Morphological Description of the Early Life Stages of Brachyplatystoma rousseauxii (Castelnau, 1855), a Large Migratory Catfish from the Amazon
The early development of Brachyplatystoma rousseauxii from the Amazon River was described through morphological, meristic and morphometric features, providing useful characters for identification of the species. The specimens of B. rousseauxii analyzed came from samples taken middle Solimões River, in the state of Amazonas, Brazil. The larvae were collected by horizontal trawls on the subsurface of the water column, with a plankton net (300 μm). The larvae and juveniles of B. rousseauxii showed a standard length ranging from 12.68 to 29.29 mm. The specimens were characterized by having a long snout (>40% of CC), small eyes (<10% of CC), anal opening reaching the mid-section of the body (≥50% of CP), a pair of barbels maxillary and two mentonian barbels, both not exceeding the anal fin, two spines on the dorsal fin, presence of filaments in the caudal fin and total myomeres with 46 segments (19 pre-anal and 27 post-anal). Number of fin segments corresponded to: pectoral= I+9, pelvic= I+5, dorsal= II+6 and anal=19. We expect that this study will expand the knowledge about the developmental biology of fish species in the Neotropical region.
Introduction
The gilded catfish or “dourada”, Brachyplatystoma rousseauxii (Castelnau, 1855), is a long- distance migratory catfish (Siluriformes) that can reach over 1.7 meters in total length. Morphologically, it has a flattened head, golden/silver colored body, and the presence of short barbels in the jaws, being considered a carnivorous fish and predator at the top of the food chain, with distribution recorded in the rivers of the Amazon, Orinoco and French Guiana river basins [1, 2, 3]. This species is known for carrying out one of the longest reproductive migrations among freshwater fish in the world, between growing and nursery areas in the Amazon estuary to spawning sites in the headwaters of tributaries of the Solimões/Amazonas River in Colombia, Peru, and Bolivia [4, 5, 6].
Brachyplatystoma rousseauxii is one of the most important and exploited fishing resources in the channel of the main white water rivers of the Brazilian Amazon, and captured by the commercial and artisanal fleet from Pucallpa, in Peru, to the region close to Belém, with thousands of families directly and/or indirectly from the fishing and commercialization of this catfish throughout the region [7, 8, 9, 10]. However, environmental changes such as deforestation, mining, and the building of hydroelectric power plant dams along numerous tributaries of the Amazon basin compromise the upward migration of adult fish and the downward migration of eggs and larvae. These modifications impact the ichthyofauna with a reduction in the recruitment population of the species, mainly of long-distance migrants [11, 12, 13]. These landscape changes can negatively affect the diversity in capture and fisheries production in the main Amazonian rivers [14].
In this context, due to the changes in river ecosystems and the ecological and economic relevance of B. rousseauxii, it is necessary to know the aspects of the initial ontogeny of this specie. Because the morphological, morphometric, and meristic information throughout the initial development is essential to identify the larvae collected in nature. In addition, assist ichthyoplankton inventory reports and subsidies of species management actions [15, 16, 17]. Therefore, this study aimed to describe the larval stages and juvenile period of B. rousseauxii captured in drift in the middle Solimões River, Amazonas, Brazil, using morphological and morphometric characteristics.
Materials and Methods
The specimens of B. rousseauxii analyzed came from samples taken middle Solimões River, in the state of Amazonas, Brazil. The larvae were collected by horizontal trawls on the subsurface of the water column, with a plankton net (300 μm). The license for the collection of biological samples was granted by SISBIO/ICMBio/ MMA, in the special license 23741-1, issued in accordance with Normative Instruction 154/2007 and followed the euthanasia protocols in accordance with the norms of the Conselho Nacional de Controle e Experimentação Animal [18]. After capture, the larvae were euthanized and fixed in a 10% formalin solution buffered with calcium carbonate. The B. rousseauxii specimens were identified through the regressive developmental sequence, which consists of the morphological comparison of juveniles to smaller stages. The descarne process (removal of tissues, muscles, and organs) was carried out on adult specimens of B. rousseauxii (Figure 1) to obtain vertebrae count data and, consequently, myomeres to assist in the identification of larvae and juveniles. Subsequently, the individuals were classified according to the developmental period: larval (preflexion, flexion, postflexion) and juvenile, as suggested by Ahlstrom EH, et al. [19], modified by Nakatani K, et al. [20].

For the analysis of morphometric relationships, standard length (SL), head length (HL), head width (HW), snout length (SnL), eye diameter (ED), interorbital distance (ID), head depth (HD), body depth (BD), snout-anus distance (StA), snout-dorsal fin distance (SnD), snout-anal fin distance (SnA), snout-pelvic fin distance (SnV) and snout- pectoral fin distance (SnP) were measured using a binocular stereomicroscope (Leica S9i) coupled with an integrated color digital camera for image capture and software analysis (Leica LAS EZ - Heerbrugg, Switzerland). The morphometry of SnL, HD, HW, ID, and ED are presented as percentage of HL, the other variables are presented as percentage of SL. Meristic counts of pre, postanal and total number of myomeres, spines and rays of dorsal, anal, pectoral and pelvic fins were performed. The larvae and juveniles used in this study are stored in the Collection of Ovos e Larvas de Peixes (<https:// specieslink.net/col/CROLP-LEIPAI/>) of Laboratório de Ecologia do Ictioplâncton e Pesca em Águas Interiores (CROLP-LEIPAI) of Universidade Federal do Oeste do Pará (UFOPA). The catalog numbers of examined specimens are as follows: LEIPAI00472, LEIPAI00473, LEIPAI00474, LEIPAI00475, LEIPAI00476, and LEIPAI00477.
Results and Discussion Larval development
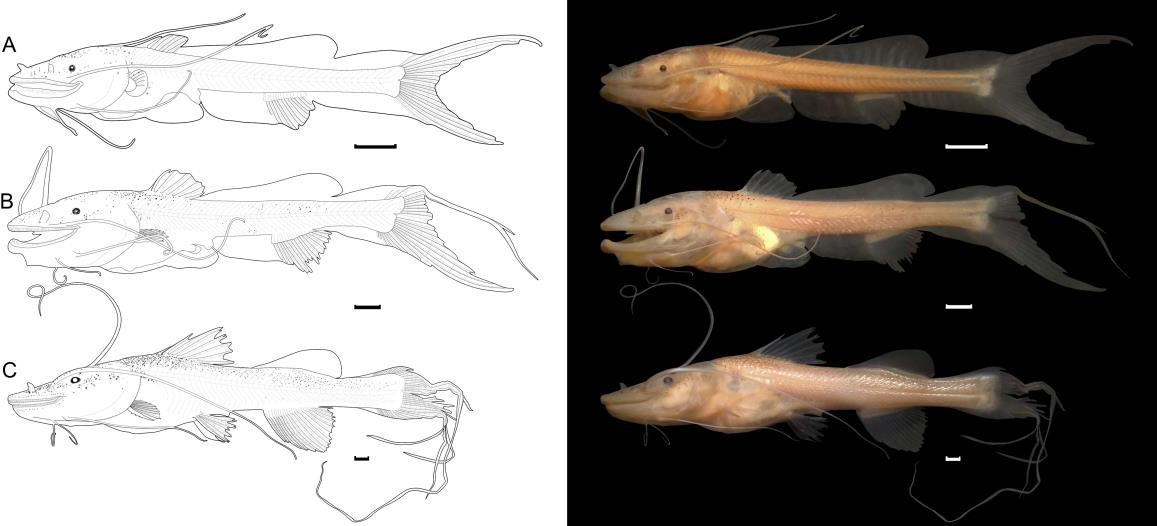
Flexion (Figure 2A): the specimen had a standard length of 12.68 mm. The final section of the notochord (visible by transparency) is flexed. The fusiform body and snout are considered long. The mouth is in a terminal position with the presence of numerous tiny teeth, with a pair of maxillary barbels (which do not extend beyond the anal fin) and two pairs of mentonian barbels (which do not extend beyond the anus). The nostrils are double, with the part closest to the snout in a tubular shape. The eyes are very small, pigmented, rounded, and far from the snout, and the operculum extends beyond the base of the pectoral fin. The anal opening reaches the middle region of the body (50.34% of SL). Pigmentation consists of irregular punctiform and dendritic chromatophores restricted to the dorsolateral region of the head, the base of the dorsal fin, and the upper part of the digestive system. There are discrete punctiform melanophores in the ventral region of the abdomen, arranged in the anterior and posterior extremities of the stomach.
The embryonic membrane (finfold) begins to be absorbed. The adipose fin has a triangular shape, considered smooth compared to other species of Brachyplatystoma. The caudal, anal, dorsal, and pectoral fins are present, and their rays are in formation. At this stage, it has a definitive number of 46 myomeres (19 preanal and 27 postanal), without variations throughout development.
Posflexion (Figure 2B): the standard length of larvae at this stage ranged from 16.09 to 17.88 mm. The body becomes moderate, and features such as the snout, mouth, nostrils, eyes, operculum, barbels, and pigmentation pattern remain similar to the previous stage. However, the pigmentation is more intense with irregular punctiform chromatophores on the body. The pelvic fin bud is present. Throughout development, the rays begin to develop. The dorsal fin has two spines and six branched rays, and in the pectoral fin, a serrated aculeus is visible, and by the end of this stage, the embryonic membrane is completely absorbed. There is a long filament in the first ray of the caudal fin, and the number of myomeres remains similar to the previous stage.
Juvenile Development
The standard length ranged from 28.03 to 29.29 mm. The mouth becomes subterminal, and there are numerous tiny teeth, the nostrils remain double, one of which is tubular, and the maxillary barbels do not extend beyond the caudal peduncle and do not have pigmentation. The pigments are restricted to the dorsal and lateral regions of the head and under the entire dorsal region of the body. All fins are hyaline. The rays of all fins are fully formed. The number of spines and fin rays are: pectoral (P): I, 9; ventral (V): I, 5; dorsal (D) II, 6; and anal (A): 19 (Figure 2C).
Morphometric Relationships
Initially, the body varies from long to moderate (17.25% to 21.35%), the eyes are small (6.28% to 7.93%), and the head ranges from moderate in larvae (25.53% to 32.49%) to large in juveniles (35.70% to 36.17%). During larval development, most morphometric variables showed an increase in their morphometric proportions, except for head depth, which decreased (60.18% to 48.18%), while in juveniles, most variables decreased their proportions (Table 1).
- Stages of development Brachyplatystoma rousseauxii (Castelnau, 1855)
- Variables (mm)
- Flexion (n=1)
- Postflexion (n=2)
- Juvenile (n=3)
- Min-Max
- X ± SD
- Min-Max
- X ± SD
- Min-Max
- X ± SD
- HL
- 3.24
- NA
- 5.23-5.63
- 5.43 ± 0.28
- 10.14-10.58
- 10.31 ± 0.23
- HW
- 2.4
- NA
- 3.30-3.90
- 3.60 ± 0.42
- 5.84-6.36
- 6.11 ± 0.26
- HD
- 1.84
- NA
- 2.71-3.14
- 2.93 ± 0.30
- 4.54-4.82
- 4.68 ± 0.14
- SnL
- 1.33
- NA
- 2.53-2.72
- 2.62 ± 0.13
- 5.05-5.16
- 5.09 ± 0.06
- ID
- 1.08
- NA
- 1.61-1.80
- 1.70 ± 0.13
- 2.48-2.53
- 2.51 ± 0.02
- DO
- 0.25
- NA
- 0.34-0.36
- 0.35 ± 0.01
- 0.63-0.81
- 0.73 ± 0.08
- SL
- 12.68
- NA
- 16.09-17.88
- 16.99 ± 1.26
- 28.03-29.29
- 28.66 ± 0.63
- Sta
- 6.38
- NA
- 8.57-9.71
- 9.14 ± 0.80
- 15.94-16.26
- 16.15 ± 0.18
- BD
- 2.19
- NA
- 3.35-3.42
- 3.38 ± 0.04
- 5.30-6.11
- 5.69 ± 0.40
- SnP
- 2.89
- NA
- 5.07-5.13
- 5.10 ± 0.04
- 9.29-9.49
- 9.41 ± 0.11
- SnA
- 8.12
- NA
- 10.98-11.92
- 11.45 ± 0.66
- 18.42-19.68
- 19.07 ± 0.62
- SnD
- 4
- NA
- 5.77-6.43
- 6.10 ± 0.46
- 10.64-11.84
- 11.23 ± 0.60
- SnV
- NA
- NA
- 7.40-7.97
- 7.69 ± 0.33
- 13.87-14.97
- 14.33 ± 0.57
- Morphometric proportions (%)
- SnL/HL
- 41.14
- NA
- 44.92-52.13
- 48.52 ± 5.09
- 47.72-50.95
- 49.40 ± 1.62
- DO/HL
- 7.8
- NA
- 6.53-6.59
- 6.56 ± 0.04
- 6.28-7.93
- 7.12 ± 0.82
- HD/HL
- 56.82
- NA
- 48.18-60.18
- 54.18 ± 8.47
- 44.41-46.33
- 45.43 ± 0.96
- ID/HW
- 44.97
- NA
- 46.13-48.73
- 47.43 ± 1.83
- 39.76-43.27
- 56.36 ± 0.73
- Sta/SL
- 50.34
- NA
- 53.29-54.33
- 53.81 ± 0.73
- 55.52-56.87
- 56.36 ± 0.73
- HL/SL
- 25.53
- NA
- 31.48-32.49
- 31.99 ± 0.71
- 35.70-36.17
- 36.00 ± 0.25
- BD/SL
- 17.25
- NA
- 18.75-21.26
- 20.00 ± 1.77
- 18.92-21.35
- 19.85 ± 1.30
- SnP/SL
- 22.8
- NA
- 28.39-31.92
- 30.15 ± 2.49
- 31.71-33.78
- 32.87 ± 1.05
- SnA/SL
- 64.03
- NA
- 66.66-68.27
- 67.46 ± 1.13
- 64.31-68.18
- 66.56 ± 2.00
- SnD/SL
- 31.57
- NA
- 35.90-35.96
- 35.93 ± 0.04
- 37.99-41.34
- 39.18 ± 1.87
- SnV/SL
- NA
- NA
- 44.56-46.02
- 45.29 ± 1.03
- 48.28-52.27
- 50.01 ± 2.04
- Myomeres
- Mode
- Mode
- Mode
- Preanal
- 19
- 19
- 19
- 19
- 19
- 19
- Posanal
- 27
- 27
- 27
- 27
- 27
- 27
- Total
- 46
- 46
- 46
- 46
- 46
- 46
- Rays
- Pectoral
- 6
- 6
- I,6-8
- NA
- I,9
- I,9
- Anal
- 13
- 13
- 13-16
- NA
- 19
- 19
- Dorsal
- I,6
- I,6
- II,6
- NA
- II,6
- II,6
- Pelvic
- FA
- FA
- 5
- NA
- I,5
- I,5
Table 1: Morphometric variables, Minimum values (Min), maximum values (Max), mean (X), standard deviation (SD) and
The absence of individuals in the early stages of development of B. rousseauxii (yolk sac and pre-flexion stage) is because of the reproductive biology of this species. Its breeding occurs in the headwaters of the rivers that form the Solimões/Amazonas system, close to the Andes - spawning areas, at a distance greater than 1500 km from the study area, which probably made it impossible to capture these stages.
Brachyplatystoma larvae and juveniles differ from other species of the Pimelodidae family by the small eyes, barely visible swim bladder, presence of filaments in the caudal fin from the end of the flexion stage vs presence of large eyes, highly visible swim bladder, and absence of caudal filaments, as in Pimelodus [21]. They are distinguished from species of genera Pseudoplatystoma, Sorubim, and Hemisorubim by possessing a triangular adipose fin vs. straight adipose fin [20, 21, 22, 23].
Brachyplatystoma rousseauxii can be easily distinguished from all other six valid species of the genus Brachyplatystoma, mainly because it is the only one to have 46 total myomeres. The vertebrae data (45) that were cleaned in the elaboration of this study corroborate the findings of [24]. In ichthyology, the intra and interspecific variations presented by the numbers of vertebrae are a useful tool for making comparisons and assisting in taxonomic framing [25, 26].
It is noteworthy that from the vertebrae number of adults when considering all the vertebrae, including the Weber apparatus and the urostyle, plus a unit, it is possible to estimate and relate the number of myomeres of the larvae (muscle segments). Thus, providing subsidies and expanding the possibilities of identification of species during the initial stages of development collected in the natural environment [27, 28]. The myomere count (considering all myomeres from the first occipital to the last urostilar) is an easy measure to be performed in the larvae since, unlike the vertebrae count, it does not require special preparation, therefore it can be performed in a stereomicroscope with polarized light.
In addition, the presence of characteristics such as eyes far from the snout; short barbels not extending beyond the anal fin; intestine reaching the middle region of the body; pectoral fin not extending laterally beyond the third ray of the dorsal fin, and the absence of pigmentation in all fins are fundamental for the identification of larvae and juveniles of B. rousseauxii.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
Acknowledgements
The authors thank the Brazilian Ministry of Science, Technology, Innovation and Communication (MCTIC) and the Mamirauá Institute for Sustainable Development (MISD) for the funding from the project. Also to the National Council for Scientific and Technological Development (CNPq) for the scholarship in scientific research granted to the first author. To Laboratory of Ichthyoplankton Ecology and Inland Waters Fishing (LEIPAIUFOPA-http://leipaiufopa.com) for the technical support.
Funding
Brazilian Ministry of Science, Technology, Innovation and Communication (MCTIC) and Mamirauá Institute for Sustainable Development (MISD).
References
-
García-Vásquez AR, Sánchez H, Rodríguez R, Montreuil V, Vargas G, et al. (2009) Hábitos alimenticios del dorado Brachyplatystoma rousseauxii (Castelnau, 1855) en la amazonía peruana. Folia Amazonica 18(1): 7-13.
-
Queiroz LJ, Torrente Vilara G, Ohara WM, Pires THS, Zuanon J, et al. (2013) Peixes do rio Madeira. Volume 3, São Paulo: Dialeto Latin American Documentary pp: 1163.
-
Dagosta FCP, Pinna, MD (2019) The Fishes of the Amazon: Distribution and Biogeographical Patterns, with a Comprehensive List of Species. Bulletin of the American Museum of Natural History 431: 1-163.
-
Cañas CM, Pine WE (2011) Documentation of the temporal and spatial patterns of Pimelodidae catfish spawning and larvae dispersion in the Madre de Dios River (Peru): insights for conservation in the Andean- Amazon headwaters. River Research and Applications 27(5): 602-611.
-
Duponchelle F, Pouilly M, Pécheyran C, Hauser M, Renno JF, et al. (2016) Trans-Amazonian natal homing in giant catfish. Journal of Applied Ecology 53(5): 1511-1520.
-
Barthem R, Goulding M, Leite RG, Cañas C, Forsberg EV, Petry P, et al. (2017) Goliath catfish spawning in the far western Amazon confirmed by the distribution of mature adults, drifting larvae and migrating juveniles. Scientific Reports 7.
-
Córdoba, EA, Joven León ÁV, Bonilla Castillo CA, Petrere MJ, Peláez M, et al. (2013) Breeding, growth and exploitation of Brachyplatystoma rousseauxii in the Caqueta River, Colombia. Neotropical Ichthyology 11: 637-647.
-
Cella-Ribeiro A, Assakawa LF, Torrente Vilara G, Zuanon J, Leite RG, et al. (2015) Temporal and spatial distribution of young Brachyplatystoma spp. (Siluriformes: Pimelodidae) along the rapids stretch of the Madeira River (Brazil) before the construction of two hydroelectric dams. Journal of Fish Biology 86: 1429- 1437.
-
Lima JL, Sousa KNS, Braga TMP (2016) Representação espacial da pesca de Brachyplatystoma rousseauxii (Siluriformes - Pimelodidae) em paisagens fluviais do complexo fluvio-lacustre do Ituqui, Baixo Amazonas, Brasil. Biota Amazônia 6(4): 74-80.
-
Cruz REA, Isaac VJ, Paes ET (2017) A pesca da dourada Brachyplatystoma rousseauxii (Castelnau, 1855) na região do Baixo Amazonas, Brasil. Boletim do Instituto de Pesca 43(4): 474-486.
-
Agostinho AA, Gomes LC, Santos NCL, Ortega JCG, Pelicice FM (2016) Fish assemblages in Neotropical reservoirs: Colonization patterns, impacts and management. Fisheries Research 173(1): 26-36.
-
Loures RC, Pompeu PS (2018) Long-term study of reservoir cascade in south-eastern Brazil reveals spatio- temporal gradient in fish assemblages. Marine and Freshwater Research 69(12): 1983-1994.
-
Zacardi DM, Suzuki MAL, Oliveira CC, Oliveira LS, Cajado RA (2021) O gradiente longitudinal (rio-barragem) do reservatório de Curuá-Una afeta a composição das assembléias de larvas de peixes?. Revista Brasileira de Meio Ambiente 9(1): 099-112.
-
Sousa RGC, Freitas HCP, Zacardi DM, Faria-Junior CH (2022) Effects of river dams on the fish guilds in the northwest region of the Brazilian Amazon. Fisheries Research 243: 106091.
-
Cajado, RA, Oliveira LS, Silva FKS, Zacardi DM (2021) Early development of the Neotropical fish known as long sardine Triportheus auritus (Valenciennes 1850) (Characiformes, Triportheidae). Journal of Applied Ichthyology 37: 759-769.
-
Oliveira LS, Cajado RA, Silva FKS, Andrade MC, Zacardi DM (2022) Early development of two commercially valuable fish from the lower Amazon River, Brazil (Characiformes: Serrasalmidae). Neotropical Ichthyology 20(1): e210024.
-
Silva FKS, Cajado RA, Oliveira LS, Santos Z, Santos JÁ, et al. (2022) Early development of Prochilodus nigricans Spix & Agassiz 1829 (Characiformes: Prochilodontidae) in captivity. Aquaculture Research pp: 1-16.
-
CONCEA (2013) Diretrizes da prática de eutanásia do Conselho Nacional de Experimentação Animal. Ministério da Ciência Tecnologia e Inovação, Brasília, DF pp: 1-49.
-
Ahlstrom EH, Butler JL, Sumida BY (1976) Pelagic stromateoid fishes (Pisces, Perciformes) of the eastern Pacific: Kinds, distributions, and early life histories and observations on five of these from the Northwest Atlantic. Bulletin of Marine Science 26(3): 285-402.
-
Nakatani K, Agostinho AA, Baumgather G, Bialetzki A, Sanches PV, et al. (2001) Ovos e larvas de peixes de água doce: desenvolvimento e manual de identificação. EDUEM.
-
Silva FKS, Cajado RA, Oliveira LS, Ribeiro FRV, Zacardi DM (2021) Early ontogeny of Pimelodus blochii Valenciennes, 1840 (Siluriformes: Pimelodidae): Neotropical catfish. Zootaxa 4948(1): 83-98.
-
Andrade FF, Makrakis MC, Lima AF de Assumpção, L de Makrakis S, Pini SFR (2014) Desenvolvimento embrionário, larval e juvenil de Hemisorubim platyrhynchos (Siluriformes, Pimelodidae) da bacia do rio Paraná. Iheringia série Zoologia 104: 70-80.
-
Andrade FF, Lima AF, Assumpção L, Makrakis S, Kasai RID, et al. (2016) Characterization of the early development of Pseudoplatystoma reticulatum Eigenmann & Eigenmann, 1889 (Siluriformes: Pimelodidae) from the Paraguay River Basin. Neotropical Ichthyology 14(2): e150032.
-
Lundberg JG, Akam A (2005) Brachyplatystoma capapretum: a new species of goliath catfish from the Amazon basin, with a reclassification of allied catfishes (Siluriformes: Pimelodidae). Copeia 2005(3): 492-516.
-
Frank KT (1991) Predictiment variation from year class specific vertebral counts: an analysis of the potencial and a plan for verification. Can J Fish Aquat Sci 48: 1350- 1357.
-
Kozhara AV, Izyumov Yu G, Kas’Yanov NA (1996) General and geographic variation of the vertebral numbers in some freshwater fish. J Ichthyol 36(3): 223-237.
-
Araújo Lima CARM, Donald E (1998) Número de vértebras de Characiformes do rio Amazonas e seu uso na identificação de larvas do grupo. Acta Amazônica 18(1-2): 351-358.
-
Borges MEE, Nakatani K, Bialetzki A, Felis MEG (2000) Diferenciação do número de vértebras de algumas espécies de Characiformes (Osteichthyes) da bacia do Rio Parana, Brasil. Acta Scientiarum Biol Sci 22: 539.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework