Taxonomy of the Superfamilies Congiopodoidea Gill, 1889 and Synanceiidea Swainson, 1839 (Pisces: Perciformes)
The genus Congiopodus includes 6 species, it is recommended that the species Congiopodus kieneri be considered valid. The genus belongs to the monotypic family Congiopodidae. Its sister family Zanclorhynchidae includes the monotypic genus Alertichthys and Zanclorhynchus with two species. Both families, in turn, form the superfamily Congiopodoidea. Its sister superfamily Synanceiidea is recommended to be considered as follows: Synanceiidae, Tetrarogidae, Choridactylidae, Minoidae, Aploactinidae, Gnathanacanthidae and Pataecidae. Both superfamilies belong to the suborder Scorpaenoidei of the order Perciformes. The position of the family Apistidae needs further clarification. Also it is needed more studies to clarify the position of genus Erosa in the family Synanceiidae. The parameters used in the previous article are given, the resulting cladogram is described.
Introduction
The position and composition of the superfamily Congiopodoidea have been revised many times and continue to be debated to this day [1]. Until recently, the group was considered as a family. The fishes of the discussed group live in the Southern Hemisphere, inhabit the shelves of South America, Africa, Australia and New Zealand in tropical to temperate waters (genus Congiopodus Perry, 1811), Alertichthys blacki Moreland, 1960 is endemic to the New Zealand Plateau, the genus Zanclorhynchus Gunther, 1880 inhabits the shelves of islands and uplifts within the Circum- Antarctic Current of the Southern Ocean [2, 3, 4].
The superfamily Congiopodoidea comprises the monotypic family Congiopodidae Gill, 1889 and Zanclorhynchidae Andriashev, 1993 with two genera, Zanclorhynchus and Alertichthys Moreland, 1960. The genus Congiopodus comprises 6 species: C. torvus (Gronow, 1772), C. peruvianus (Cuvier, 1829), C. spinifer (Smith, 1839), C.
leucopaecilus (Richardson, 1846), C. kieneri (Sauvage, 1878), and C. coriaceus Paulin et Moreland, 1979 [4, 5, 6]. Congiopodus kieneri is now often reduced to a synonym, although in 1971 Hureau [7] unconditionally confirmed the validity of the species. But this work was published in French and was ignored. Thus, in the well-known Eschmeyr’s Catalog of Fishes, C. kieneri is synonymized with C. peruvianus without reference to supporting works [4], although, on the contrary, in this catalog are works where C. kieneri is indicated as valid [5, 8]. Moreover, for example, Moland JM [9] argued that only the C. peruvianus inhabits the waters off South America, but almost 20 years later, in collaboration with Paulin [5], he changed his mind and mentioned C. kieneri as valid species. A clear diagnosis of C. kieneri was also made by Mandritsa SA [10]. In this paper, the author follows Hureau and accepts the validity of C. kieneri.
The family Zanclorhyncidae includes the monotypic genus Alertichthys Moreland, 1960 and Zanclorhynchus Gunther, 1880 with two species Z. spinifer Günther, 1880
and Z. chereshnevi Balyshkin, et al. Zhukov M [11]. In turn, Z. spinifer includes 4 subspecies: Z. s. spinifer Günther, 1880, Z. s. heracleus Zhukov et Balushkin, 2018, Z. s. armatus Zhukov, 2019 and Z. s. macquariensis Zhukov M [3, 12].
Not so long ago, the poorly studied genus Perryena Whitley, 1940 was traditionally included in the family Congiopodidae, usually as incertae sedis [13, 14, 15, 16]. Later, it was assigned to Neocentropogoninae Mandrytsa, 2001 in the family Tetrarogidae Smith, 1949 [10, 15]. Now the genus Perryena with a single species Perryena leucometopon (Waite, 1922) is separated into a family Perryenidae Honma, Imamura et Kawai, 2013, a sister clade to Tetrarogidae + Synanceiidae Swainson, 1839 + Aploactinidae Jordan et Starks, 1904 + Сongiopodidae + Gnathanacanthidae Gill, 1892 + Pataecidae Gill, 1872, where the basal group is family Apistidae Gill, 1859 [17]. Thus, Perryena is not closely related to Congiopodidae.
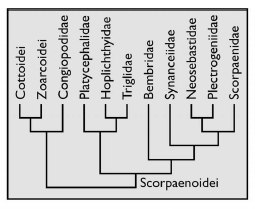
Initially, the genus Zanclorhynchus was assigned to the family Scorpaenidae Risso, 1827 [18, 19], but since 1937 it has been assigned to the family Congiopodidae [20]. The monophyly of this group has been shown by a number of authors [10, 16, 21] and is now beyond doubt. Japanese researchers, who devoted many years to the study of scorpionfishes, attributed Congiopodidae s.l. to superfamily Scorpaenoidea sensu Imamura, 2004 with Sebastidae Kaup, 1873, Sebastolobidae Imamura, 2004, Scorpaenidae, Apistidae, Tetrarogidae, Synanceiidae, Aploactinidae, Gnathanacanthidae, Pataecidae, Caracanthidae Gill, 1885, Eschmeyeridae Mandrytsa, 2001, Neosebastidae Matsubara, 1943, Plectrogeniidae Fowler, 1938, Parabembridae Jordan et Hubbs, 1925, Bembridae Kaup, 1873, Triglidae Rafinesque, 1815, Peristediidae Jordan et Gilbert, 1883, Hoplichthyidae Kaup, 1873, and Platycephalidae Swainson, 1839 [22]. They considered the position of this superfamily in the suborder Parameters Used for Cladistis Analysis Scorpaenoidei of the order Scorpaeniformes, but now the rank of the order has been reduced to the rank of a suborder in the order Perciformes, and the rank of Congiopodidae s.l. raised to a superfamily, which requires a revision of the composition of Scorpaenoidea sensu Imamura, 2004, but does not cancel the relationship.
Genetic studies show a different picture from the morphological described above. The studies of Smith, et al. [23] deny the monophyly of the group Congiopodidae + Zanclorhynchidae, for example. Other works confirm their monophyly, but Congiopodoidea fishes are placed on the phylogenetic tree at a considerable distance from Scorpaenidae, closer to the root of all scorpion fish [24, 25]. This position, in turn, is consistent with other morphological and taxonomic studies, according to the results of which it is proposed to raise the rank of Congiopodoidea to the level of a suborder [10, 26, 27, 28]. The purpose of this work is to clarify the above constructions of relationships. It also became necessary to describe all parameters used for cladistic analysis in previous paper as well as to present full character matrix and to describe the resulting cladogram [1].
Materials and Methods
Phylogenetic analysis was carried out using the ASADO 1.61 and PAUP 4.0a169 software packages, the Heuristic algorithm was used; out of 77 characters, 9 are uninformative. The tree is rooted, the outgroups are Sebastes, Scorpaena and Scorpaenodes. All parameters were given equal weight, the values of the parameters were ordered, except Nos. 10, 19, 45. The value of all parameters follows Voskoboinikova, et al. [1] except character 24, mostly follows Imamura H [21] but their numbering may differ.
| 1–5 | 6–10 | 11–15 | 16–20 | 21–25 | 26–30 | 31–35 | 36–40 | 41–45 | 46–50 | 51–55 | 56–60 | 61–65 | 66–70 | 71–75 | 76–77 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sebastes | 00000 | 00100 | 00000 | 00000 | 00020 | 00000 | 00000 | 00000 | 00000 | 01000 | 00000 | 00000 | 00000 | 00100 | 00000 | 00 |
| Scorpaena | 00000 | 01210 | 00000 | 00000 | 00020 | 00000 | 10000 | 00000 | 00010 | 01000 | 00000 | 00000 | 00100 | 00100 | 00000 | 00 |
| Scorpaenoides | 00000 | 00210 | 00000 | 00000 | 00020 | 00001 | 00000 | 00000 | 00010 | 00000 | 00000 | 00000 | 00100 | 00000 | 00000 | 00 |
| Apistus | 00000 | 01211 | 00100 | 00020 | 00020 | 01000 | 01000 | 00000 | 00010 | 01010 | 00000 | 00000 | 00100 | 00101 | 00000 | 00 |
| Tetraroge | 00230 | 01211 | 00000 | 00000 | 00100 | 00000 | 01100 | 00001 | 01011 | 01000 | 00100 | 00000 | 21131 | 01111 | ?0000 | 00 |
| Synanceia | 00230 | 01212 | 00000 | 00020 | 00021 | 00001 | 01100 | 00010 | 01112 | 01000 | 00100 | 00000 | 00131 | 01111 | 00000 | 10 |
| Erosa | 00230 | 01211 | 00000 | 00000 | 10021 | 00001 | 01100 | 00001 | 0111? | 01000 | 00010 | 00000 | 00101 | 01111 | 10000 | 10 |
| Inimicus | 00230 | 01211 | 00000 | 00000 | 00021 | 00001 | 01100 | 00101 | 01012 | 01020 | 00100 | 00010 | 20131 | 01101 | 10101 | 10 |
| Choridactylus | 00230 | 01211 | 00000 | 20000 | 00021 | 00001 | 01100 | 10101 | 01110 | 21030 | 00100 | 00010 | 20131 | 01101 | 10000 | 00 |
| Minous | 00230 | 01211 | 00100 | 10020 | 10021 | 00001 | 01100 | 00101 | 01112 | 11011 | 00001 | 10011 | 20131 | 01101 | 10100 | 01 |
| Aploactis | 00230 | 01212 | 00000 | 00021 | 00021 | 00001 | 01000 | 01101 | 0?212 | 01001 | 00121 | 01011 | 00131 | 10101 | 10000 | 01 |
| Erisphex | 00230 | 01211 | 00000 | 00001 | 00121 | 00001 | 01000 | 01101 | 01112 | 01001 | 01121 | 01011 | 21121 | 10111 | 10000 | 01 |
| Congiopodus | 01121 | 01220 | 00111 | 21001 | 11101 | 00111 | 01010 | 12001 | 00012 | 10101 | 01100 | 11010 | 00101 | 01111 | 11101 | 10 |
| Alertichthys | 00110 | 11221 | 21111 | 21101 | 01121 | 10011 | 01011 | 01101 | 10012 | 11001 | 11201 | 11001 | 00101 | 10111 | 11101 | 11 |
| Zanclorhynchus | 00110 | 11221 | 11111 | 21001 | 01111 | 10011 | 01011 | 01101 | 10012 | 11001 | 11201 | 11111 | 10101 | 10111 | 11100 | 11 |
| Gnathanacanthus | 10230 | 01211 | 00000 | 20010 | 00001 | 00001 | 0101? | 10001 | 01111 | 01001 | 00001 | 00001 | 20111 | 00101 | 11011 | 01 |
| Pataecus | 10230 | 00012 | 00000 | 00121 | 00001 | 00001 | 01010 | 11001 | 01111 | 01101 | ?023? | 01201 | 20131 | 01111 | 11111 | 21 |
Table 1: Character matrix of superfamilies Synanceiidea and Congiopodoidea.
- The first three infraorbitals: firmly connected (0), separated (1). The first three infraorbitalia are fused in all the studied groups (0), except for Gnathanacanthus and Pataecus, in which the value is 1
- Vertical ridge on parietale: absent (0), present (1). Only Congiopodus (
53. The supratemporal commissure (CST) of sensory system: continious (0), inerrupted (1), interrupted three times (2), ordered as 0–1–2. CST is continuous in Sebastes, Scorpaena, Scorpaenodes, Apistus, Erosa, Minous, and Gnathanacanthus (0); interrupted once in Tetraroge, Synanceia, Inimicus, Choridactylus, Aploactis, Erisphex, Congiopodus (1); the commissure interrupted three times in Pataecus, Alertichthys, and Zanclorhynchus (2). 54. Number of rays of the ventral fin: six (0), five (1), four (2), absent (3), ordered as 0–1–2–3. Five rays of the ventral fin are noted in Erosa (1); four in Aploactis and Erisphex (2); no rays in Pataecus (3); the rest have a value of 0. 55. Branched rays of the ventral fin: present (0), absent (1). The branched rays are absent in Minous, Aploactis, Erisphex, Alertichthys, Zanclorhynchus, and Gnathanacanthus (1); due to the absence of rays in Pataecus, the value of this character is not assigned; the rest have a value of 0. 56. Number of spines on the first pterygiophore of the dorsal fin: two (0), one (1). One spine is noted in Minous, Congiopodus, Zanclorhynchus, and Alertichthys (1), the rest have a value of 0. 57. Spine on the first pterygiophore of the anal fin: present (0), absent (1). Character 57-1 in Aploactis, Erisphex, Congiopodus, Zanclorhynchus, Alertichthys, and Pataecus, the rest have a value of 0. 58. Second ray on first pterygiophore of anal fin: present (0), sometimes present (1); absent (2), ordered as 0–1– 2. The second element on the first pterygiophore as an intraspecific variability is often absent in Zanclorhynchus (1), absent in Pataecus (2); the rest have a value of 0. 59. Ray on the second pterygiophore of the anal fin: spine (0), soft (1). A soft ray was noted in Inimicus, Choridactylus, Minous, Aploactis, Congiopodus, and Zanclorhynchus (1), the rest have a value of 0. 60. Branched rays of the dorsal and anal fins: present (0), absent (1). The rays in the anal and dorsal fins are not branched in Minous, Aploactis, Erisphex, Alertichthys, and Zanclorhynchus (1), the rest have a value of 0. 61. Number of rays on the last pterygiophores of the dorsal and anal fins: two (0), usually two (1); one (2), ordered as 0–1–2. The ray on the last pterygiophore of the dorsal and/or anal fins is sometimes single as an intraspecific variability in Zanclorhynchus (1); always one ray in Tetraroge, Inimicus, Choridactylus, Minous, Gnathanacanthus, and Pataecus (2); the rest have a value of 0. 62. First pterygiophore of dorsal fin: free (0), fused with neurocranium (1). It is fused with the neurocranium in Tetraroge and Erisphex (1), the rest have a value of 0. 63. Supraneural: present (0); absent (1). One supraneural was noted in Sebastes (0); the rest have a value of 1. 64. Stays of the dorsal and anal fins: free and ossified (0), fused with the previous pterygiophore (1), free cartilage (2), absent (3), ordered as 0–1–2–3. The ossified stay is separated from the previous pterygiophore in Sebastes, Scorpaena, Scorpaenodes, Apistus, Erosa, Congiopodus, Zanclorhynchus, and Alertichthys (0); ossified and firmly attached to the previous pterygiophore in Gnathanacanthus (1); unossified in Erisphex (2); absent in Tetraroge, Synanceia, Inimicus, Choridactylus, Minous, Aploactis, and Pataecus (3). 65. Tubular scales of the lateral line: absent (0), present (1). Lateral line with perforated scales found in Sebastes, Scorpaena, Scorpaenodes, and Apistus (0); in other genera, the lateral line is represented by tubular elements (1). 66. Spines on the scales of the lateral line: absent (0), present (1). Spines are noted in Aploactis, Erisphex, Zanclorhynchus, and Alertichthys (1); the rest have a value of 0. 67. Scales on the body: present (0), absent (1). Body scales absent in Tetraroge, Synanceia, Erosa, Inimicus, Choridactylus, Minous, Congiopodus, and Pataecus (1); the rest have a value of 0. 68. Third and fourth hypurals: separate (0); fused (1). Separate hypurals are noted for Scorpaenodes (0), the rest have a value of 1. 69. Fifth hypurale: present (0), absent (1). The fifth hypurale is absent in Tetraroge, Synanceia, Erosa, Erisphex, Congiopodus, Zanclorhynchus, Alertichthys, and Pataecus (1); for the rest, the attribute value is 0. 70. Lower hypural plate and parhypurale: separate (0), fused (1). Parhypurale separated from lower hypural plate in Sebastes, Scorpaena, and Scorpaenodes (0), the rest have a value of 1. 71. Hemal spine and centrum of the third preural vertebra: separated (0), fused (1). The character 71-0 found in Sebastes, Scorpaena, Scorpaenodes, Apistus, and Synanceia, no value defined for Tetraroge, the rest have a value of 1. 72. Hemal spine and centrum of the second preural vertebra: separated (0), fused (1). The character 72-1 noted for Congiopodus, Zanclorhynchus, Alertichthys, Gnathanacanthus, and Pataecus (1); the rest have a value of 0. 73. Urostyle and upper hypural plate: separated (0), fused (1). The upper hypural plate is fused with the urostyle in Inimicus, Minous, Congiopodus, Alertichthys, Zanclorhynchus, and Pataecus (1), the rest have a value of 0. 74. Urostyle and lower hypural plate: separated (0), fused (1). The lower hypural plate is fused with the urostyle in Gnathanacanthus and Pataecus (1), the rest have a value of 0. 75. Uroneural: present (0), absent (1). Absent in Inimicus, Congiopodus, Alertichthys, Gnathanacanthus, and Pataecus (1); the rest have a value of 0. 76. Number of epurals: three (0), two (1), one (2), ordered as 0–1–2. Two epurals were noted in Synanceia, Erosa, Inimicus, Congiopodus, Zanclorhynchus, and Alertichthys (1); one epurale in Pataecus (2); the rest have a value of 0. 77. Branched rays of the caudal fin: present (0), absent (1). In the caudal fin, the rays do not branch in Minous, Aploactis, Erisphex, Zanclorhynchus, Alertichthys, Gnathanacanthus, and Pataecus (1), the rest have a value of 0.
Discussion
Morphology
Imamura H [21] used 17 synapomorphies to substantiate the monophyly of Congiopodoidea, Ishii, et al. [16] used only 11 of them because of the impossibility of including 6 reversions in a particular method of building phylogenetic relationships, but added 2 more synapomorphies. In total, 35 characters are apomorphic according to Ishii, et al. [16] for Congiopodidae sensu Honma, Imamura, and Kawai, 2013:
- Absence of fifth os infraorbitale.
- The presence of tubercles on the skull, bone granules are also found on the head of Apistidae, Minous Cuvier, 1829, Triglidae, Peristediidae, Hoplichthyidae and some Platycephalidae (e.g. Suggrundus Whitley, 1930 and Rogadius Jordan et Richardson, 1908).
- Ossa nasalia firmly attached to the cranium.
- Os intercalare and os posttemporale sutured.
- Praeoperculo-mandibular and temporal canals of the head are not connected.
- Baudelot’s ligament is absent.
- Os palatinum is separated from ectopterigoideum (an autapomorphy of Congiopodidae s.l. among the superfamily Scorpaenoidea sensu Imamura 2004).
- Absence of the first vertebra.
- One ray on the first pterigiophore.
- Absence of levator posterior.
- Presence of adductors I-III.
Ishida M [29], as well as Imamura, et al. [22], include the presence of separate adductors I-III into autapomorphic characters of Congiopodidae s.l. in the superfamily Scorpaenoidea sensu Imamura
14. No teeth on the vomer. 15. Urostyle fused with the upper hypural plate. 16. Suborbital stay broad and firmly connected to os praeoperculum. 17. The fourth os infraorbitale absent. 18. Os prooticum and os intercalare separated. 19. Teeth on os palatinum are absent. 20. The metapterigoid lamina absent. 21. Branchiostegal rays 5-6. 22. Os praeoperculum with smooth edge. 23. No tooth plate on third epibranchiale. 24. Os extrascapulare mediale absent. 25. Os extrascapulare laterale with one element (elongated tubular) with two openings of sensory pores. 26. No first spine on first anal pterygiophore. 27. Supraneurale absent. 28. The first and second hypuralia are fused (in the lower hypural plate). 29. The third and fourth hypuralia are fused (in the upper hypural plate).
30. Fifth hypurale absent. 31. Fused lower hypural plate and parhypurale. 32. Attachment of a A2A3 musculus adductor mandibulae partially medially in relation to m. levator arcus palatini. 33. Musculus levator opercula attaches to os pteroticum and os posttemporale in Zanclorhynchus and Alertichthys (an even more advanced position in Congiopodus in the family - attachment of m. levator opercula only to os posttemporale). 34. Musculus transversus ventralis anterior and m. ventralis posterior separated. 35. Tubular lateral line segments (tubular lateral line scales are also characteristic of representatives of the following families from the superfamily Scorpaenoidea sensu Imamura, 2004: Tetrarogidae, Synanceiidae, Aploactinidae, Gnathanacanthidae, and Pataecidae) [21].
In total, the clade Congiopodidae sensu Honma, Imamura, Kawai, 2013 in the work of Ishii, et al. [16] is supported by 9 characters (Figure 1).
![Figure 1: Phylogenetic relationships of the family Congiopodidae sensu Honma, Imamura, et al. [16] (Figures 1&2).](/fulltextimages/9202/fig_1.png)
In this paper, the author does not agree with the absence of Baudelot’s ligament and the loss of the first vertebra. As evidence of such a loss, Ishii, et al. [16] presented the following characteristics, in their opinion, of Congiopodoidea sensu Voskoboinikova et Zhukov, 2021:
- The absence of Baudelot’s ligament.
- First neural spine bent anteriorly, although in all families belonging to the same clade with Congiopodidae and Zanclorhynchidae: Apistidae, Tetrarogidae, Synaiceiidae, Aploactinidae, Gnathanacanthidae, and Pataecidae (Imamura, 2004), the first two neural spines are bent anteriorly.
- In Zanclorhynchus and Alertichthys, the first two pterygiophores inserted between the first and second neural spines; in Congiopodidae, the pterygiophores are shifted onto the cranium – while in other families the first 2 pterygiophores inserted between the second and third neural processes [16].
Thus, for example, the second neural spine is homologous to the third one in all other fish of the superfamily Scorpaenoidea sensu Imamura, 2004.
The erroneousness of that statements was recently proven in a study on comparative osteology of the neurocraniums Alertichthys, Zanclorhynchus, and Congiopodus [1]. Baudelot’s ligament was found. In Congiopodus, the pterygiophores and neural spines are not shifted onto the cranium. The pterygiophores are inclined strongly anteriorly in the long and wide cavity formed for them from the upper ridges of the ossa parietalia, and with their lower ends they are inserted between the neural spines of the first and second vertebrae already behind the posterior profile of the skull.
Pterygiophores shifted onto the skull, for example, are found in Gnathanacanthidae and Pataecidae from the neighboring clade, but not in Congiopodidae. Congiopodidae have the first vertebra; in Congiopodus, it is fused with the cranium and is actually included in the posterior wall of the skull; its neural spine separates the medial ends of the ossa epiotici. In Zanclorhynchus and Alertichthys, the first vertebra is not completely fused to the cranium and shows some mobility.
In addition to clarifying the position of pterygiophores and the first vertebra, Voskoboinikova and Zhukov identified 13 additional synapomorphies. In total, they used 77 parameters for analysis, of which 16 were corrected and added regarding to the work of Imamura [21]:
- The presence of a vertical ridge on parietale.
- Position of the first pterygiophores of the dorsal fin.
- Number of first pterygiophores between the first and second neural spines.
- The presence of Baudelot’s ligament.
- The presence of connection of os supraoccipitale and os frontale.
6. Position of the os supraoccipitale. 7. Medial connection of ossa epiotici. 8. The presence of the first vertebra. 9. Os posttemporal state. 10. The presence of spines on the frontale, pteroticum and posttemporale. 11. Presence of posteroventral and ventromedial ridges on os cleithrum. 12. Condition of the precoracoid foramen. 13. The presence of a notch between the anterior processes of the pelvic bones. 14. State of the infraorbital cannal in infraorbital 2. 15. Connection between the praeoperculo-mandibular and temporal canals. 16. State of the supratemporal commissure.
In the group analyzed Voskoboinikova OS [1] the position of Congiopodus + Zanclorhynchus + Alertichthys changed, the group, relative to other genera, took a position closer to the root of the tree, and the genus Apistus also turned out to be basal (Figures 2&3).
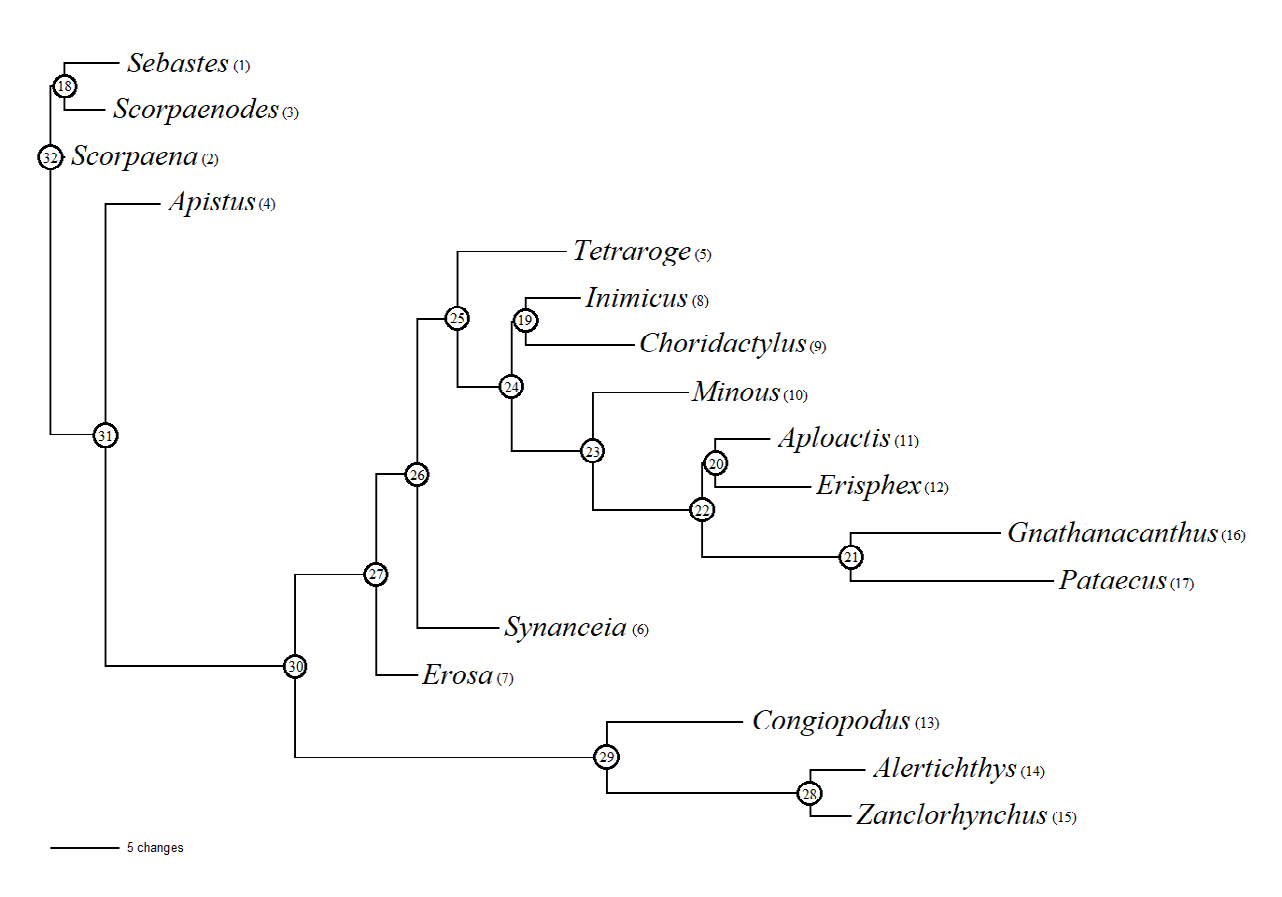
Length of resulting tree 202, CI 0.505, RI 0.664. The Sebastes+Scorpaenodes branch is supported by character 7 (10). Node 18 is supported by characters 8 (21), 9 (10), 44 (10), 63 (10) to Sebastes and 30 (01), 47 (10), 68 (10) to Scorpaenodes. The Scorpaena branch is supported by character 31 (01). In recent molecular genetic studies [24], all the genera studied in the cladistic analysis were assigned to the same family Synanceiidae, with the exception of outgroups (Sebastes, Scorpaena, Scorpaenodes) and genera of the superfamily Congiopodoidea (Congiopodus, Alertichthys, Zanclorhynchus). The resulting cladogram (Figure 2) is consistent with this hypothesis, except for the position of the genus Apistus. It took an basal position relative to other genera under study, its branch is supported by three characters: 19 (02), 27 (01), 49 (01). The branch containing all other analyzed genera is supported by twelve features: 3 (01), 4 (02), 25 (01), 30 (01), 40 (01), 45 (02), 53 (01), 65 (01), 67 (01), 69 (01), 71 (01), 76 (01). The branch Synanceiidae sensu Smith et al., 2018 without Apistus (3027) is supported by six characters: 3 (12), 4 (23), 13 (10), 33 (01), 42 (01), 43 (01). The clade Congiopodoidea (3029) is supported by 22 characters: 9 (12), 14 (01), 15 (01), 16 (02), 17 (01), 20 (01), 22 (01), 23 (01), 24 (21), 25 (12), 29 (01), 34 (01), 37 (01), 46 (01), 50 (01), 52 (01), 56 (01), 57 (01), 59 (01), 72 (01) , 73 (01), 75 (01). The branch of the family Congiopodidae (2913) is supported by ten characters: 2 (01), 5 (01), 10 (10), 21 (01), 24 (10), 28 (01), 36 (01), 37 (12), 47 (10), 48 (01). The branch of the family Zanclorhynchidae (2928) is supported by 15 characters: 4 (21), 6 (01), 11 (01), 12 (01), 26 (01), 35 (01), 38 (01), 41 (01), 51 (01), 53 (12), 55 (0), 60 (01), 66 (01), 67 (10), 77 (01). Alertichthys is supported by four parameters 11 (12), 18 (01), 24 (12), 59 (10). Zanclorhynchus is supported by three - 58 (01), 61 (01), 75 (10).
Erosa is supported by characters 21 (01), 53 (10), 54 (01). In the other direction, the node is supported by one feature - 64 (03). Synanceia is supported by characters 10 (12), 19 (02), 39 (01), 40 (10), 71 (01). The branch 2625 is supported by two parameters 61 (02) and 76 (10). Tetraroge is supported by seven attributes: 23 (01), 24 (20), 25 (01), 30 (01), 43 (01), 45 (21),
62 (01). The branch 2524 is supported by characters 38 (01), 49 (01), 59 (01), 69 (10). The Inimicus + Choridactylus branch is supported by one parameter 49 (12). Within it, Inimicus is supported by characters 43 (10), 73 (01), 75 (01), 76 (01), and Choridactylus by 16 (02), 36 (01), 45 (20), 46 (02), 49 (23). Minous is supported by seven attributes: 13 (01), 16 (01), 21 (01), 46 (01), 53 (10), 56 (01), 73 (01). The 2322 branch is also supported by seven parameters: 20 (01), 33 (10), 37 (01), 49 (01), 54 (02), 57 (01), 67 (01). The clade Aploactis + Erisphex is supported by one character 66 (01). Aploactis is supported by parameters 10 (12), 43 (12), 61 (20), while Erisphex is supported by characters 19 (20), 23 (01), 52 (01), 62 (01), 64 (32), 69 (01). The clade Gnathanacanthus + Pataecus is supported by ten characters: 1 (01), 24 (20), 34 (01), 36 (01), 38 (10), 45 (21), 59 (10), 72 (01), 74 (01), 75 (01). Gnathanacanthus supported by characters 16 (02), 19 (21), 20 (10), 37 (10), 53 (10), 54 (20), 57 (10), 64 (31); Pataecus – 7 (10), 8 (20), 10 (12), 18 (12), 48 (01), 53 (12), 54 (23) , 58 (02), 67 (01), 69 (01), 73 (01), 76 (02).
![Figure 3: Phylogenetic relationships of the superfamily Scorpaenoidea sensu Imamura H [21]. The fish of the group under discussion are shown by an arrow.](/fulltextimages/9202/fig_3.png)
Contradictory information has been published about the presence of teeth on the vomer and os palatinum. The teeth of Alertichthys are described by Moreland JM [9] as small, forming wide oval groups on the symphyses on both jaws. He indicated the presence of teeth on the vomer, visible only on the stained material in large (i.e., about 80 mm) specimens in his collection, and all of his specimens are juvenile. Imamura argued that the genus Congiopodus lacks teeth in the jaws [21]. However, already in 2008, together with Ishii, et al. [16], he suggested the presence of intraspecific variability in the species C. leucopaecilus (teeth are absent / present on both jaws) and C. coriaceus (teeth are absent / present on the lower jaw). Or they suggested the probability that teeth are always present, but can easily fall out [16]. In the original description of the genus Zanclorhynchus, Gunther A [18] indicates the absence of teeth at all: “Snout pointed; mouth very protractile, lateral, narrow, and toothless”. However, Waite ER [19], in describing the second case of capturing Zanclorhynchus from Macquarie Island, describes the presence of very small teeth on the jaws and vomer. The author agrees with the results of a number of studies [3, 6, 11] and acknowledges the presence of teeth on both jaws in the Congiopodoidea group, but their absence on the vomer. Thus, out of 35 apomorphic characters proposed for Congiopodoidea by Ishii, et al. [16], in this work the author recognizes 32 apomorphies.
Agassiz was the first to point out a significant scale weight in the taxonomy of fish, taxonomically dividing fish on this basis: ganoid, placoid, ctenoid, and cycloid fish [30]. This taxonomic division did not last long, but the scale nomenclature introduced by Agassiz still exists. Roberts, in his work on the comparative morphology of Teleostei scales within the group, described the presence of cycloid and three main types of spined scales: spinoid, crenate, and ctenoid [31]. The spinoid type, in turn, was divided into five, the fourth of which is specific, in addition to Congiopodoidea, also for Luvaridae Gill, 1885, Chiasmodontidae Jordan et Gilbert, 1883, juvenile specimens of some Istiophoridae Rafinesque, 1815 and Xiphiidae Rafinesque, 1815, Aploactinidae, some Cottidae Bonaparte , 1831, most Triacanthodidae Gill, 1862 and Monacanthidae Nardo, 1843, and some Molidae Bonaparte, 1835 [31]. This fourth type of spinoid scale is characterized by “scales lack division into fields and are not closely imbricated; they have one or more stout, often buttressed and recurved, spines which arise submarginally, generally from the central region; shape of scale base is often circular” [31]. In the original description, Günther did not consider the submerged scales of Zanclorhynchus to be scales at all, but noted small tubercles on the body [18]. However, there are scales, the scaling is very dense in some parts [3, 11, 12], long spines protrude on the surface of the skin of Zanclorhynchus and Alertichthys [3, 9, 32]. Thus, out of the entire related clade Synanceiidae sensu Smith WL [23], which includes 169 species, the value of this parameter unites Zanclorhynchidae only with Aploactinidae (17 genera, 50 species) and one species of Congiopodus – C. spinifer.
The caudal fin skeletal characters has long been widely used for the bony fish system [33, 34, 35]. According to Ishii, et al. [16], the upper hypural plate in is formed by fusion the third and fourth hypurals, uroneurals and urostyle (H3+H4+ur+ct), H5 is absent; the lower hypural plate was formed from the first and second hypuralia and parhypuralia (H1+H2+PH) [21]. Imamura argues that uroneuralia is absent in all Congiopodidae, which brings them closer to Pataecidae and Peristediidae; in 2008, in collaboration with Ishii N [16], he came to the conclusion that uroneuralies are fused with the upper hypural plate in all Congiopodus, except for C. leucopaecilus, in which the uroneuralia are separate. The latest data obtained on Zanclorhynchus suggest that one pair of uroneurales separated from the upper hypural plate exist at least in the form of intraspecific variability in the genus Zanclorhynchus [3]. The lower hypural plate fused with the parhypuralia considers a synapomorphy belonging to the clade of Apistidae, Tetrarogidae, Synanceiidae, Aploactinidae, Congiopodidae s.l., Gnathanacanthidae, and Pataecidae [17, 21]. A distinctive parameter for the superfamily Scorpaenoidea sensu Imamura, 2004 is a presence of three epuralia, while Congiopodoidea has two, and Pataecidae has one [21].
Thus published morphological data point to the monophyly of Congiopodoidea, but different researchers come to conflicting conclusions regarding the sister group.
Genetic
In the Introduction notes the results of Smith and Wheeler WL [23], they analyzed the mitochondrial and nuclear genes: mtDNA (12S rDNA, 16S rDNA and tRNA-Val), nuclear DNA (28S rDNA, histone HS and TMO-4c4 ). In the constructed tree, the clade with Congiopodus peruvianus included, for example, Perca flavescens (Mitchill 1814), and the sister clade included, among others, Zanclorhynchus spinifer and Harpagifer kerguelensis Nybelin, 1947. This result should be considered a clear error and should be used only for illustration of still imperfections of genetic methods.
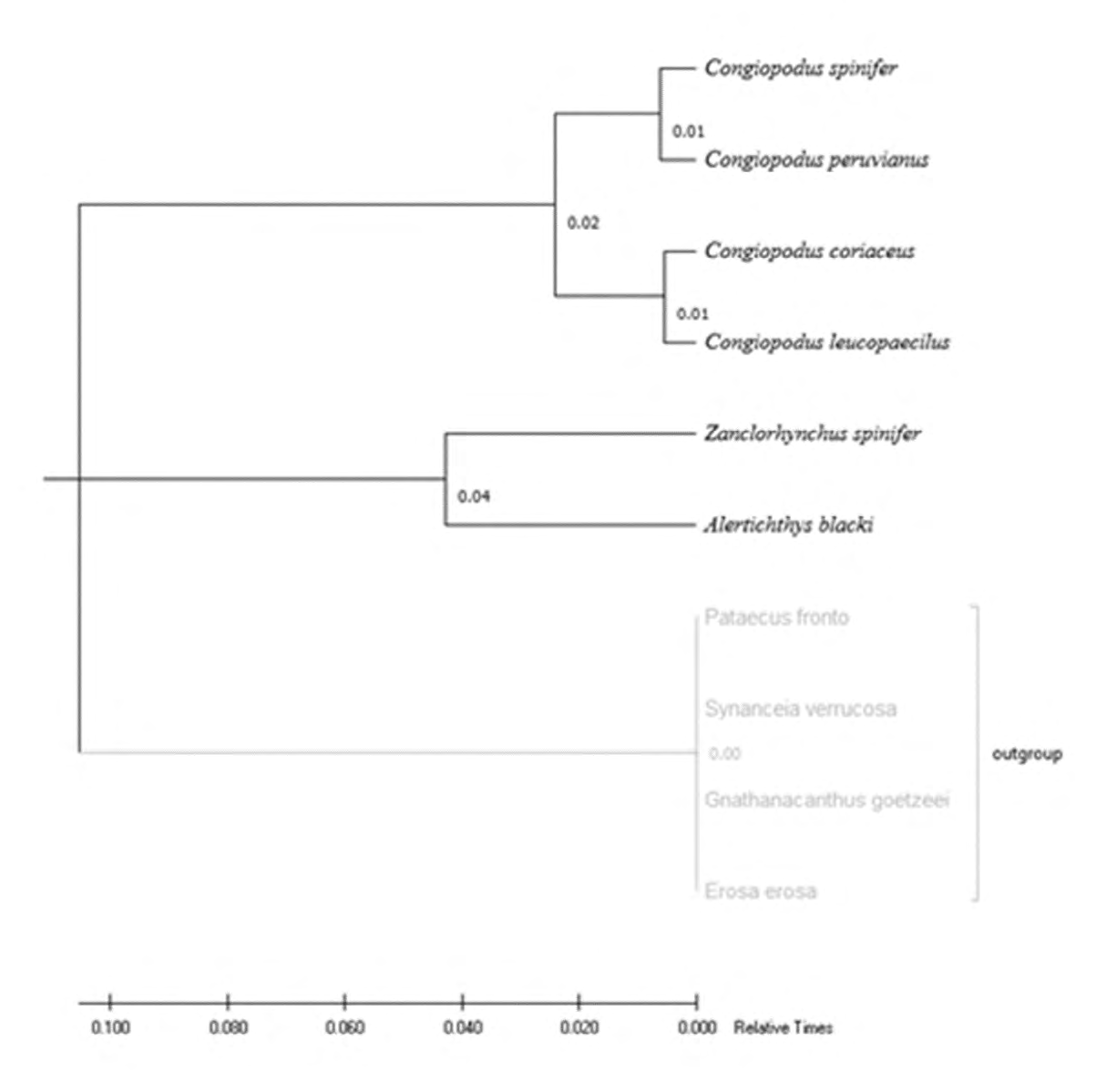
Zhukov’s M result [36] of comparison of nucleotide sequences of COI within the superfamily Congiopodoidea (Figure 4) showed a slightly different pattern from the results of Ishii and Imamura (Figure1). But it is the genetic result that shows the more explanatory geographic distribution of Congiopodus - C. coriaceus and C. leucopaecilus live sympatrically in Australia and New Zealand, while C. peruvianus and C. spinifer inhabit waters off southern South America and southern Africa, respectively. Two clades of Congiopodus may indicate an eastward distribution along the Circumantarctic Current, similar to the distribution of Zanclorhynchus [3], which cannot explain, for example, the external position of C. spinifer to other Congiopodus in the Ishii and Imamura cladogram (Figure 1).
The results of genetic studies are varied. Smith WL, et al. [25] provide some of them (Figure 5). It can be seen that the position of Congiopodoidea with different methods vary strongly. According to Smith, et al. [37], Congiopodoidea inserted into the clade with Neosebastidae, and, together with even Cottoidei, formed the sister group to the clade including Synanceiidae. Other researchers Lautredou AC [38] put Congiopodoidea in an external position to the clade, which includes not only Synanceiidae, but even Scorpaenidae. For Smith WL et al. [25] themselves, the result is that Congiopodoidea is even more distant from all Scorpaenoidei, the superfamily, together with two suborders Cottoidei and Zoarcoidei, formed a sister clade to Scorpaenoidei (Figure 6).
![Figure 5: Phylogenetic relationships in A - Scorpaenoidei and related groups [37], B - Scorpaeniformes and related groups [38]. (Figure 2).](/fulltextimages/9202/fig_5.png)
The results of Smith, et al. [25] correspond to the hypothesis of the taxonomic rank of the suborder, but one cannot ignore the many synapomorphies that unite Congiopodoidea and Synanceiidae sensu Smith et al., 2018. Undoubtedly, the described results need additional verification and, due to the wide range of interpretations, cannot be accepted without confirming studies.
Contradictions in the Results of Relations of Congiopodoidea and Proposals for their Correction
On the obtained phylogenetic trees (Figures 2&3), the sister branch to the superfamily Congiopodoidea includes a taxonomically complex group, now belonging to one family Synanceiidae [4, 39]. Previously [15, 40] the following systematic position of these taxa was considered: two genera Apistus and Tetraroge belonged to the corresponding subfamilies Apistinae Gill, 1859 and Tetraroginae Smith, 1949 within the family Scorpaenidae. The same family included Minous in the tribe Minoini, Inimicus and Choridactylus in the tribe Choridactylini, Synanceia and Erosa in the tribe Synanceini of the subfamily Synanceiinae Swainson, 1839. Gnathanacanthus and Pataecus in Gnathanacanthidae and Pataecidae, respectively. Imamura H [21] introduced a slightly different taxonomy, relegating Apistus and Tetraroge to separate families Apistidae and Tetrarogidae, respectively. But the unresolved contradiction is the family Synanceiidae, which simultaneously includes Synanceia and Erosa, Inimicus, Choridactylus, Minous, albeit in different subfamilies Synanceiinae, Inimicinae Gill, 1905 Choridactylinae Kaup, 1859 and Minoinae Jordan et Starks, 1904, respectively. Against this background, the decision to unite the entire sister to superfamily Congiopodoidea group into one family Synanceiidae Smith WL, et al. [25] is understandable and removes almost all contradictions. Here, only the inclusion of the genus Apistus in the same family looks erroneous. Its position is clearly basal both in the study by Imamura H [21] and by Voskoboinikova, et al. [1]. Considering, in turn, the entire group as part of the family Scorpaenidae, which additionally includes two subfamilies Zanclorhynchinae and Congiopodinae, is also erroneous. The morphological differences between Congiopodoidea and the rest of Scorpaenoidea sensu Imamura 2004 are so great that Mandritsa SA [10] proposed to separate Congiopodidae and Zanclorhynchidae into their suborder Congiopodoidei. Justifying the special position of Congiopodoidei sensu Mandritsa, 2001, he pointed out a number of characters that are absent in Scorpaenoidei sensu Imamura and Yabe, 2002:
- The infraorbital sensory canal in os lacrimale contains two neuromasts (three in Scorpaenoidei sensu Imamura and Yabe, 2002).
- Abdominal (trunk) vertebrae 14–19 (9–13 in Scorpaenoidei).
- Significant distance between os palatinum and the pterygoid bones.
- Main pore in praeoperculo-mandibular sensory canal is absent between os dentale and os anguloarticulare.
- Os dentale bears only 1–2 main pores (Scorpaenoidei usually has 4).
- Articular process on os praemaxillare not developed.
- Temporal fossa absent (Scorpaenoidei always has).
However, the erection of the group to a suborder has not received further support [15, 28, 40, 41, 42]. To remove contradictions, Apistus should be considered as belonging to its own family Apistidae, and the family Synanceiidae sensu Smith 2018 without Apistidae should be considered in the rank of the superfamily Synanceiidea with a corresponding increase in the rank of subfamilies to families. Also it is needed more studies to clarify the position of genus Erosa in the family Synanceiidae.
The proposed taxonomy of the group as part of the suborder Scorpaenoidei of the order Perciformes satisfies both the majority of genetic and morphological studies (the family Tetrarogidae can be further divided into two families, which will remove some contradictions in the results of molecular studies): superfamily Synanceiidea Swainson, 1839 sedis mutabilis family Apistidae Gill, 1859 genus Apistops Ogilby, 1911 genus Apistus Cuvier, 1829 genus Cheroscorpaena Mees, 1964 superfamily Synanceiidea Swainson, 1839 family Synanceiidae Swainson, 1839 incertae sedis genus Erosa Swainson, 1839 family Synanceiidae Swainson, 1839 genus Dampierosa Whitley, 1932 genus Leptosynanceia Bleeker, 1874 genus Pseudosynanceia Day, 1875
genus Synanceia Bloch et Schneider, 1801 genus Trachicephalus Swainson, 1839 family Tetrarogidae Smith, 1949 genus Ablabys Kaup, 1873 genus Centropogon Günther, 1860 genus Coccotropsis Barnard, 1927 genus Cottapistus Bleeker, 1876 genus Glyptauchen Günther, 1860 genus Gymnapistes Swainson, 1839 genus Liocranium Ogilby, 1903 genus Neocentropogon Matsubara, 1943 genus Neovespicula Mandrytsa, 2001 genus Notesthes Ogilby, 1903 genus Ocosia Jordan et Starks, 1904 genus Paracentropogon Bleeker, 1876 genus Pseudovespicula Mandrytsa, 2001 genus Richardsonichthys Smith, 1958 genus Snyderina Jordan et Starks, 1901 genus Tetraroge Günther, 1860 genus Vespicula Jordan et Richardson, 1910 family Choridactylidae Kaup, 1859 genus Choridactylus Richardson, 1848 genus Inimicus Jordan et Starks, 1904 family Minoidae Jordan et Starks, 1904 genus Minous Cuvier, 1829 family Aploactinidae Jordan et Starks, 1904 genus Acanthosphex Fowler, 1938 genus Adventor Whitley, 1952 genus Aploactis Temminck et Schlegel, 1843 genus Aploactisoma Castelnau, 1872 genus Bathyaploactis Whitley, 1933 genus Cocotropus Kaup, 1858 genus Erisphex Jordan et Starks, 1904 genus Kanekonia Tanaka, 1915 genus Matsubarichthys Poss et Johnson, 1991 genus Neoaploactis Eschmeyer et Allen, 1978 genus Paraploactis Bleeker, 1864 genus Peristrominous Whitley, 1952 genus Prosoproctus Poss et Eschmeyer, 1979 genus Pseudopataecus Johnson, 2004 genus Ptarmus Smith, 1947 genus Sthenopus Richardson, 1848 genus Xenaploactis Poss et Eschmeyer, 1980 family Gnathanacanthidae Gill, 1892 genus Gnathanacanthus Bleeker, 1855 family Pataecidae Gill, 1872 genus Aetapcus Scott, 1936 genus Neopataecus Steindachner, 1884 genus Pataecus Richardson, 1844 superfamily Congiopodoidea Gill, 1889 family Congiopodidae Gill, 1889 genus Congiopodus Perry, 1811 family Zanclorhynchidae Andriashev, 1993 genus Alertichthys Moreland, 1960 genus Zanclorhynchus Günther, 1880
Funding
The study was supported by the State Research Program number 122031100285-3.
Conflict Of Interest
The author declare that he have no conflict of interest.
References
-
Voskoboinikova OS, Zhukov MY (2021) Comparative osteology of the superfamily Congiopodoidea (Pisces: Scorpaenoidei). Zoomorphology 140: 373-385.
-
Roberts CD, Stewart AL, Struthers CD (2015) Family Congiopodidae. In: Roberts CD, Stewart AL, et al. (Eds.), The fishes of New Zealand. Te Papa Press, Wellington, New Zealand, pp: 1103-1106.
-
Zhukov MY (2019) Two new subscpecies of Antarctic horsefish (Zanclorhynchus, Scorpaeniformes: Congiopodidae) from Indian Sector of Southern Ocean. Tr Zool Inst RAS 323(4): 541-557.
-
Fricke R, Eschmeyer WN, Laan RVD (2022) Genera/ species by family/subfamily in Eschmeyer’s catalog of Fishes.
-
Paulin CD, Moreland JM (1979) Congiopodus coriaceus, a new species of pigfish, and a redescription of C. leucopaecilus (Richardson), from New Zealand (Pisces: Congiopodidae). New Zealand Journal of Zoology 6(4): 601-608.
-
Zhukov MY (2020) First record of the Deepsea pigfish Congiopodus coriaceus (Scorpaeniformes: Congiopodidae) in the Great Australian Bight with new data on morphology. Tr Zool Inst RAS 324(4): 476-484.
-
Hureau JC (1978) Notes sur la famille des Congiopodidae (Teleosteens, Perciformes): redecouverte de Zanclorhynchus spinifer Gunther. 1880, auxiles Kerguelen et rehabilitation de Congiopodus kieneri (Sauvage, 1878). Bulletin du Museum Natianal d’Histaire Naturelle 42(5): 1019-1026.
-
Pequeno G (1989) Peces de Chile. Lista sistemática revisada y comentada. Revista de Biologia 24(2): 1-132.
-
Moreland JM (1960) A new genus and species of congiopodid fish from southern New Zealand. Records of The Dominion Museum 3(3): 241-246.
-
Mandrytsa SA (2001) Seismosensory system and classification of scorpionfishes (Scorpaeniformes: Scorpaenoidei). Perm State Univ Press, Russia, pp: 393.
-
Balushkin AV, Zhukov MY (2016) Polytypy of the genus Zanclorhynchus (Zanclorhynchinae: Congiopodidae): description of the new species Z. chereshnevi sp. n. from the Indian sector of the Southern Ocean. J Ichthyol 56: 791-798.
-
Zhukov MY, Balushkin AV (2018) Description of a New Subspecies Zanclorhynchus spinifer heracleus subsp. nov. from the Ridge of Hercules (Pacific-Antarctic Ridge). J Ichthyol 58: 104-112.
-
Paxton JR, Hoese DF, Allen GR, Hanley JE (1989) Pisces Petromyzonidae to Carangidae. Australian Biological Resources Survey. 7: 1-665.
-
Poss GS (1994) Family Congiopodidae. In: The fishes of Australia’s south coast. In: Gomon MF, Glover JCM, et al. (Eds.). Adelaide, State Printer, pp: 525-526.
-
Nelson JS (2006) Fishes of the world. In: 4th (Edn.), New York, John Wiley & Sons, pp: 601-624.
-
Ishii N, Imamura H (2008) Phylogeny of the family Congiopodidae (Perciformes: Scorpaenoidea), with a proposal of new classification. Ichthyological Research 55: 148-161.
-
Honma Y, Imamura H, Kawai T (2013) Anatomical description of the genus Perryena, and proposal to erect a new family for it based on its phylogenetic relationships with related taxa (Scorpaeniformes). Ichthyological Research 60(2): 122-141.
-
Gunther A (1880) Report on the shore fishes. Scientific results of the voyage of H.M.S. Challenger during the years 1873-1876. Challenger Reports 1(6): 1-82.
-
Waite ER (1916) Fishes. Australasian Antarctic Expedition. Scientific reports. Series C 3(1): 1-92.
-
Norman JR (1937) Coast fishes. Part II. The Patagonian region. Discovery Reports 16: 1-150.
-
Imamura H (2004) Phylogenetic Relationships and New Classification of the Superfamily Scorpaenoidea (Actinopterygii: Perciformes). Species Diversity 9(1): 1-36.
-
Imamura H, Yabe M (2002) Demise of the Sсorpaeniformes (Actinopterii: Percomorpha): an altemative phylogenetic hypothesis. Bulletin of Fisheries Sciences, Hokkaido University 53(3): 107-128.
-
Smith WL, Wheeler WC (2004) Polyphyly of the mail- cheeked fishes (Teleostei: Scorpaeniformes): evidence from mitochondrial and nuclear sequence data. Molecular Phylogenetics and Evolution 32(2): 627-646.
-
Li B, Dettai A, Cruaud C, Couloux A, Meniger MD, et al. (2009) RNF213, a new nuclear marker for acanthomorph phylogeny. Mol Phylogenet Evol 50(2): 345-363.
-
Smith WL, Everman E, Richardson C (2018) Phylogeny and Taxonomy of Flatheads, Scorpionfishes, Sea Robins, and Stonefishes (Percomorpha: Scorpaeniformes) and the Evolution of the Lachrymal Saber. Copeia 106(1): 94- 119.
-
Greenwood PH, Rosen DE, Weitzman SH, Myers GS (1966) Phyletic studies of teleostean fishes, with a provisional classification of living forms. Bull Amer Mus Nat Hist 131: 339-456.
-
Nelson JS (1976) Fishes of the world. New York: John Wiley & Sons, pp: 416.
-
Lloris D, Rucabado J (1991) Ictiofauna del canal Beagle (Tierra de Fuego), aspectos ecologicos y analisis biogeografico. Ministerio de Agricultura, Pesca y Alimentacion. Secretaria General Tecnica, Madrid 8: 1-182.
-
Ishida M (1994) Phylogeny of the suborder Scorpaenoidei (Pisces: Scorpaeniformes). Bulletin of the Nansei National Fisheries Research Institute 27: 1-112.
-
Agassiz L (1834) Recherches sur les poisons fossils. Neuchatel 1: 16.
-
Roberts CD (1993) Comparative morphology of spined scales and their phylogenetic significance in the teleostei. Bulletin of Marine Science 52(1): 60-113.
-
Hureau JC (1985) Congiopodidae. In: Fischer W, Hureau JC (Eds.), FAO species identification sheets for fishery purposes. Southern Ocean (Fishing areas 48, 58 and 88). Rome 2: 278-279.
-
Hollister G (1936) Caudal skeleton of Bermuda shallow water fishes. I. Order Isospondyli: Elopidae, Megalopidae, Albulidae, Clupeidae, Dussumieridae, Engraulidae. Zoologica 21(23): 257-290.
-
Gosline WA (1960) Contribution towards a classification of modern isospondylous fishes. Bulletin of the British Museum (Natural History) 6: 325-365.
-
Nybelin O (1963) Zur Morphologie und Terminologie des Schwanzskelettes der Actinopterygier. Arkiv för Zoologi 15(35): 485-516.
-
Zhukov MYu (2022) Phylogeny of the superfamily Congiopodoidea (Pisces: Scorpaenoidei) based on nucleotide sequences of the COI gene. International scientific conference Biodiversity protection and environmental problems of nature management 3: 49- 53.
-
Smith WL, Craig MT (2007) Casting the percomorph net widely: the importance of broad taxonomic sampling in the search for the placement of serranid and percid fishes. Copeia 2007(1): 35-55.
-
Lautredou AC, Motomura H, Gallut C, Costaz CO, Lecointre G, et al. (2013) New nuclear markers and exploration of the relationships among Serraniformes (Acanthomorpha, Teleostei): the importance of working at multiple scales. Molecular Phylogenetics and Evolution 67(1): 140-155.
-
Fricke R, Eschmeyer WN, Van der Laan R (2022) Eschmeyer’s Catalog of Fishes: Genera, Species, References.
-
Nelson JS, Grande TC, Wilson MVH (2016) Fishes of the World. In: 5th (Edn.), Hoboken: John Wiley and Sons, pp: 707.
-
Eschmeyer WN (1998) Catalog of fishes. San Francisco: Special Publication, California Academy of Sciences 3: 2905.
-
Laan RVD, Eschmeyer WN, Fricke R (2014) Family-group names of Recent fishes. Zootaxa 3882(2): 1-230.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework