Hemocytes Lysate Supernatant Derived Phenoloxidase Activity in the Hemolymph of Giant Freshwater Prawn Macrobrachium rosenbergii (De Man, 1879)
The aim of the present study attempted to find the cellular immune response in Macrobrachium rosenbergii by melanization reaction produced by hemocytes lysate supernatant (HLS) phenoloxidase (PO) activity. The substrate affinity of the PO enzyme was determined using different phenolic substrates and it was found that the diphenols and polyphenol were oxidized. Hence, the enzyme was characterized as a catechol oxidase type of PO and 3,4-dihydroxy-DL-phenylalanine (DL-DOPA) showed the highest substrate affinity to the HLS. The optimal enzyme activity was observed at 5 mM DL-DOPA in 10 mM Tris-HCl buffer at a pH of 7.5 at 25°C for 20 min and absorbance at 490 nm. Kinetic characteristics of HLS from the prawn were determined. Determination of optimal conditions of PO activity in the HLS has also been attempted. These results depicted that in the presence of PO and peroxidase inhibitors, phenylthiourea (PTU) and tropolone respectively have a decreased HLS PO activity. The determination of PO activity was also highly activated by trypsin, sodium dodecyl sulphate (6 mg.ml-1) and laminarin (4 mg.ml-1) enzyme expression. We also identified the chemicals causing in vitro inhibition or activation of the enzyme as an HLS of the freshwater prawn having a potent PO activity.
Introduction
Aquaculture is economically and nutritionally important in many countries, especially in India and China and other Southwest and Southeast Asian countries and it is an increasingly important source of animal protein [1]. The giant freshwater prawn, Macrobrachium rosenbergii (De Man, 1879) is an important crustacean species cultured in India. In the last few years, M. rosenbergii was familiar as a species with great aquaculture value [2]. The M. rosenbergii has excellent cultivation characteristics such as fast growth, better survival rate, adaptation, strong resistance to stress, disease susceptibility, tolerance to a wide range of temperatures, high market demand and high international export value [3, 4]. However, the production of M. rosenbergii also known as scampi is curtailed by diseases caused by opportunistic pathogens in the rearing environment. Resistance to diseases is based on yhe strategic improvement of the immune system of the animal and requires intensive research on immune components and their function [5, 6, 7]. As for any invertebrate, the innate immune mechanism in prawns includes humoral [5, 6, 7] and cellular components [8, 9, 10, 11, 12, 13].
The cellular immune components of crustaceans primarily include particular fixed cells such as branchial podocytes, nephrocytes, and circulating blood cells [14, 15, 16] or hemocytes including phagocytosis, encapsulation, nodule formation, cytotoxicity and exocytosis [12, 17, 18, 19] as well as hemocytes and hematopoietic cells [20].
The humoral immune components include a variety of soluble substances detectable in the plasma, serum, or hemocyanin such as agglutinins [6, 21, 22, 23, 24] and the prophenoloxidase (_pro_PO) system [5, 7], β-1,3-glucan and/ or bacterial lipopolysaccharide (LPS)-binding proteins [25, 26, 27], antimicrobial proteins [24, 28], hemolytic system [29, 30], antiproliferative [31], antitumor or anticancer [32], antimicrobial peptides [33] and antifungal proteins [26, 34], as well as cytotoxic molecules [35]. The cellular and humoral immunity of crustaceans are closely related and mutually complementary [36].
In the humoral defense mechanism, the secretion of antimicrobial peptides is considered to be one of the important steps in defense against pathogenic infections [33]. The other important crustacean defense systems include enzymes and proteins in the prophenoloxidase (pro_PO) activation and blood coagulation systems. Earlier research indicated that the _M. rosenbergii hemocytes are the main target cells of Spiroplasma eriocheiris and they are very important for systematic infection in the prawn body [37]. So hemocytes are typically used to study the immune responses of crustaceans [18, 38, 39].
Crustacean hemocytes play an important role in the host’s innate immune responses. They are involved in cellular immune responses, such as phagocytosis, encapsulation, and mediation of cell-cell communication and also in humoral immune responses, such as proteolytic cascades [40, 41]. The general classification of hemocyte cells in crayfish, penaeid shrimps, prawns and crabs is based on the presence or absence of cytoplasmic granules [42, 43, 44, 45]. Additionally, hemocytes are involved in phagocytosis of small microbes, nonself recognition [18, 46], encapsulation of parasites [47], elimination of reactive oxygen intermediates [48], in vitro generation of superoxide anion [49] and agglutinin-mediated phagocytosis-associated generation of superoxide anion and nitric oxide [12, 18].
Invertebrate animals lack an adaptive immune response and are therefore completely dependent on their innate immune systems for host defense. Melanization, which is a major innate defense system in invertebrates, is controlled by the enzyme phenoloxidase [5, 7, 50, 51]. The active PO is a bifunctional enzyme that catalyzes the o-hydroxylation of monophenols to catechols and the oxidation of o-diphenols into o-quinones [52].
For a long period, it has been recognized that the pro_phenoloxidase (_pro_PO) activating system has an important role as a non-self recognition system in invertebrates especially crustaceans [51], and participates in the innate immune response through melanization, cytotoxic reactions, cell adhesion encapsulation, and phagocytosis [18, 19, 53, 54, 55, 56]. Phenoloxidase catalyses the hydroxylation of monophenols to _o-diphenols (monophenoloxidase or cresolase activity) and the oxidation of o-diphenols to o-quinones (diphenoloxidase activity) leading to the synthesis of melanin [57, 58]. In addition to its role in the immune response [5, 6, 7] and the formation of melanin is also involved in other important physiological processes such as wound healing and sclerotization [57].
Proteins with those enzymatic activities are ubiquitously distributed in bacteria, fungi, plants, and vertebrates, for example, tyrosinases and polyphenoloxidases (PPO) with both mono and diphenolase activities, and catecholoxidase (CO) with only diphenolase activity [59, 60]. The first reaction involves monophenolase activity, which converts tyrosine to 3,4-dihydroxy-L-phenylalanine (L-DOPA) which is then oxidized to quinone by POs diphenolase activity [52, 61]. The resulting quinones are converted to melanin by a series of intermediate steps involving enzymatic and non-enzymatic reactions [62, 63]. In one of these enzymatic reactions, dopachrome is decarboxylated by dopachrome isomerase (also called dopachrome tautomerase or Dopachrome Conversion Factor) to form dihydroxy indole, which is then converted to melanin [64, 65].
In crustaceans and insects, phenoloxidase usually exists as a non-active zymogen, prophenoloxidase (_pro_PO), whose activation to the PO form is tightly regulated via an enzymatic cascade because the melanization reaction generates toxic compounds such as quinone species. This cascade is triggered by the presence of several microbial cell wall components such as β-1,3-glucan, lipopolysaccharides, and peptidoglycan [56]. There is a detectable or high amount of PO activity in crustaceans plasma [5, 7, 66, 67] that could be derived from _pro_PO released from hemocytes [68] or from hemocyanin [5] to melanization activity still remains unclear.
According to the results of our previous research, the hemocyanin showed PO activity in freshwater prawns and appears to be enhanced with activators such as proteases, SDS, and LPS [5], agglutination activity with vertebrate red blood cells (RBC) [6] and optimal condition of plasma PO activity [7]. The present study attempted to characterize HLS PO activity in terms of absorbance maxima, substrate specificity, time interval, optimum ionic strength, pH and temperature in order to determine the biochemical and physiological conditions that support enzyme activity.
Furthermore, to understand the substrate affinity of the HLS PO enzyme activity the kinetics of the enzymes rate of reaction was determined in the Lineweaver-Burk plot. It is evident that the kinetics of the crustaceans phenoloxidases vary among the different components of the hemolymph as well as species [7, 8, 69, 70, 71, 72]. Moreover, to our knowledge, there is no information on the optimized conditions of the HLS phenoloxidase enzyme. Hence, an attempt has been made to optimize the conditions for determining PO activity of HLS including Km and Vmax value of freshwater prawn M. rosenbergii. Based on the determination of activation the PO activity in trypsin, SDS and laminarin has also been attempted.
Materials and Methods
Source of Experimental Prawns
Adult intermoult of the giant freshwater prawn, M. rosenbergii weighing around 30-40 ± 2.26 g was collected from private hatcheries in and around Chennai, India. In the laboratory, the prawns were maintained in 500 L FRP tanks containing continuously aerated freshwater. The animals were fed with egg white ad libitum and were acclimatized to the laboratory conditions for at least 4 to 5 days prior to use. Fifty percent of water was renewed daily to remove the unfed and fecal materials. The uninjured, intermoult animals were used throughout this study.
Hemolymph Collection and Preparation of HLS
To 100 µl of hemolymph was collected by cardiac puncture using a 23G needle attached to a clean, sterile plastic syringe containing 1.9 ml of ice-cold iso-osmotic buffer (TBS: Tris- 50 mM, NaCl 210 mM, KCl 5 mM, MgCl2 2.5 mM; pH 7.5, 480 mOsm), following [5]. Rapidly the syringe was gently rotated to enable thorough mixing of the hemolymph sample with iso- osmotic buffer and immediately transferred to a pre-chilled polypropylene tube, centrifuged at (161 x g for 8 min at 4 °C). The hemocyte sediment was washed and resuspended in 2 ml of iso-osmotic buffer (TBS: pH 7.5, 480 mOsm), homogenized by ultrasonicator (4 pulses of 30 s, 4 °C). The resulting cell homogenate was centrifuged at 15000 x g for 20 min at 4 °C and the hemocyte lysate supernatant (HLS) was collected as a clear supernatant and stored at −80 °C.
Determination of Protein
The protein content in the HLS was determined according to Bradford MM [73] using bovine serum albumin as the standard. All chemicals during this and other assays were obtained from Sigma-Aldrich (St. Louis, MO, USA).
Oxidation of Phenolic Substrates by HLS
We tested the oxidative activity of 50 µl HLS (mg.ml-1) by incubating with 950 µl of different phenolic substrate solutions (Tris-HCl 10 mM, pH 7.5; 5 mM L-tyrosine, tyramine, L-DOPA, DL-DOPA, dopamine, catechol, hydroquinone, and pyrogallol) for 20 min at 25 °C. The colour developed was measured spectrophotometrically (UV-160A spectrophotometer, Shimadzu, Kyoto, Japan) at 300–800 nm against a reagent blank in which particular substrates were substituted for HLS.
Optimal Conditions for PO Measurement of HLS
The oxidative activity was tested by incubating 50 µl of HLS (mg.ml-1) in 950 µl of DL-DOPA (Tris-HCl 10 mM, pH 7.5) with different concentrations of DL-DOPA (1.0–10.0 mM) for 20 min at 25 °C. The colour developed was measured spectrophotometrically at 490 nm against a reagent blank (DL-DOPA). We similarly measured color development way in all further tests using 50 µl of HLS (mg.ml-1) with 950 µl of 5 mM DL-DOPA (Tris-HCl 10 mM, pH 7.5) incubated for different time intervals of 60–70 min at 25 °C. The effect of ionic strength of the buffer on oxidation of 5 mM DL-DOPA by HLS (mg.ml-1) was assessed by incubating 50 µl of HLS with 950 µl of DL-DOPA prepared in Tris-HCl buffer (pH 7.5) with different ionic strength (5–100 mM) incubated for 20 min at 25 °C. The ability of oxidation of 5 mM DL-DOPA at different pH values was tested by incubating 50 µl of HLS (mg.ml-1) with 950 µl of substrate solution prepared in Tris-HCl buffer (10 mM) at pH 6.0–9 incubated for 20 min at 25 °C. The effect of different temperatures was tested by incubating 50 µl of HLS (mg.ml-1) with 950 µl solutions of the substrate (5 mM DL-DOPA) prepared in 10 mM Tris-HCl (pH 7.5) buffer at temperatures of 10–80 °C incubated for 20 min.
Kinetic Parameters, Km and Vmax of HLS Phenoloxidase Enzyme
To measure the kinetic parameters of the HLS PO enzyme, different concentrations of DL-DOPA (1 - 10 mM) were mixed with 50 µl of HLS incubated for 20 min and absorbency read at 490 nm. Michaelis-Menten constant was estimated by plotting substrate concentrations [S] and rate of PO activity [V]. Lineweaver-Burk plot was plotted as reciprocal of substrate concentration [1/S] and rate of PO activity [1/V]. The resultant plot gave a line that intercepted X-axis to give the -1/Km value and intercepted Y axis to give the 1/Vmax. The slope Km/Vmax was determined and the resultant plot was rechecked from equation Y= mx + c.
Inhibition of PO Activity
In this experiment, 50 µl of HLS (mg.ml-1) with an equal volume of an inhibitors 3 mM phenylthiourea (PTU) or 16 mM tropolone containing 16 mM H2O2 was pre-incubated for 5 min at 25 °C. An aliquot of 100 μl of these reaction mixtures from control or experimental was incubated with 0.9 ml of 5 mM DL-DOPA for 20 min at 25 °C. After incubation, the colour developed was measured spectrophotometrically at an absorbance of 490 nm against a reagent blank of substrate solution (DL-DOPA).
Activation of PO Activity
In this experiment, 50 μl of HLS (mg.ml-1) was separately mixed with 50 μl of trypsin (1-10 mg.ml-1), SDS (1-10 mg.ml-1) and laminarin (1-10 mg.ml-1). In control, these chemicals were substituted with Tris-HCl buffer (10 mM; pH 7.5), and pre-incubated for 5 min at 25 °C. An aliquot of 100 μl of the reaction mixture from control or experimental was incubated with made up to 1 ml of 5 mM DL-DOPA for 20 min at 25 °C. The optical density of both control and experimental was measured spectrophotometrically at 490 nm against a blank reagent of DL-DOPA.
Statistical Analysis
The data were expressed as mean ± SD of triplicate experimental trials from five determinations. The variation between experimental trials and control were evaluated by SPSS/18 software (version 20; SPSS, New York) by one-way analysis of variance (ANOVA) at 0.01, 0.001 probabilities (P < 0.01, *P < 0.001).
Results
Effect of PO Activity with Various Substrates
The HLS separated from the hemolymph of freshwater prawn M. rosenbergii showed the highest activity with DL- DOPA (490 nm) when compared to L-DOPA (490 nm), dopamine (450 nm), catechol (480 nm), and pyrogallol (510 nm). In contrast, the monophenols, including tyramine and L-tyrosine and polyphenol hydroquinone failed to show any oxidation by the HLS. Because the highest oxidative activity was obtained with DL-DOPA, this substrate was used to detect PO activity in all subsequent trials (Figure 1).
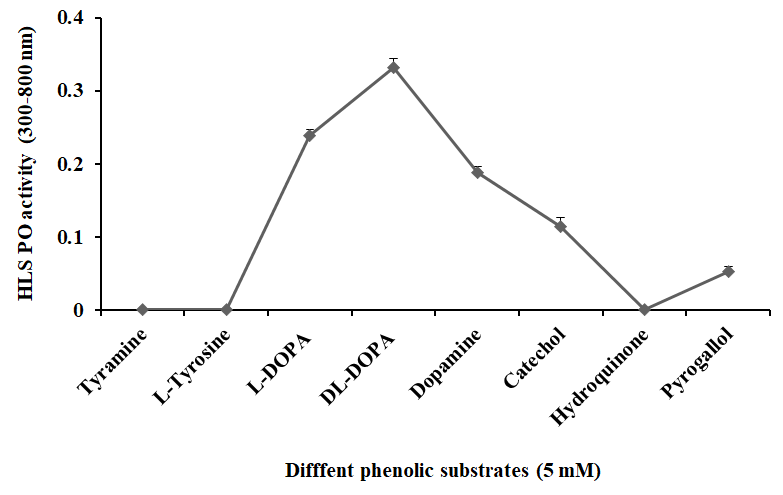
Figure 1: Phenoloxidase (PO) activity of the HLS of freshwater prawn Macrobrachium rosenbergii on different phenolic substrates (5 mM) in Tris buffer (Tris 10 mM, pH 7.5) incubated at 25 °C for 20 min and absorbance at 300–800 nm. The PO activity in optical density obtained at absorbance maxima of respective substrates. Data represent mean of triplicate repeats of five determinations (mean ± SD) in the same way in all further experiments.
Effect on Optimal Conditions for PO Activity
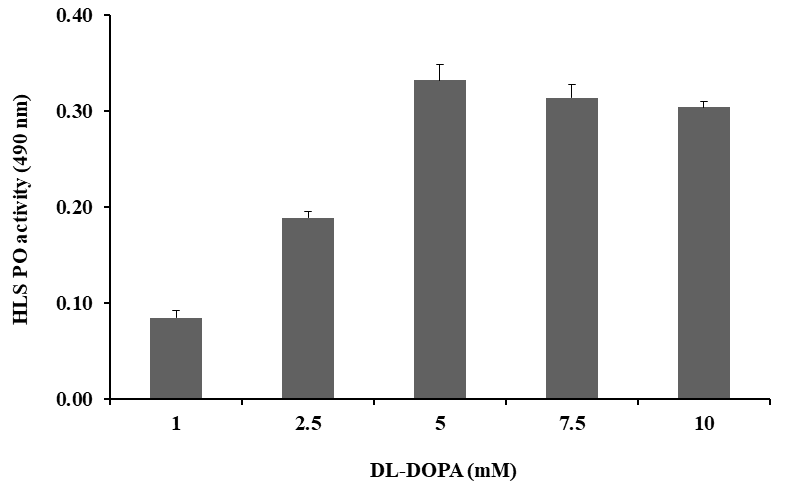
The HLS phenoloxidase activity increased with 1.0–10 mM concentrations of DL-DOPA. Of the tested concentrations,
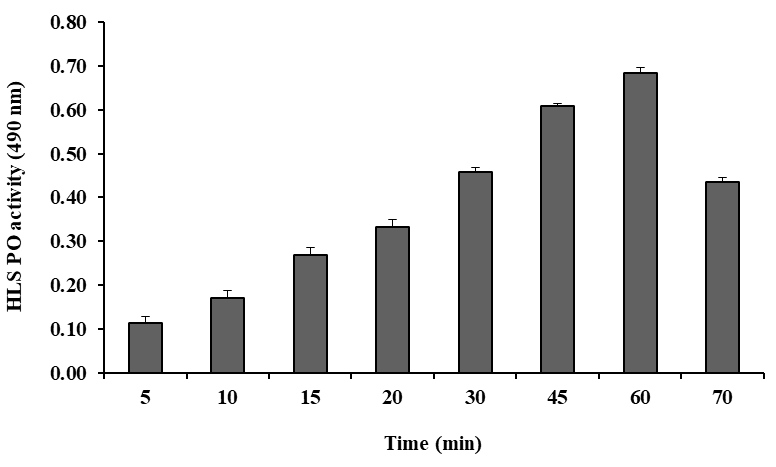
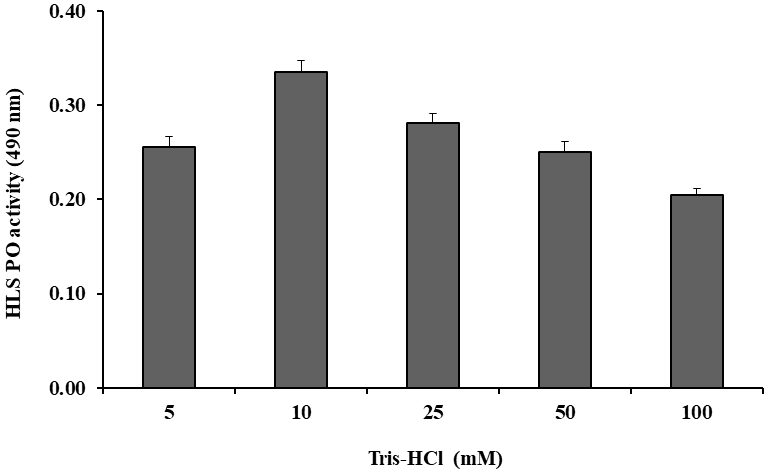
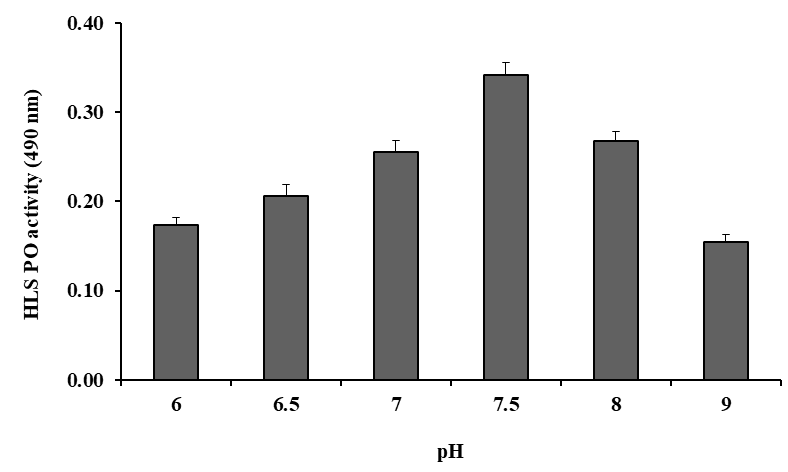
5 mM was taken as the optimal concentration and was used for further tests (Figure 2). The PO activity was also tested by HLS at various time intervals (5–70 min). The highest PO activity was found at 60 min, which was determined as the optimal incubation time was taken for 20 min (Figure 3). PO activity was also tested with Tris-HCl buffer (pH 7.5) of different ionic strengths (5–100 mM) and the highest activity was observed at 10 mM. This ionic strength was used as buffer concentration for further tests (Figure 4). Similarly, PO activity was tested with DL-DOPA of different pH values (6.0–9.0) of Tris-HCl buffer (10 mM). The highest PO activity was observed at pH 7.5, decreasing above and below a pH of
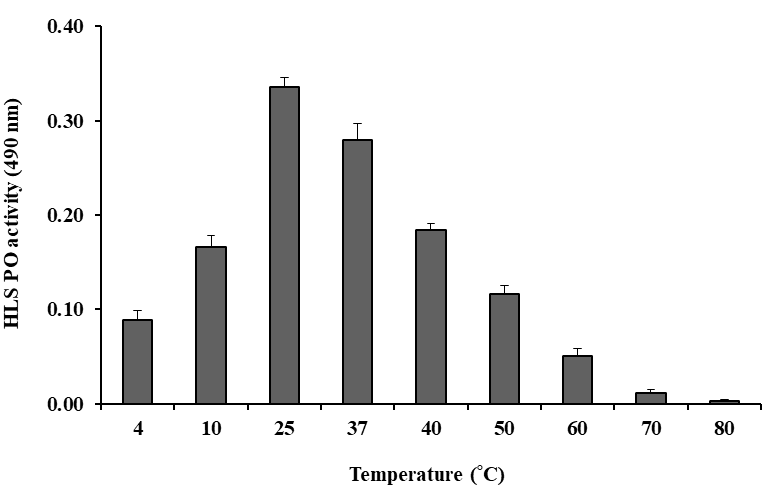
7.5 (Figure 5). The HLS samples were incubated for 20 min at different temperatures ranging from 4–80 °C. Stable and peak PO activity was observed at 25 °C, so this temperature was considered optimal (Figure 6). In summary, the optimal conditions for measuring HLS PO activity on DL-DOPA (5 mM) were 10 mM Tris-HCl, pH 7.5 at 25 °C for 20 min at 490 nm.
Kinetic Behavior
The kinetic characteristics of HLS PO activity were determined from the rate of reactions as calculated from oxidation of DL-DOPA at different concentrations (1 – 10 mM) in 20 min. The Michaelis-Menten constant Km was calculated to be 1.1 and the maximum velocity (Vmax) as 0.45 as shown in Figure 7A. Application of Km and Vmax yielded Lineweaver- Burk plot with a line slope (1.55) which on extrapolation intercepted at -0.4 that was plotted as -1/Km and on Y-axis 1/ Vmax was derived at 1.74 on X-axis (Figure 7B).
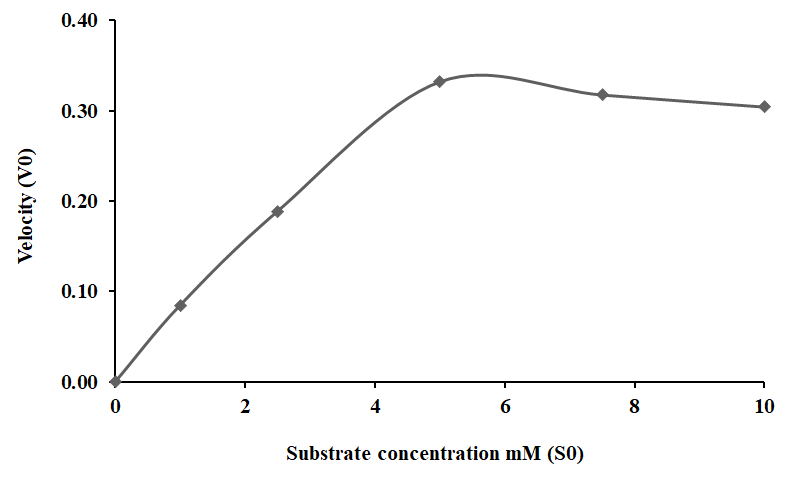
Figure 7A: Kinetic properties of PO activity in HLS of freshwater prawn M. rosenbergii at different substrate concentrations of DL-DOPA as shown in Michaelis–Menten curve.
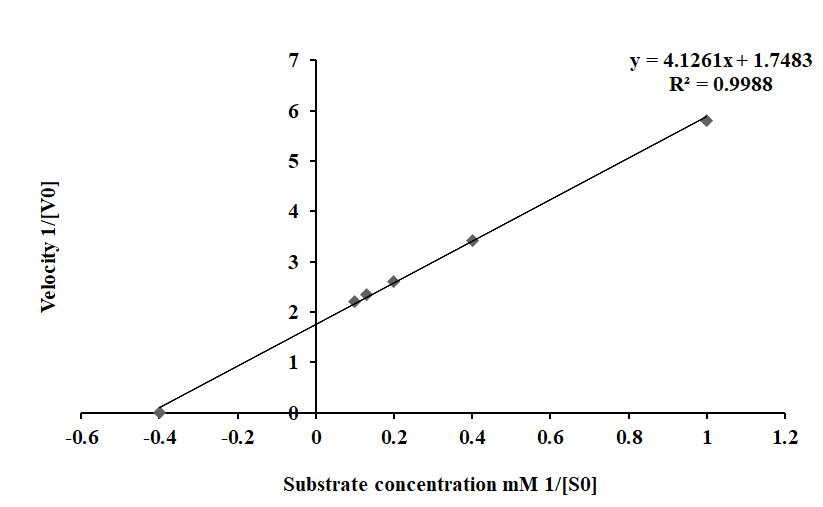
Figure 7B: The Km and Vmax values are calculated using Lineweaver Burk plot of PO activity in HLS of freshwater prawn M. rosenbergii with DL-DOPA as substrate.
Inhibition of HLS PO Activity
Pre-treatment of HLS with PTU (3 mM) decreased the oxidation of 5 mM DL-DOPA compared to control and the reduction was found to be about 85.62% statistically significant (p < 0.01). However, pre-treatment of HLS with tropolone, another strong inhibitor of PO activity, also showed a significant reduction in the oxidation of DL-DOPA when compared to control and the reduction in the phenoloxidase with HLS (77.24%) was statistically significant (p < 0.01) as shown in Figures 8 & 9.
![Figure 9: Effect of activators [(A) trypsin, (B) SDS, (C) laminarin] on activation of PO activity of the HLS of freshwater prawn M. rosenbergii.](/fulltextimages/9260/fig_9.png)
Figure 8: Inhibition of PO activity in HLS of freshwater prawn M. rosenbergii and conformation of PO activity using inhibitors (PTU and tropolone). Asterisk indicates significant variation determined from the value obtained for a treatment to untreated control by one way ANOVA at P < 0.01; *P < 0.001. Data represents mean of triplicate repeats of five determinations (mean ± SD).
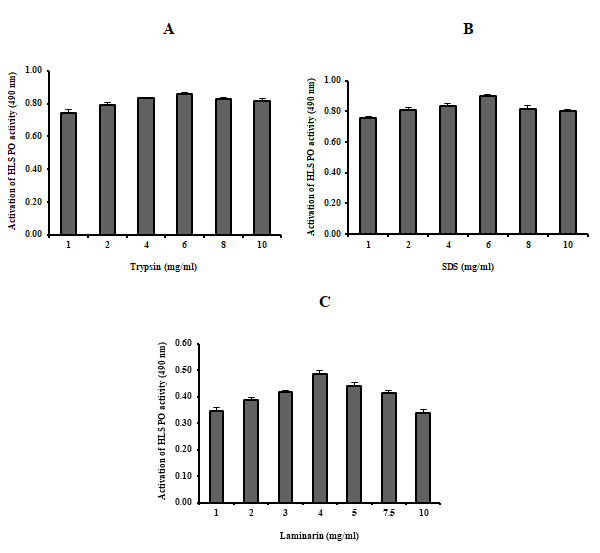
Effect of Activation on HLS PO Activity
The HLS was tested with trypsin, SDS and laminarin, and the PO activity was found to be higher with trypsin (6 mg.ml-1), SDS (6 mg.ml-1) and laminarin of 4 mg.ml-1 concentration than that of other concentrations as shown in the Figure 9A-C. However, these results clearly suggested that the optimum concentration for activating PO activity is 6 mg.ml-1 of trypsin and SDS, and laminarin for 4 mg.ml-1.
Discussion
The hemocyanin showed PO activity in the freshwater prawn M. rosenbergii and appears to be enhanced with activators such as proteases, detergent, and lipopolysaccharides [5]. Reports about the determination of plasma and hemocyanin PO and agglutination activity were predictable [6, 7]. Considering the complexity of crustacean immune defenses, our study attempted to explain the variance in the immune function of PO activity in the HLS of M. rosenbergii. In most crustacean species active PO is a bifunctional enzyme that catalyzes o-hydroxylation of monophenols to diphenols and then oxidizes o-diphenols into o-quinone [5, 7, 59, 62, 72]. The substrate affinity of HLS PO activity was attempted with monophenols, di-phenols or polyphenols. The results clearly suggested that the HLS showed that highest substrate affinity with diphenols and among the diphenol, DL-DOPA exhibited the highest PO activity suggesting catechol oxidase activity. The earlier report showed PO activity in the HLS of freshwater prawn M. rosenbergii using L-DOPA [8]. Our results clearly revealed that the highest activity was shown by diphenolic substrate DL-DOPA and not by L-DOPA.
Biochemical studies were undertaken in order to describe the optimum condition of the HLS PO activity. The enzyme reaction was observed with different concentrations of DL- DOPA. There was a steady increase in the enzyme activity from 5 mM concentration of DL-DOPA after which an increase in substrate concentration did not enhance the enzyme activity clearly proving substrate inhibition as the cause of the decline in enzyme activity. For better understanding, the PO activity was measured spectrophotometrically under different wavelengths (300-800 nm) due to the formation of brown pigment DL-DOPA at 20 min. The highest PO activity was found to be at a wavelength of 490 nm. The value shows a resemblance with shrimp Penaeus paulensis [74], freshwater prawn M. rosenbergii [8] and many other decapod crustaceans along with some variations observed such as in M. malcolmsonii [75] and mud crab Scylla olivacea [16].
Time is an important factor for PO enzyme activity that can either enhance or decline it. In the present study, the oxidation of the substrate (DL-DOPA) was tested at various time intervals from 5-60 minutes and the activity was observed to be high at 20 minutes of incubation. The hemocyte of M. rosenbergii showed optimum PO activity at one minute of L-DOPA [8], 40 minutes in crab Charybdis japonica [70], 30 minutes in shrimp P. paulensis and mud crab Scylla olivacea [16, 74, 75] and 20 minutes in spiny lobster Panulirus argus [68]. The differences in time interval optima in substrate variation clearly suggest a difference in PO characteristics.
Since PO is an enzyme and its activity depends on the steady state of the active sites which are necessary for substrate binding and subsequent activity. The optimum ionic interactions were studied by taking the HLS in different ionic strength of Tris-HCl buffer and PO activity was determined. The optimum ionic strengths of 10 mM Tris-HCl was found to show the highest PO activity and was used as a buffer for the study. To continue ionic interactions the optimum pH of the buffer required for HLS PO activity was also undertaken. The optimum pH was observed at pH 7.5 (brown colour formation of dopachrome) which was the same as that of purified Charybdis japonica PO [70] against DL-DOPA, Penaeus chinensis [76] and anomuran crab Albunea symmysta [72], but different from that of brown shrimp Penaeus californiensis which showed optimum pH at 8.0 [77], spiny lobster P. argus at pH 8.0 [68], mud crab Scylla olivacea at pH 8.0 [16] and Penaeus setiferus at pH 7.5 [78]. The differences in optimum pH may be correlated with the species specificity.
Like time, the temperature is also an important factor that can either enhance or decline enzyme activity. In the present study, the optimum temperature of HLS PO activity in M. rosenbergii was determined by incubating HLS at various temperatures ranging from 10-90 °C. The optimum temperature was found to be at 25 °C which lowered to 37 °C and then the enzyme activity declined to 90 °C. The precise temperature of 25 °C was found to be optimum for PO activity in DL-DOPA in the serum of anomuran crab A. symmysta [72]. The previous report differs from the present study that hemocytes of M. rosenbergii showed optimum PO activity in L-DOPA at 37 °C [8]. The shrimp powders from Penaeus duorarum and Penaeus setiferus showed an optimum temperature of 40 °C and 45 °C for PO activity in DL-DOPA [79] and polyphenoloxidase (PPO from the carapace powder of P. argus exhibited greater stability at 35 °C [69]. However, the spiny lobster P. argus correlated with the present study showing an optimum temperature of 25 °C [68]. The differences in temperature optima in HLS (hemocytes) substrate and species specific clearly suggest a difference in PO characteristics among different species of Crustacea can be attributed to existing physiological conditions.
The enzyme kinetics of the HLS PO activity was determined using the Michaelis-Menten curve by plotting various concentrations of DL-DOPA (1-10 mM) and the rate of reaction was determined in 20 min (1/V). The initial rate of reaction increased up to a maximum reaction velocity after which it stabilized and then declined. The Km value determined for substrate enzyme affinity was 2.2 mM and this suggested a strong affinity between the enzyme and DL- DOPA and the Vmax was calculated as 0.45. The Lineweaver- Burk plot showed a slope of 0.45 with a correlation coefficient of R2 = 0.998. This indicated that the enzyme had active sites to maintain a steady increase in the rate of reaction. The kinetic and biochemical characteristics of the HLS and plasma PO activity demonstrate a distinct PO activity among the other crustaceans [70, 72, 77].
Comparative inhibition studies with the PTU and tropolone were made to confirm the PO activity in the HLS. The specific o-diphenoloxidase activity inhibitor, PTU inhibited the enzyme activity drastically in the HLS and also peroxidase enzyme inhibited tropolone confirm the phenoloxidase activity in HLS. The inhibition studies clearly revealed that the HLS showed PO activity. Phenylthiourea (PTU), known as a chelating reagent of copper [80], effectively inhibited the HLS PO activity suggesting that PO from M. rosenbergii prawn has copper in its active site. The previous report demonstrated inhibition of anomuran crab A. symmysta serum was high at a 7 mM concentration of PTU was added to the incubation mixture to completely inhibit the activity of the phenoloxidase [72]. The Allogamus auricollis hemolymph cell-free fraction increased in absorbance and stopped immediately when this specific inhibitor (PTU) was added 5 min after the initiation of the assay [81]. Furthermore, the observed oxidation of DL-DOPA was not due to peroxidase since tropolone which inhibited PO activity in the HLS did not act as a substrate for peroxidase in the presence of H2O2 [5, 7, 82, 83].
Interestingly, our research focused on the effect of activators on protease like trypsin and detergent like SDS and fungal cell wall laminarin among which trypsin, SDS and laminarin showed enhancement of PO activity in the HLS of M. rosenbergii. Our results were in accordance with the earlier report using trypsin as an activator with the L-DOPA tested in HLS PO activity increased in prawn M. rosenbergii [8], but our results clearly demonstrated diphenolic substrate DL-DOPA only exhibited the highest PO activity in the HLS, not L-DOPA. However, our previous report about the plasma (non-cellular) of the freshwater prawn M. rosenbergii revealed that the highest PO activity was by L-DOPA [5, 7]. However, for the shrimp P. paulensis, most of the PO activity (more than 90%) was found in the HLS [74].
Conclusion
The immunological function of phenoloxidase observed in HLS of M. rosenbergii appears to enhance resistance against various diseases, and investigation of PO activity in HLS protein revealed catechol oxidase type PO. In contrast to the previous reports about the conversion of tyrosine to L-DOPA in monophenolase activity, our study revealed a new mechanism in the conversion ratio that is higher than diphenol DL-DOPA. So, we conclude that initially tyrosine is transformed to DL-DOPA which is then oxidized to quinone by phenoloxidase diphenolase activity. Finally, the resulting quinones are converted to melanin by a series of reactions. These results clearly signify the diversity of PO activity in cellular immune response in various crustacean species. The finding provides evidence that the HLS of M. rosenbergii act as a potent immune system with the ability to cellular enzymatically functions as phenoloxidase.
Conflict of Interest
The authors declare that they have no competing of interest.
References
-
Habib A, Das NG, Hossain MB (2014) Growth performance and survival rate of _Macrobrachium rosenbergii_ (De Man, 1879) larvae using different doses of probiotics. Pakistan J Biol Sci 17(7): 920-924.
-
Keysami MA, Mohammadpour M, Saad CR (2012) Probiotic activity of _Bacillus subtilis_ in juvenile freshwater prawn, _Macrobrachium rosenbergii_ (de Man) at different methods of administration to the feed. Aquacult Int 20: 499-511.
-
Indulkar ST, Belsare SS, Ranade AM, Raje PC (2007) Culture of giant fresh water prawn _Macrobrachium_ _rosenbergii_ with low inputs in Ratnagiri district of Maharashtra. Proc Zool Soc India 6(1): 1-5.
-
Moffitt CM, Cajas-Cano L (2014) Blue Growth: The 2014 FAO state of world fisheries and aquaculture. Fisheries 39(11): 552-553.
-
Sivakumar MR, Denis M, Shanthi S, Arumugam M (2017) Phenoloxidase activity in humoral plasma, hemocyanin and hemocyanin separated proteins of the giant freshwater prawn _Macrobrachium rosenbergii_. Int J Biol Macromol 102: 977-985.
-
Sivakumar MR, Denis M, Shanthi S, Arumugam M (2020) Agglutination of plasma, hemocyanin, and separated hemocyanin from the hemolymph of the freshwater prawn _Macrobrachium rosenbergii_ (De Man, 1879) (Decapoda: Caridea: Palaemonidae). J Crustac Biol 40(3): 309-315.
-
Sivakumar MR, Shanthi R (2022) Plasma and hemocyanin phenoloxidase derived from the hemolymph of giant freshwater prawn _Macrobrachium rosenbergii_ (De Man, 1879). Crustacea-Nano, Micro and Macro Studies.
-
Sung HH, Chang HJ, Her CH, Chang JC, Song YL (1998) Phenoloxidase activity of hemocytes derived from _Penaeus monodon_ and _Macrobrachium rosenbergii_. J Invertebr Pathol 71(1): 26-33.
-
Cheng W, Chen JC (2000) Effects of pH, temperature and salinity on immune parameters of the freshwater prawn _Macrobrachium rosenbergii_. Fish Shellfish Immunol 10(4): 387-391.
-
Sung HH, Hwang SF, Tasi FM (2000) Responses of giant freshwater prawn (_Macrobrachium rosenbergii_) to challenge by two strains of _Aeromonas_ spp. J Invertebr Pathol 76(4): 278-284.
-
Cheng W, Liu CH, Hsu JP, Chen JC (2002) Effect of hypoxia on the immune response of giant freshwater prawn _Macrobrachium rosenbergii_ and its susceptibility to pathogen _Enterococcus_. Fish Shellfish Immunol 13(5): 351-365.
-
Raman T, Arumugam M, Mullainadhan P (2008) Agglutinin_-_mediated phagocytosis_-_associated generation of superoxide anion and nitric oxide by the hemocytes of the giant freshwater prawn _Macrobrachium_ _rosenbergii_. Fish Shellfish Immunol 24(3): 337-345.
-
Kumaresan V, Rajesh P, Mukesh P, Arockiaraj J (2017) Impacts of environmental and biological stressors on immune system of _Macrobrachium rosenbergii_. Rev Aquac 9(3): 283-307.
-
Johnson PT (1987) A review of fixed phagocytic and pinocytotic cells of decapod crustaceans, with remarks on hemocytes. Dev Comp Immunol 11(4): 679-704.
-
Martin GG, Kay J, Poole D, Poole C (1998) _In vitro_ nodule formation in the ridgeback prawn, _Sicyonia ingentis_, and the American lobster, _Homarus americanus_. Invertebr Biol 117(2): 155-168.
-
Mangkalanan S, Sanguanrat P, Utairangsri T, Sritunyalucksana K, Krittanai C (2014) Characterization of the circulating hemocytes in mud crab (_Scylla olivacea_) revealed phenoloxidase activity. Dev Comp Immunol 44(1): 116-123.
-
Vázquez L, Alpuche J, Maldonado G, Agundis C, Pereyra Morales A, et al. (2009) Review: Immunity mechanisms in crustaceans. Innate Immun 15(3): 179-188.
-
Denis M, Sivakumar MR, Tamilarasan K, Karthigayani T, Prabu M, et al. (2017) Activation of phenoloxidase activity by humoral lectin in hemocytes of freshwater crab _Paratelphusa jacquemontii_. Int J Biol Macromol 97: 258-263.
-
Liu S, Zheng SC, Li YL, Li J, Liu HP (2020) Hemocyte mediated phagocytosis in Crustaceans. Front Immunol 11: 268.
-
Irene S, Fasterius E, Ekblom C, Söderhäll K (2022) Characterization of hemocytes and hematopoietic cells of a freshwater crayfish based on single-cell transcriptome analysis. iScience 25(8): 104850.
-
Yan F, Zhang Y, Jiang R, Zhong M, Hu Z, et al. (2011a) Identification and agglutination properties of hemocyanin from the mud crab (_Scylla serrata_). Fish shellfish Immunol 30(1): 354-360.
-
Mohanty J, Sahoo S, Badhe MR, Pillai BR, Sahoo PK, et al. (2020) Lectin like activity of hemocyanin in freshwater prawn _Macrobrachium rosenbergii_. Protein J 39(4): 358- 365.
-
Coates CJ, Costa Paiva EM (2020) Multifunctional roles of hemocyanins. Subcell Biochem 94: 233-250.
-
Zhao X, Qiao J, Zhang P, Zhang Z, Aweya JJ, et al. (2021) Protein diversity and immune specificity of hemocyanin from shrimp _Litopenaeus vannamei_. Front Immunol 12: 772091.
-
Lee SY, Wang R, Söderhäll K (2000) A Lipopolysaccharide- and beta-1,3-glucan-binding protein from hemocytes of the freshwater crayfish _Pacifastacus leniusculus_. Purification, characterization, and cDNA cloning. J Biol Chem 275(2): 1337-1343.
-
Destoumiéux Garzón D, Saulnier D, Garnier J, Jouffrey C, Bulet P, et al. (2001) Antifungal peptides are generated from the C terminus of shrimp hemocyanin in response to microbial challenge. J Biol Chem 276(50): 47070– 47077.
-
Yu XQ, Kanost MR (2003) _Manduca_ _sexta_ lipopolysaccharide-specific immulectin-2 protects larvae from bacterial infection. Dev Comp Immunol 27(3): 189-196.
-
Lee SY, Lee BL, Söderhäll K (2003) Processing of an antibacterial peptide from hemocyanin of the freshwater crayfish _Pacifastacus leniusculus_. J Biol Chem 278(10): 7927-7933.
-
Zhang Y, Yan F, Hu Z, Zhao X, Min S, et al. (2009) Hemocyanin from shrimp _Litopenaeus vannamei_ shows hemolytic activity. Fish Shellfish Immunol 27(2): 330- 335.
-
Yan F, Qiao J, Zhang Y, Zhou N, Liu Y, et al. (2011b) Hemolytic properties of hemocyanin from mud crab _Scylla serrata_. J Shellfish Res 50(3): 957-962.
-
Zheng L, Zhao XL, Zhang P, Chen C, Liu S, et al. (2016) Hemocyanin from shrimp _Litopenaeus vannamei_ has antiproliferative effect against HeLa cell _in vitro_. PLoS ONE 11(3): e0151801.
-
Liu S, Zheng L, Aweya JJ, Zheng Z, Zhong M, et al. (2017) _Litopenaeus vannamei_ hemocyanin exhibits antitumor activity in S180 mouse model _in vivo_. PLoS ONE 12(8): e0183783.
-
Tassanakajon A, Rimphanitchayakit V, Visetnan S, Amparyup P, Somboonwiwat K, et al. (2018) Shrimp humoral responses against pathogens: antimicrobial peptides and melanization. Dev Comp Immunol 80: 81- 93.
-
Iijima R, Kurata S, Shunji N (1993) Purification, characterization, and cDNA cloning of an antifungal protein from the hemolymph of _Surcophugu peregrinu_ (Flesh Fly) larvae. Dev Comp Immunol 268(16): 12055- 12061.
-
Söderhäll K, Wingren A, Johansson MW, Bertheussen K (1985) The cytotoxic reaction of hemocytes from the freshwater crayfish, _Astacus astacus_. Cell Immunol 94(2): 326-332.
-
Huang Y, Ren Q (2020) Research progress in innate immunity of freshwater crustaceans. Dev Comp Immunol 104: 103569.
-
Liang TM, Li XL, Du J, Yao W, Sun G, et al. (2011) Identification and isolation of a _Spiroplasma_ pathogen from diseased freshwater prawns, _Macrobrachium_ _rosenbergii_, in China: a new freshwater crustacean host. Aquaculture 318(1-2): 1-6.
-
Gai YC, Wang LL, Zha JM, Qiu L, Song LS, et al. (2009) The construction of a cDNA library enriched for immune genes and the analysis of 7535 ESTs from Chinese mitten crab _Eriocheir sinensis_. Fish Shellfish Immunol 27(6): 684-694.
-
Zhao D, Song SH, Wang Q, Zhang XW, Hu SN, et al. (2009) Discovery of immune-related genes in Chinese mitten crab (_Eriocheir sinensis_) by expressed sequence tag analysis of haemocytes. Aquaculture 287(3-4): 297-303.
-
Johansson MW, Keyser P, Sritunyalucksana K, Söderhäll K (2000) Crustacean haemocytes and haematopoiesis. Aquaculture 191(1-3): 45-52.
-
Cerenius L, Kawabata SI, Lee BL, Nonaka M, Söderhäll K (2010) Proteolytic cascades and their involvement in invertebrate immunity. Trends Biochem Sci 35(10): 575- 583.
-
Battistella S, Bonivento P, Amirante GA (1996) Hemocytes and immunological reactions in crustaceans. Ital J Zool 63(4): 337-343.
-
Sritunyalucksana K, Gangnonngiw W, Archakunakorn S, Fegan D, Flegel TW (2005) Bacterial clearance rate and a new differential hemocyte staining method to assess immunostimulant activity in shrimp. Dis Aquat Org 63(1): 89-94.
-
Giulianini PG, Bierti M, Lorenzon S, Battistella S, Ferrero EA (2007) Ultrastructural and functional characterization of circulating haemocytes from the freshwater crayfish _Astacus leptodactylus_: cell types and their role after in vivo artificial non-self challenge. Micron 38(1): 49-57.
-
Deyashi M, Chakraborty SB (2022) Cytomorphological characterization, classification and counting of haemocytes in freshwater crab, _Varuna litterata_ (Crustacea: Decapoda). Invertebr Zool 19(2): 120-134.
-
Vázquez L, Perez A, Millan D, Agundis C, Martin G, et al. (1997) Morphology of haemocytes from the freshwater prawn _Macrobrachium rosenbergii_. J Morphol 234(2): 147-153.
-
Martin GG, Poole D, Poole C, Hose JE, Arias M, et al. (1993) Clearance of bacteria injected into the hemolymph of the penaeid shrimp, _Sicyonia ingentis_. J Invert Pathol 62(3): 308-315.
-
Munoz M, Cedeno R, Rodriguez J, Van der Knaap WPW, Mialhe E, et al. (2000) Measurement of reactive oxygen intermediate production in haemocytes of the penaeid shrimp, _Penaeus vannamei_. Aquaculture 191(1-3): 89- 107.
-
Vidya N, Thiagarajan R Arumugam M (2007) _In vitro_ generation of superoxide anion by the hemocytes of _Macrobrachium rosenbergii_: Possible mechanism and pathways. J Exp Zool A Ecol Genet Physiol 307(7): 383- 396.
-
Liu H, Jiravanichpaisal P, Cerenius L, Lee BL, Söderhäll I, et al. (2007) Phenoloxidase is an important component of the defense against _Aeromonas hydrophila_ infection in a crustacean _Pacifastacus leniusculus_. J Biol Chem 282(46): 33593-33598.
-
Cerenius L, Söderhäll K (2021) Immune properties of invertebrate phenoloxidases. Dev Comp Immunol 122: 104098.
-
Yamamoto K, Yakiyama M, Fujii H, Kusakabe T, Koga K, et al. (2000) Expression of prophenoloxidase mRNA during silkworm hemocyte development. Biosci Biotechnol Biochem 64(6): 1197-1202.
-
Lee SY, Söderhäll K (2002) Early events in crustacean innate immunity. Fish Shellfish Immunol 12(5): 421- 437.
-
Jiravanichpaisal P, Lee BL, Söderhäll K (2006) Cell- mediated immunity in arthropods: Hematopoiesis, coagulation, melanization and opsonization. Immunobiology 211(4): 213-236.
-
Söderhäll K, Cerenius L (1998) Role of the prophenoloxidase-activating system in invertebrate immunity. Curr Opin Immunol 10(1): 23-28.
-
Cerenius L, Söderhäll K (2004) The prophenoloxidase- activating system in invertebrates. Immunol Rev 198(1): 116-126.
-
Sugumaran M (2002) Comparative biochemistry of eumelanogenesis and the protective roles of phenoloxidase and melanin in insects. Pigment Cell Res 15(1): 2-9.
-
Nappi AJ, Christensen BM (2005) Melanogenesis and associated cytotoxic reactions: Applications to insect innate immunity. Insect Biochem Mol Biol 35(5): 443- 459.
-
Decker H, Jaenicke E (2004) Recent findings on phenoloxidase activity and antimicrobial activity of hemocyanins. Dev Comp Immunol 28(7-8): 673-687.
-
Kanteev M, Goldfeder M, Fishman A (2015) Structure- function correlations in tyrosinases. Protein Sci 24(9): 1360-1369.
-
Bai G, Brown JF, Watson C, Yoshino TP (1997) Isolation and characterization of phenoloxidase from egg masses of the gastropod mollusc, _Biomphalaria glabrata_. Comp Biochem Physiol B118(2): 463-469.
-
Sritunyalucksana K, Söderhäll K (2000) The _pro_PO and clotting system in crustaceans. Aquaculture 191(1-3): 53-69.
-
Fang J, Han Q, Johnson JK, Christensen BM, Li J (2002) Functional expression and characterization of _Aedes_ _aegypti_ dopachrome conversion enzyme. Biochem Biophys Res Commun 290(1): 287-293.
-
Shelby KS, Adeyeye OA, Okot Kotber BM, Webb BA (2000) Parasitism-linked block of host plasma melanization. J Invertebr Pathol 75(3): 218-225.
-
Olivares C, Jimenez Cervantes C, Lozano JA, Solano F, Garcia Borron JC (2001) The 5,6-dihydroxyindole- 2-carboxylic acid (DHICA) oxidase activity of human tyrosinase. Biochem J 354(1): 131-139.
-
Hernández López J, Gollas Galván T, Góumez Jiméunez S, Portillo Clark G, Vargas Albores F (2003) In the spiny lobster (_Panulirus interruptus_) the prophenoloxidase is located in plasma not in haemocytes. Fish Shellfish Immunol 14(2): 105-114.
-
Jaenicke E, Decker H (2003) Tyrosinases from crustaceans form hexamers. Biochem J 371(2): 515-523.
-
Perdomo Morales R, Montero Alejo V, Perera E, Pardo Ruiz Z, Alonso Jiménez E (2007) Phenoloxidase activity in the hemolymph of the spiny lobster _Panulirus argus_. Fish Shellfish Immunol 23(6): 1187-1195.
-
Chen JS, Rolle RS, Marshall MR, Wei CI (1991) Comparison of phenoloxidase activity from Florida spiny lobster and western Australian lobster. J Food Sci 56(1): 154-157.
-
Liu G, Yang L, Fan T, Cong R, Tang Z, et al. (2006) Purification and characterization of phenoloxidase from crab _Charybdis japonica_. Fish Shellfish Immunol 20(1): 47-57.
-
Aladaileh S, Rodney P, Nair SV, Raftos DA (2007) Characterization of phenoloxidase activity in Sydney rock oysters (_Saccostrea glomerata_). Comp Biochem Physiol B 148(4): 470-480.
-
Shanthi S, Sivakumar MR, Rayvathy B (2021) Serum phenoloxidase activity in the hemolymph of the anomuran crab _Albunea symmysta_ (Linnaeus, 1758) (Decapoda: Anomura: Albuneidae). J Crustac Biol 41(1): ruab003.
-
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem 72: 248-254.
-
Perazzolo LM, Barracco MA (1997) The prophenoloxidase activating system of the shrimp _Penaeus paulensis_ and associated factors. Dev Comp Immunol 21(5): 385-395.
-
Acharya S, Mohanty J, Sahoo PK (2004) Humoral defense factors in Indian River prawn, _Macrobrachium_ _malcolmsonii_. Fish Shellfish Immunol 17(2): 137-147.
-
Fan TJ, Wang XF (2002) Purification and partial biochemical characterization of phenoloxidase from _Penaeus chinensis_. Acta Biochim Biophys Sin 34(5): 589- 594.
-
Gollas Galván T, Hernández López J, Vargas Albores F (1999) Prophenoloxidase from brown shrimp _Penaeus_ _californiensis_ hemocytes. Comp Biochem Physiol B 122(1): 77-82.
-
Simpson BK, Marshall MR, Otwell WS (1987) Phenoloxidase from shrimp (_Penaeus_ _setiferus_): purification and some properties. J Agric Food Chem 35(6): 918-921.
-
Rolle RS, Guisan N, Chen JS, Marshall MR, Yang JS, et al. (1991) Purification and characterization of phenoloxidase isoforms from Taiwanese black tiger shrimp (_Penaeus monodon_). J Food Biochem 15(1): 17- 32.
-
Laskin JD, Piccinini LA (1986) Tyrosinase isozyme heterogeneity in differentiating B16/ C3 melanoma. J Biol Chem 261(35): 16626-16635.
-
Brivio MF, Mazzei C, Scarf G (1996) proPO system of _Allogamus auricollis_ (Insecta): effects of various compounds on phenoloxidase activity. Comp Biochem Physiol B 113(2): 281-287.
-
Kahn V, Andrawis A (1985) Tropolone as a substrate for horseradish peroxidase. Phytochemistry 24(5): 909- 913.
-
Kahn V (1985) Tropolone-a compound that can aid in differentiating between tyrosinase and peroxidase. Phytochemistry 24(5): 915-920.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework