made before B. biloculoides was described. However, considering the obvious differences in the scolex anatomy and the presence or lack of acetabular loculi, it is still not understandable to confuse the G. catostomi with any other form totally lacking acetabular suckers.
Discussion
Morphology
The morphology of specimens from both of our Wisconsin collections from C. commersoni was comparable to that of Mackiewicz and McCrae [13] from Colorado and New York with a few exceptions. Comparative measurements (Table 1) show that our Wisconsin specimens were about the same length but reached greater width at gonopore. Missing measurements from the original description made it hard to compare those of the scolex, neck, and width of ovary arms. We include the following new information. Scolex is scarcely detectable and undifferentiated from wider neck in young immature specimens which contained only 1 row of lateral pre-ovarian vitellaria, embryonic testes, and 1 cluster of female reproductive primordia with scarce post-ovarian vitellaria (Figure 1). The same scolex-neck morphology remains in larger immatures with better developed testes (Figures 2 & 3). The scolex develops further into ovoid then globular form in maturing adults (Figures 4-7). During this process, up to 3 pairs of inner longitudinal muscles become apparent. Outer longitudinal muscles are inapparent or absent and Mackiewicz and McCrae [13] noted that “Inner longitudinal muscles well developed.” They did not number the muscles but their line drawings nos. 5, 9, 14 showed 2 muscle fibers per worm.
Concurrent Infections
“The occupation of 2 ecologically distinct intestinal regions by the posterior movement of A. dirus and the anterior movement of B. biloculoides (and G. catostomi) in concurrent infections demonstrates a clear case of competitive exclusion”. Competition may have been for physical space and or biochemical (feeding) factors. The severity of interspecific competition would be reduced by habitat isolation. Cross SX [26] explored the relationship between infections of bloater (ciscoe) Coregonus hoyi Milner in Silver Lake, northern Wisconsin with Neoechinorhynchus species sp. in mid-intestine and Proteocephalus exiguus La Rue, 1911 in pyloric ceca. Cross SX [26] noted that hosts infected with 15 or more acanthocephalans contained a maximum of 4 cestodes while those infected with 25 or more cestode usually had no acanthocephalans and suggested that “nonspecific immunity” limits 1 parasite when the other is present in large numbers. Schad GA [27] introduced “non-specific cross immunity” to interpret Cross’s SX [26] observations suggesting that first arrivals stimulated host antibody production effective against the other parasite. Chappell LH [28] described spatial competitive exclusion between Proteocephalus filicollis (Rudolphi, 1802) Weinland, 1858 and Neoechinoehynchus rutuli (Müller, 1780) in natural concurrent infections of the 3-spined stickleback Gasterosteus aculeatus Linn. in England. The biochemistry in the intestinal tract of fish may constitute another variable regulating the spatial distribution and longitudinal movement of helminth parasites in the gut. Uglem and Beck [29] correlated habitat specificity with aminopeptidase activity (APase) in acanthocephalans Neoechinorhynchus cristatus Lynch, 1936 and N. crassus Van Cleave, 1919 situated in the posterior and anterior intestines of the Large- scale sucker Catostomus macrocheilus (Girard) in Idaho and Washington. Interest in the various aspects of concurrent infections has been addressed by other earlier observers including Beck [30], Crompton DWT [31], Holmes JC [32, 33, 34], Milne JC [35], Read and Phifer [36] and Thomas JD [37]. Studies of the longitudinal ecological isolation of helminth species along the digestive tract of fish have declined since. A more recent study [37] of concurrent infections in the rabbitfish Siganus rivulatus Forsskål (Siganidae) from the Red Sea at Sharm El-Sheikh, Egypt noted the separation of the acanthocephalan Sclerocollum rubrimaris Schmidt and Paperna, 1978 (Cavisomidae), the trematode Gyliauchen volubilis Nagaty, 1956 (Gyliauchenidae), and the nematode Procamallanus elatensis Fusco and Overstreet, 1979 (Cucullanidae) in anterior, middle, and posterior intestinal sites, respectively. Fundamental niches in single infections were affected by interactive site segregation into realized niches in concurrent infections exhibiting strong intraspecific aggregation patterns triggered by competitive interactions.
Acknowledgments
This project was supported by an Institutional Grant from the Parasitology Center, Inc. (PCI), Scottsdale, Arizona.
Declarations
Compliance with Ethical Standards
- Conflict of Interest: The authors declare no conflicts of interest or competing interests.
- Ethical Approval: The authors declare that they have observed all applicable ethical standards.
- Availability of Data: All presented and related data are available by contacting the senior author.
References
-
Mackiewicz JS (1972) Caryophyllidea (Cestoidea): a review. Experim Parasitol 31: 417-512.
-
Mackiewicz JS (1982) Caryophyllidea (Cestoidea): Perspect Parasitol 84: 397-417.
-
Mackiewicz JS (1994) Order Caryophyllidea van Beneden in Carus, 1863. In: LF Khalil, A Jones, and RA Bray (eds), Keys to the cestode parasites of vertebrates. CAB Intern Wallingford UK, pp: 21-43.
-
Ash A, Scholz T, Oros M, Kar PK (2011) Tapeworms (Cestoda: Caryophyllidea), parasites of _Clarias batrachus_ (Pisces: Siluriformes) in the Indomalayan region. J Parasitol 97(3): 435-459.
-
Ash A, Scholz T, Oros M, Levron C, Kar PK (2011b). Cestodes (Caryophyllidea) of the stinging catfish _Heteropneustes fossilis_ (Siluriformes: Heteropneustidae) from Asia. J Parasitol 97(5): 899-907.
-
Scholz T, Kuchta R, Oros M (2021a) Tapeworms as pathogens of fish: A rev J Fish Dis pp: 1-18
-
Scholz T, Kuchta R (2017) A digest of fish tapeworms. Vie et milieu - Life and environment 67(2): 43-58.
-
Chubb JC (1982) Seasonal occurrence of helminths in freshwater fishes. Part IV. Adult Cestoda, Nematoda and Acanthocephala. Adv parasitol 20: 1-292.
-
Amin OM (1969) Helminth fauna of suckers (Catostomidae) of the Gila River System, Arizona. II. Five parasites from _Catostomus_ spp. Am Midland Nat 82: 429- 443.
-
Amin OM (1974) Intestinal helminthes of the white sucker, _Catostomus commersoni_ (Lacépède), in SE Wisconsin. Proc Helminth Soc Wash 41: 81-88.
-
Amin OM (1986) Caryophyllaiedae (Cestoda) from lake fishes in Wisconsin with a description of _Isoglaridacris multivitellaria_ sp. n. from _Erimyzon sucetta_ (Catostomidae). Proc Helminthol Soc Wash 53:48-58.
-
Amin OM (1968) Helminth fauna of Suckers (Catostomidae) of the Gila River System, Arizona. PhD Thesis Ariz St Univ 202 pp (No 68-1644) Univ Microfilms Ann Arbor Michigan; Dissert Abstr 28: 3521.
-
Mackiewicz JS, McCrae RC (1965) _Biacetabulum_ _biloculoides_ n. sp. (Cestodea: Caryophyllaeidae) from _Catostotomus commersoni_ (Lacépède) in ‘ North America. Proc Helm Soc Wash 32: 225-228.
-
Amin OM (1975a) Intestinal helminthes of some southeastern Wisconsin fishes. Proc Helminthol Soc Wash 42: 43-46.
-
Amin OM (1975b) Host and seasonal associations of _Acanthocephalus parksidei_ Amin, 1974 (Acanthocephala: Echinorhynchidae) in Wisconsin fishes. J Parasitol 61: 318-329.
-
Mitchum DL (1995) Parasites of fishes in Wyoming. 1995. Wyoming Game and Fish Dept Cheyenne pp: 304.
-
Molnar K (1974) Parasites of fishes from Laurel Creek, Ontario. J Fish Biol 6(6): 717-728.
-
Buckner RL, Denner MW, Brooks DR, Buckner SC (1985) Parasitic endohelminths from fishes of Southern Indiana. Ind Acad of Sc Monogr 94: 615-620.
-
Olson PD, Scholz T, Poddubnaya LG, Littlewood DTJ (2008) On the derived position of _Archigetes_ and the early evolution of the tapeworms (Platyhelminthes: Cestoda). J Parasitol 94: 898-904.
-
Brabec J, Scholz T, Králová-Hromadová I, Baszalovicsová E, Olson PD (2012) Substitution saturation and nuclear paralogs of commonly employed phylogenetic markers in the Caryophyllidea, an unusual group of non- segmented tapeworms (Platyhelminthes). Int J Parasitol 42: 259-267.
-
Scholz T, Oros M (2017) Caryophyllidea. In: Caira JN, Jensen K, et al. (Eds.), Planetary Biodiversity Inventory (2008-2017): Tapeworms from Vertebrate Bowels of the Earth. Univ Kans, Nat Hist Mus, Spec Publ No 25, Lawrence, Kansas, pp: 47-64.
-
Scholz T, Waeschenbach A, Oros M, Brabec J, Littlewood DTJ (2021) Phylogenetic reconstruction of early diverging tapeworms (Cestoda: Caryophyllidea) reveals ancient radiations in vertebrate hosts and biogeographic regions, Intern J Parasitol 51(4): 236-277.
-
Van Cleave HJ, Mueller JF (1934) Parasites of Oneida Lake fishes. Part III. A biological and ecological survey of the worm parasites. Roosevelt Wildl An 3: 161-334.
-
Huggins EJ (1959) Parasites of fishes in South Dakota. Agr Expt Sta SD State College and SD Dept Game Fish and Parks Bull No 484, pp: 73.
-
Meyer FP (1958) Helminths of fishes from Trumbull Lake, Clay County, Iowa. Proc Iowa Acad Sci 65: 477-516.
-
Cross SX (1934) A probable case of non-specific immunity between two parasites of ciscoes of the Trout Lake region of northern Wisconsin. J Parasit 20(4): 244-
-
Schad GA (1966) Immunity, competition and natural regulation of helminth populations. Am Nat 100: 359- 364.
-
Chappell LH (1969) Competitive exclusion between the two intestinal parasites of the three-spined stickleback, _Gasterosteus aculeatus_ L J Paras it 55: 775-778.
-
Uglem GL, Beck SM (1972) Habitat specificity and correlated aminopeptidase activity in the acanthocephalans _Neoechinorhynchus cristatus_ and _N._ _crassus._ J Parasit 58: 911-920.
-
Beck JW (1951) Effect of diet upon singly established _Hymenolepis diminuta_ in rats. Exp Parasit 1: 46-59.
-
Crompton DWT (1970) An Ecological Approach to Acanthocephalan Physiology. Cambridge Univ Press, pp: 125.
-
Holmes JC (1961) Effects of concurrent infections on _Hymenolepis diminuta_ (Cestoda) and _Moniliformis dubius_ (Acanthocephala). I. General effects and comparison with crowding. J Parasit 47: 209-216.
-
Holmes JC (1962a) Effects of concurrent infections on _Hymenolepis diminuta_ (Cestoda) and _Moniliformis dubius_ (Acanthocephala). II. Effects on growth. J Parasit 48(3): 87-96.
-
Holmes JC (1962b) Effects of concurrent infections on _Hymenolepis diminuta_ (Cestoda) and _Moniliformis dubius_ (Acanthocephala). III. Effects in hamsters. J Parasit 48: 97-100.
-
Milne A (1961) Definition of competition among animals. Symp Soc Exp Biol 15: 40-61.
-
Read CP, Phifer K (1959) The role of carbohydrates in the biology of cestodes. VII. Interactions between individual tapeworms of the same and different species. Exp Parasit 8: 46-50.
-
Hassasanine RM El-S, Al-Jahdali MO (2007) Ecological comments on the intestinal helminths of the rabbitfish _Siganus rivulatus_ (Teleostei, Siganidae) from the northern Red Sea. Act Parasitol 52: 278-285.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework