The Morphology and Description of Glaridacris catostomi Cooper, 1920 (Cestode: Caryophyllidae) from Catostomid Fishes in Lakes and Rivers of Wisconsin, with Morphological Observations
Thirteen species in 8 genera of caryophyllid cestodes including Glaridacris catostomi Cooper, 1920 were found in Catostomus commersoni (Lacépède), Cyprinus carpio (Raf.), and Erimyzon sucetta (Lacépède) in 2 lakes and 2 streams in SE Wisconsin and/or SW Lake Michigan. The morphology of the Wisconsin populations of G. catostomi is revisited using light microscopy revealing new features of the scolex and reproductive organs. The development of scolex and some reproductive structures were observed in maturing specimens. We describe new specimens and provide a comparative morphometric data with other earlier descriptions by Cooper, Hunter, and Mackiewicz. Our specimens were largest compared to those examined by other observers and many structures were correspondingly larger. The status of the genus Glaridacris was discussed in terms of most recent taxonomy and its evolutionary relationships.
Introduction
Mackiewicz OS [1] reviewed the morphology, anatomy, development, zoogeography, and systematics of the Caryophyllidea (Cestoidea) and indicated the existence of 89 species and 37 genera in these monozoic cestodes. Ten years later, Mackiewicz JS [2] provided perspectives on the evolution, biology and zoogeography of 128 species in 45 genera of caryophyllids from the Nearctic, Neotropical, Palearctic, Ethiopian, Oriental, and Australian regions. Twelve years later, Mackiewicz JS [3] recognized about 150 species in 41 genera worldwide (except for the Neotropical Region). Of these, 14 genera and 90 species belonging to three families have been described from the Indomalayan region from catfishes (Siluriformes: Bagridae, Clariidae, Heteropneustidae, Schilbeidae and Siluridae), cyprinid, and cobitid fishes. A large number of inadequately described species from clariid and heteropneustid catfishes in India and neighboring countries have been invalidated by Ash, et al. [4, 5]. Most recently, however, Scholz, et al. [6] and Scholz T, et al. [7] recognized only 117 species and 46 genera in order Caryophyllidea van Beneden in Carus, 1863 parasitizing teleost fish (Cypriniformes, Siluriformes, and some catfish). Chubb JC [8] reviewed the seasonal ecology of the Caryophyllidea in the various climate zone of the world.
The order consists of four families, namely the Balanotaeniidae, Capingentidae, Caryophyllaeidae and Lytocestidae; see Mackiewicz JS [3]. Classification at the family level is based on the position of the inner longitudinal musculature in relation to the testes and vitelline follicles; see Mackiewicz OS [1, 3]. “The caryophyllideans are exclusively intestinal parasites of freshwater teleosts with 85 species reported from cypriniform and 29 from siluriform fishes. Only few species of the Lytocestus with 2 species and monotypic Lytocestoides are known from mormyrids (Osteoglossiformes), characiform and cichlid (Perciforms) fishes, respectively. Within families, 40 % of caryophyllideans parasitize North American suckers (Catostomidae), 26 % cyprinids, 10 % clariid catfishes and just a few caryophyllideans have been reported from 12 additional fish families” [7]. Freshwater annelids of the family Naididae, especially species previously placed in the Tubificidae (Tubifex tubifex, Limnodrilus spp.), serve as the only intermediate hosts [1].
In Arizona and Wisconsin, Amin OM [9, 10, 11] recognized 14 species in 8 families of caryophyllid cestodes from suckers (Catostomidae) in lake and river systems. These included Glaridacris catostomi Cooper, 1920 from Catostomus commersoni (Lacépède) in SE Wisconsin lakes and streams. See collections, following, for details. Table 1 includes a list of the 14 species of cestodes collected and organized alphabetically by host species from Arizona (A) and Wisconsin (W) based on Amin OM [9, 11].
Glaridacris catostomi is widespread in North America; see Hoffman GL [12] for a list of records with a large number of definitive hosts. Mackiewicz JS [13] redescribed the species and added numerous host and locality records. In Wisconsin, Amin OM [14, 11] discussed G. catostomi infections in the anterior intestine of 231 C. commersoni from the Pike River in 1972-1974, 10 from Silver Lake, and 105 from Tichigan Lake in 1976 and 1984. The anterior localization of infections did not vary by season. Amin OM [15] (p. 213) reported 34 specimens of G. catostomi “from the anterior half of the intestine of 9 of 14 C. commersoni examined during January, March and April” in 1973-1974 from coastal SW Lake Michigan at the Wisconsin-Illinois state line near the outlet of the Pike River. The Lake Michigan infections appear to have resulted from the overflow of infected individuals of C. commersoni from the Pike River into the lake. Our Wisconsin specimens were “similar to those described by Cooper AR [16] and Mackiewicz JS [13] but exhibited some variations. Mackiewicz’s JS [13] redescription of G. catostomi was adapted from that of Hunter III GW [17], included more elaborate figures, and was justified as providing “more specific host and distributional data than presented in an earlier abstract” (p. 554). In Wisconsin, G. catostomi was also reported from various parts of the state by Anthony JD [18], Bangham RV [19] and Fischthal JH [20, 21, 22]. Mackiewicz JS [13] (p.556) provided exhaustive records from many parts of North America from additional hosts including Catostomus catostomus (Forster), Catostomus occidentalis Ayres, Catostomus marcrocheilus Girard, Erimyzon oblongus (Mitchill), and “unverified” records from Hypentellium nigricans (LeSueur), Moxostoma rubreques Hubbs, sucker, white-nosed sucker, long-nosed sucker, pickerel, quillbacks, buffaloes, log perch, and mud puppy. Records after 1966 included Moxostoma anisurum Rafinesque, Moxostoma erythrurum Rafinesque, and Esox lucius Linn. Based on Hoffmann GL [12]. Mitchum DL [23] (p.107) reported it from C. commersoni in “waters throughout Wyoming.” The only record of G. catostomi outside of North America is that of Bauer ON [24] from the Siberian sucker Catostomus catostomus rostratus (Tilesius) in the Kolyma River which flows into the Kolyma Gulf of the East Siberian Sea, a division of the Arctic Ocean. The transmission of G. catostomi from infected North American suckers to Siberia must have taken place before the crossing of American suckers to Asia “between Early and Middle Pleistocene when prolonged reproductive isolation led to interspecies divergence with the appearance of amino acid substitutions… fixed due to positive selection” [25]. The introduction of G. catostomi to northeastern Siberian rivers, such as Kolyma and Indigirka, may have, however, occurred as a result of human introduction of C. catostomus to Russia [26].
“Molecular phylogenetic studies confirm monophyly of caryophyllideans, but also reveal the non-monophyly of three of the four families (the family Balanotaeniidae is mono-generic)” [27, 28, 29]. In contrast, most genera seem to be monophyletic, with a few exceptions such as Glaridacris Cooper, 1920” [6, 7].
Collections
In SE Wisconsin, 139 specimens of G. catostomi were collected from 5 of 10 examined white suckers, C. commersoni (mean 13.9) in the land-locked eutrophic natural Silver Lake (Kenosha County) (42°32′58″N,88°10′9″W) in April, 1977, and 141 specimens collected from 8 of 105 individuals of C. commersoni (mean 1.34) in Tichigan Lake, a relatively large eutrophic impoundment on the Fox River which is a tributary of the Mississippi River in Racine County (42°48′20″N,88°13′00″W) in April, 1976, April and November, 1977 and April, 1984 [11]. Over 100 specimens of G. catostomi were also recovered from C. commersoni in the Pike River in Racine and Kenosha counties (42°38′40″N,87°51′09″W) which flows in Lake Michigan, in May, 1973, April, 1983 [15] and miscellaneous. At least one white sucker from the nearby Root River was found infected with at least one specimen of G. catostomi in the autumn of 1974 [15]. The larger Root River in Milwaukee and Racine counties (42° 44’ 1” N-87° 46’ 42” W) and the Pike River flow eastwards into Lake Michigan. The poorer parasitic fauna in the Root River is attributed to its higher flow rate and its higher non-fecal organic pollutant content than in the Pike River [15]. Both rivers are independent streams not connected to either Silver or Tichigan lakes. Twenty specimens from 100 newly examined specimens from Silver and Tichigan lakes were used for microscopy.
Methods
Worms were initially stored in cold water then fixed in cold 70% ethanol. They were subsequently punctured with a fine needle then stained in Semichon’s carmine, destained in 4% hydrochloric acid in 70% ethanol, dehydrated in ascending concentrations of ethanol and cleared in clove oil or oil of winter green. Whole worms were mounted in Canada balsam. Measurements are in micrometers or mm as noted; the range is followed by the mean values between parentheses. Width measurements represent maximum width. Microscope images were created using 10X and 40X objective lenses of a BH2 light Olympus microscope (Olympus Optical Co., Osachi-shibamiya, Okaya, Nagano, Japan) attached to an AmScope 1000 video camera (United Scope LLC, dba AmScope, Irvine, California), linked to an ASUS lab top equipped with HDMI high-definition multimedia interface system (Taiwan-USA, Fremont, California). Images from the microscope are transferred from the lab top to a USB and stored for subsequent processing on a computer.
Results
Amin OM [11] briefly described specimens from C. commersoni in Silver and Tichigan lakes, SE Wisconsin and noted that his specimens were similar to those described by Cooper [16] and Mackiewwicz [13], with some variations including: “Cooper [16] reported fewer (150-160) and smaller testes (100-145 by 127-181) and shorter ovarian arms (800-900), and Mackiewicz (1965) reported shorter ovarian arms (700-900) and larger eggs (55-77 by 50-57).” Amin OM [14] initially reported over 100 specimens of G. catostomi from C. commersoni in the spring of 1973 from SE Wisconsin’s Pike River.
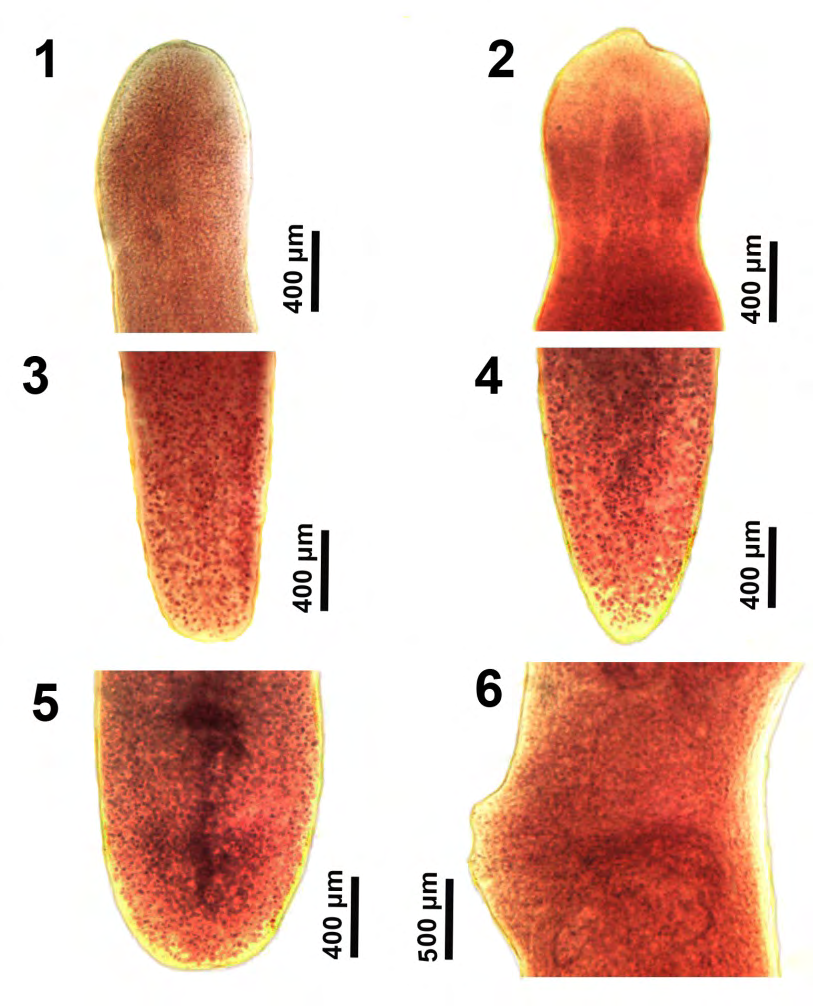
Figures 1-6: The development of anterior and posterior portions of larval and maturing specimens of Glaridacris catostomi collected from Catostomus commersoni in Silver and Tichigan lakes, SE Wisconsin. 1, 2. Early scolex. Note the external and internal muscles in Fig. 2. 3-5. Corresponding development in the posterior part of the body showing the beginning of formation of the incipient reproductive organs in Fig. 5. 6. Lateral view of the emerging cirrus sac (anterior) and coils of the pre-ovarian uterus (posterior) above and below the projecting genital swelling.
Amin OM [11] reported 60 juvenile, 103 adult, and 117 gravid specimens from 13 of 87 examined white suckers from Silver and Tichigan lakes in the autumn and spring between 1976 and 1984, but not in 28 suckers examined in the summer. Suckers from the land-locked Silver Lake were considerably more heavily and frequently infected than those from Tichigan Lake. We measured 20 additional mostly gravid adults and combined their measurements with those of measurements of the 10 specimens already measured by Amin OM [11] (p. 53) and combined the measurements (our recent Table 2). We also included microscope images of various structures including scolexes and reproductive structures (Figures 1-18). We provide below a brief revised account of our qualitative morphological findings with emphasis on first reported variations in the developing scolex and some reproductive structures (Figure 1-18) and a comparative morphometrical data for mature adults (Table 1) emphasizing variations and additions to our earlier account [11] including new measurements of 20 additional specimens, and those of Cooper AR [16], Hunter III GW [17], and Mackiewicz JS [13]. Cooper’s AR [16] description was comprehensive but Hunter III GW [17] (p. 51) considered it “at fault in several instances” even though he quoted most of Cooper’s description verbatim, used much of his measurements, and claimed his (Cooper’s) “ignorance” of an occasional inaccurate oversight. Hunter’s III [17] (pp. 51, 59) description was based on Cooper’s vials and slides loaned to him by Henry B. Ward as well as on other specimens collected by H. E. Essex from Burntside Lake, Minnesota, and by Hunter from Lake Erie, Silver Creek, New York, respectively, and on additional specimens loaned by George R. La Rue from C. commersoni in Douglas Lake totaling 75-80 specimens. Mackiewicz’s JS [13] (p. 554) specific diagnosis was “modified after Hunter [17] and based on (his) measurements…”; he provided only 5 measurements (see Table 2) but 14 informative figures.
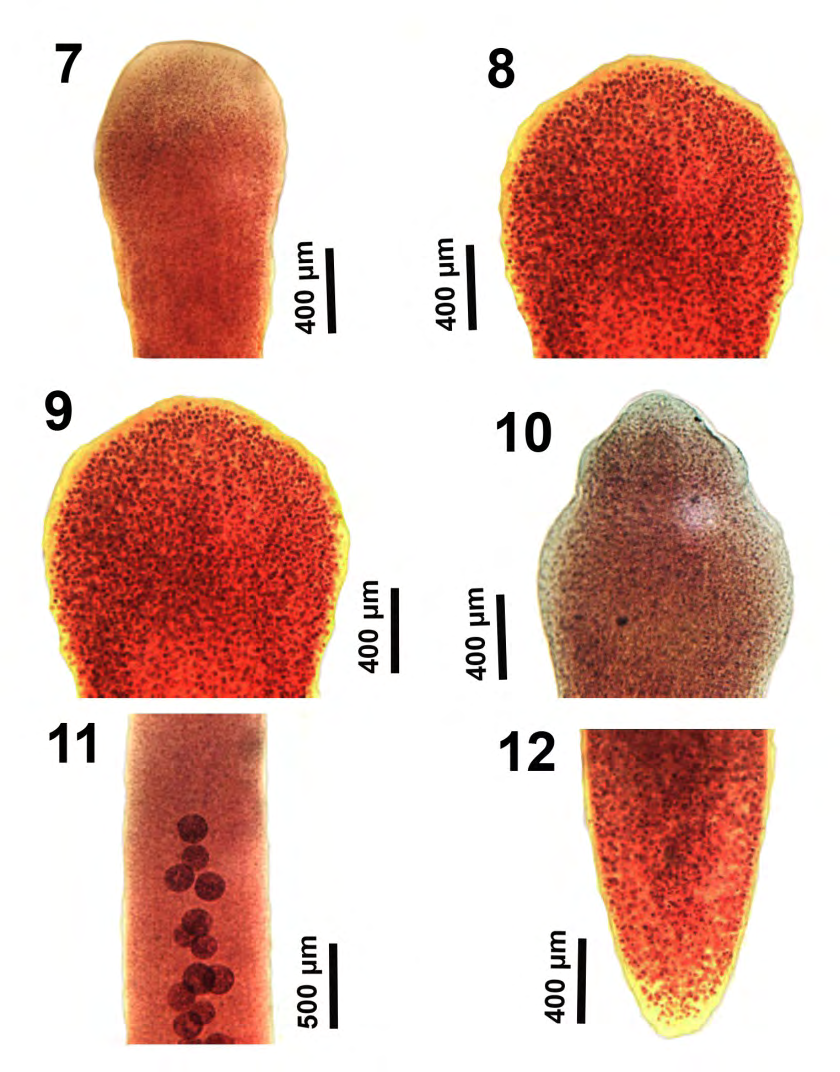
Figures 7-12: The development of anterior and posterior portions of larval and maturing specimens of Glaridacris catostomi collected from Catostomus commersoni in Silver and Tichigan lakes, SE Wisconsin. 7-10. Further development of the scolex. 11. A juvenile showing the early development of testes jest below the neck in the absence of vitellaria. 12. The posterior portion of the same worm in Figure 11 showing the corresponding stage of development of reproductive organs.
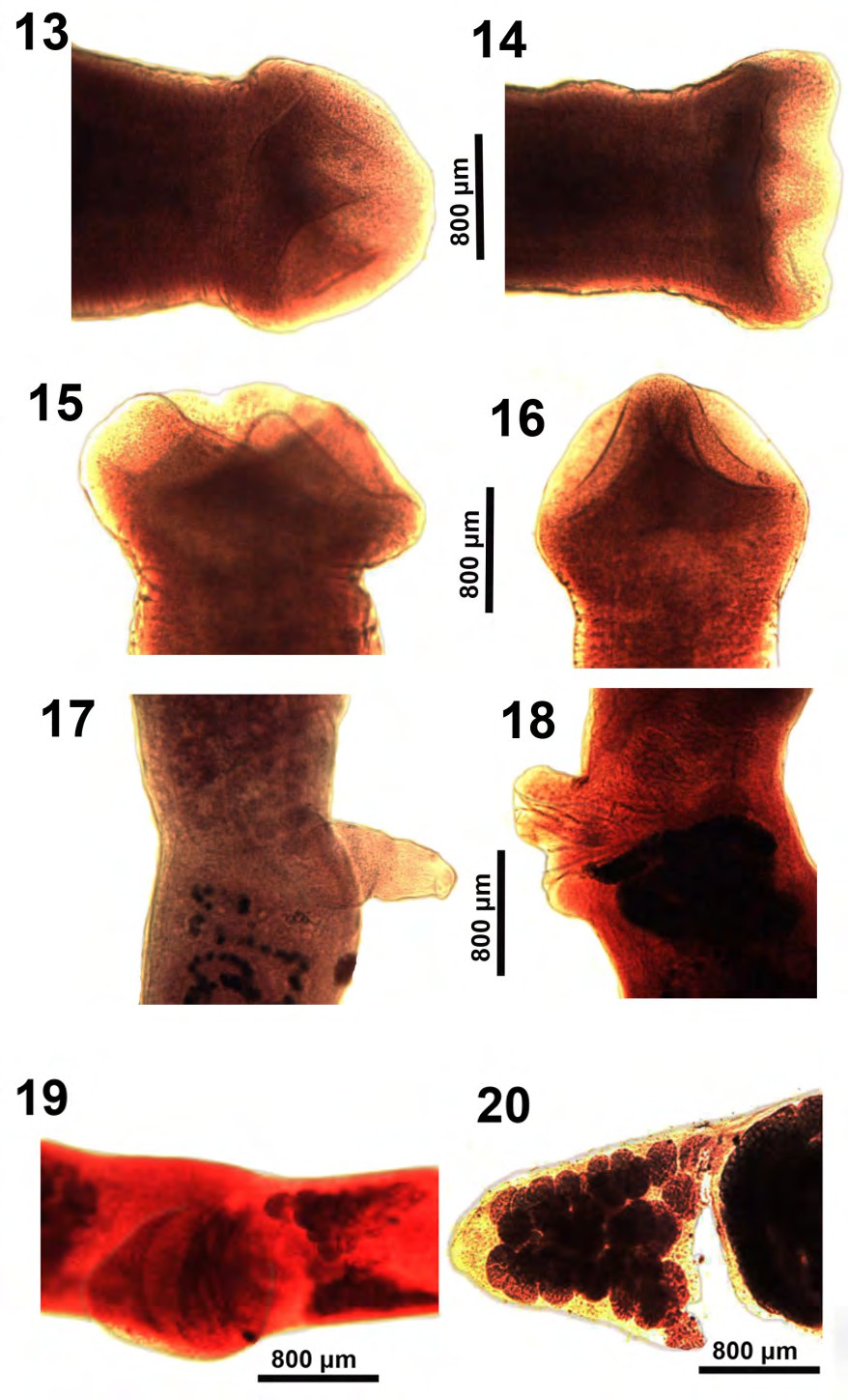
Figures 13-18: The anterior and posterior portions of gravid adults of Glaridacris catostomi collected from Catostomus commersoni in Silver and Tichigan lakes, SE Wisconsin. 13-16. Different perspectives of fully developed scolexes. 17. A lateral view of a worm with fully evaginated cirrus sac appearing like an erect penis and may be functioning as such. 18. A rare view of the separate male and female reproductive terminalia in a state of prolapsed evagination. The dark coils of the pre-ovarian uterus gravid with eggs feed into the more posterior genital opening.
Figures 19,20: Some reproductive structures of gravid adults of Glaridacris catostomi collected from Catostomus commersoni in Silver and Tichigan lakes, SE Wisconsin. 19. The follicular H-shaped ovary and expanded cirrus sac of one gravid adult. 20. Post-ovarian vitellaria at the posterior tip of another gravid specimen artificially broken off a post-ovarian uterine loop packed with eggs (right).
Considerably smaller immature specimens and younger unreproductive adults with undefined scolex were more common especially in younger fish. Worms were more prevalent, more mature, and larger in larger hosts [11]. Observations of new specimens show that testes develop first then vitelline glands and other reproductive structures appear next.
We provide below a brief revised account of our qualitative morphological findings and a comparative morphometrical data based on 30 specimens from C. commersoni in Silver and Tichigan lakes, Wisconsin (Table 2) highlighting variations from the other accounts of Cooper AR [16], Hunter III GW [17] and Mackiewicz JS [13].
Morphological Description of Adults
Body of medium length, dorso-ventrally flattened, covered with thin cuticle and sub-cuticle enclosing a layer of outer longitudinal muscles delimited by a basement membrane surrounding inner longitudinal muscles and medullary tissue enclosing, reproductive, nervous, and excretory systems. See table 1 for measurements and Figures 1-20 for microscopic images. Anterior scolex conical anteriorly developing into cuneiform chisel-shaped in older specimens, rounded posteriorly (Figures 1,2,7-10), divided in 2 dorsal and ventral lobes each with 3 loculi divided by 2 variably distinct ribs displaying a wide range of diversity (Figure 13-16). Neck, narrowest part of body, not especially differentiated. Vitelline glands in 2 regions; the first beginning just posterior to neck anteriorly and extending in 2 lateral bands, past cirrus sac extending to mid-anterior uterus, and the second in a separate cluster posterior to ovary (post ovarian vitellaria) (Figure 20). Anterior vitellaria partially encircling testes. Oblong-ellipsoidal testes develop first in medullary parenchyma occupy central space (Figure 11) shorter than that occupied by vitellaria posteriorly and extending only to cirrus sac. Coils of the vas deference at mid-body of juveniles (Figure 6) dilate posteriorly into a muscular ejaculatory duct which narrows ventrally into large spherical muscular cirrus sac (Figure 17-19). Cirrus sac independent of uterovaginal canal which opens separately more posteriorly (Figure 18). Lobate ovary shaped like letter-H (Figure 19) often beginning just posterior to level of anterior loops of uterus. Convoluted thin-walled uterus continues from anterior to posterior ovary passing between long arms of ovary emptying into female atrium next to the vagina posterior to cirrus sac (Figure 18). Posterior end bluntly rounded where excretory bladder drains into excretory pore (Figures 3-5,12,20). Operculate eggs with smooth surface.
Deposited Specimens: USNM Helm. Coll. Nos. 68657, 78811; Milwaukee Publ. Mus. Coll. No. IZ985-01C.
Remarks
Our microscope images document the outline and the inner perspective of the scolex and some reproductive structures that are hardly perceptible in the excellent line drawings by Mackiewicz [13] (Figs. 1-14), and by Oros et al. [26] (Figures 1A,2A&2B) or in the SEM images of the scolex by Oros, et al. [26] (Figure 4A). The development of the shape, depth, and loculi of the scolex from the shallow undefined immature form (Figures 1,2,7&8) to the more demarcated form of juveniles (Figures 910), leading to the more defined chisel-cuneiform adult form with well-defined loculi (Figures 13-16) are well demonstrated. Corresponding changes in the developing posterior end of worms are shown in figures 3-5, 12. Inner and outer longitudinal muscles begin to appear in immatures (Figure 2). The early development of the testes begins in young adults (Figure 11), and so does the early development of the cirrus sac (Figure 6). The everted cirrus sac and the two separate genital terminalia are well documented (Figures 17&18). The shape of the follicular ovarian lobes and that of a fully enlarged cirrus sac (Figure 19) are well presented. The posterior end of a sexually mature adult shows an image of the post-ovarian vitellaria artificially separated from the posterior-most loop of the egg-loaded post-ovarian uterus (Figure 20).
Our specimens were morphometrically compared with those of Cooper AR [16] and Hunter III GW [17] who quoted much of Cooper’s text and measurements. Mackiewicz’s JS [13] redescription missed many measurements (Table 1). Our specimens from Wisconsin were the largest reaching over 60 mm in total length compared to a maximum of 41.0 mm in New York [13] and of 25.0 mm in Michigan, Minnesota, and New York [16]; Hunter III GW [17]. Body width measurements at gonopore, diameter of testes, vitelline glands, cirrus sac, ovarian arm, and eggs reached correspondingly larger dimensions in our Wisconsin specimens. Mackiewicz JS [13] counted more testes reaching 463 and Cooper AR [16] counted least “150-160”. Shortest distance between anterior-most testis to anterior worm tip was shorter (0.82 mm) than that between anterior-most vitelline gland and anterior worm tip (1.60) in our Wisconsin specimens (Table 2). The Wisconsin specimens had the largest number of post-ovarian vitellaria 17-56 (35). The number of post-ovarian vitellaria in other collections was deduced from published figures and probably represent an underestimate. Other than size differences, our microscope images provided a vestige of the development of structures like the scolex from larval to fully developed adult stage. The total range of variability presented from the same host species is considered a measure of intraspecific variability and gives no reason for assigning different types or varieties.
| Parasite species | State | |
|---|---|---|
| Carpoides Cyprinus (LeSueur); quillback | Rowardleus pennensis Mackiewicz and Deutsch, 1976 | W |
| Catostomus clarkii Baird and Girard; desert sucker | Isoglaridacris hexacotyle (Linton, 1897) Mackiewicz, 1968 | A |
| Catostomus commersoni (Lacépède); white sucker | Biacetabulum biloculoides Mackiewicz and McCrae, 1965 | W |
| Catostomus commersoni (Lacépède); white sucker | Biacetabulum macrocephalum MacCrae, 1962 | W |
| Catostomus commersoni (Lacépède); white sucker | Biacetabulum sp. Hunter, 1927 | W |
| Catostomus commersoni (Lacépède); white sucker | Glaridacris catostomi cooper, 1920 | W |
| Catostomus commersoni (Lacépède); white sucker | Glaridacris laruei (Lamont, 1921) Hunter, 1927 | W |
| Catostomus commersoni (Lacépède); white sucker | Hunterrella nodulosa Mackiewicz and McCrae, 1962 | W |
| Catostomus commersoni (Lacépède); white sucker | Isoglaridacris folius Fredrickson and Ulmer, 1967 | W |
| Catostomus commersoni (Lacépède); white sucker | Monobothium hunteri Mackiewicz, 1963 | W |
| Catostomus insignis Baird and Girard; Sonora sucker | Isoglaridacris hexacotyle (Linton, 1897) Mackiewicz, 1968 | A |
| Cyprinus carpio Linn.; common carp | Atractolytocestus huronensis Anthony, 1958 | W |
| Cyprinus carpio Linn.; common carp | Khawia iowensis Calentine and Ulmer, 1961 (W) | W |
| Erimyzon sucetta (Lacépède); lake chubsucker | Glaridacris laruei (Lamont, 1921) Hunter, 1927 | W |
| Erimyzon sucetta (Lacépède); lake chubsucker | Isoglaridacris agminis Williams and Rogers, 1972 | W |
| Erimyzon sucetta (Lacépède); lake chubsucker | Isoglaridacris multivitellaria Amin, 1986 | W |
Table 1: Caryophyllid cestodes collected from fish in Arizona (A) and Wisconsin (W) as reported in Amin (1969, 1986).
| Amin (1986) & this paper | Cooper (1920) | Hunter (1930) | Mackiewicz (1965) | |
|---|---|---|---|---|
| Silver & Tichigan lakes, Wisconsin | Douglas Lake (DL), Michigan | DL, Michigan Minnesota, N.Y. | Central New York (N.Y.) | |
| N | 10 & 20 | 15 | 75-80 | 29 |
| Length (L) (mm) | 16.80-41.21 (25.60)* worms >60 mm in ethanol | 5.0-25.0 | 5.0-25.0** | 11.5-41.0 |
| Width at gonopore (mm) | 0.90-1.87 (1.41) | 1.0 | 0.4-1.0 | --- |
| Scolex length (mm) | 0.75-1.12 (0.85) | 0.30-0.45 | 0.30-0.45 | --- |
| Scolex width (mm) | 0.68-1.62 (1.16) | 0.45-1.10 | 0.45-1.10 | --- |
| Testes (T) number | 160-401 (292) | 150-160 | 405-420 | 171-463 |
| Testes diameter μm | 77-500 (254) | 100-227 | 120-190 | --- |
| Vitelline glands diameter (μm) | 50-250 (174) | Up to 200 | Up to 200 | --- |
| First T to anterior tip (mm) | 0.82-3.25 (2.28) | --- | --- | --- |
| First pre-ovarian vitellarium to anterior tip (mm) | 1.60-3.00 (2.17) | 1.50-2.50 | --- | --- |
| % of worm length | 6-13 (9) | --- | --- | --- |
| Cirrus sac diameter (μm) | 350-750 (605) | 400-600 | 400-600 | |
| Post-gonopore distance (mm) | 2.20-5.88 (3.55) | --- | --- | --- |
| % of worm length | 112-17 (14) | --- | --- | --- |
| No. post-ovarian vitellaria | 17-56 (35) | 10 (Fig. 6) | 15 (Fig. 25) | 25 (Fig. 3) |
| Ovarian arms length (mm) | 0.69-2.00 (1.32) | 0.8-0.9 | 0.65-0.90 | 0.70-0.90 |
| Egg length (μm) | 42-78 (56) | 54-66 | 54-66 | 57-77 |
| Egg diameter (μm) | 35-51 (40) | 38-48 | 38-48 | 50-57 |
Table 2: ** Comparative measurements of mature _Glaridacris catostomi_ from _Catostomus commersoni_ collected in Wisconsin, centr
*Range (mean). Hunter (1930) used bolded figures from Cooper (1920). Table 2:** Comparative measurements of mature Glaridacris catostomi from Catostomus commersoni collected in Wisconsin, central New York, and Michigan.
Discussion
“It appears that the Nearctic and Palearctic Caryophylleaidae are distinct from each other and therefore the presence of G. catostomi in Catostomus catostomus, the only catostomid common to both of these two regions, assumes zoogeographical importance…. Glaridacris catostomi may have been possibly introduced into the Palearctic through Catostomus [13] (p. 558). Waeschenbach A, et al. [30] proposed that caryophyllideans underwent radiations not only within host groups but also within zoogeographical regions: in Afro-Indian catfishes, in Palaearctic cypriniforms, and in Nearctic catostomids. With high host specificity, it is perhaps not surprising that the evolutionary radiation of caryophyllideans is also “strongly influenced by biogeography” [6]. Scholtz et al. [6] made no definitive statements about the geographical origin of the group but indicated (p. 3) that “Biogeographic patterns are indicative of separate Gondwanan and Laurasian radiation.” Mackiewicz JS [3] noted the large diversity of characters within the genus Glaridacris, among 4 Holarctic genera, that he designated as a complex reflecting their systematic uncertainty and whether they represent complexes of multiple genera or possibly subgenera. Oros et al. [26] recognized the artificial character of the genus Glaridacris and split it into 2 different, morphologically distinct genera. (A) Glaridacris now includes only 3 species, G. catostomi (type species), Glaridacris terebrans (Linton, 1893), and Glaridacris vogei Mackiewicz, 1976, which are characterized by an elongate body, a cuneiform or wedge-shaped scolex with 6 shallow loculi, distant male and female gonopores, follicular ovary, and lateral and median vitellaria. (B) A new genus Pseudoglaridacris, now accommodates 3 other species: P. laruei (Lamont, 1921) Oros, Uhrovič, Scholz, 2018 (type species), P. confusa (Hunter, 1929) Oros, Uhrovič, Scholz, 2018, and P. oligorchis (Haderlie, 1953) Oros, Uhrovič, Scholz, 2018, with a shorter body, a bothrioloculate scolex with a pair of deeper median bothria and 2 shallower loculi, close male and female gonopores, non-follicular ovary, and with only lateral vitellaria., p. 60 [26]. The genus Glaridacris remains exclusively a Nearctic genus comprised of species parasitizing suckers [3, 26].
We document for the first time some of the characteristic morphological features of G. catostomi using microscopic images that were not reported by other observers that have used line drawings. The developing scolex form in immature, juvenile and mature adult form is particularly noteworthy. Images of the independent female and male genital orifices were particularly demonstrative. Comparative sizes of the body and other structures between our larger specimens from Wisconsin and those measured by Cooper AR [16], Hunter III GW [17] and Mackiewicz JS [13] revealed morphological variabilities attributable to geographical locations in addition to intraspecific variabilities.
• Acknowledgment This project was supported by an Institutional Grant from the Parasitology Center, Inc. (PCI), Scottsdale, Arizona.
Declarations
Compliance with Ethical Standards
- Conflict of Interest: The authors declare no conflicts of interest or competing interests.
- Ethical Approval: The authors declare that they have observed all applicable ethical standards.
- Availability of Data: All presented and related data are available by contacting the senior author.
References
-
Mackiewicz ОS (1972) Caryophyllidea (Cestoidea): a review. Exper Parasitol 31(3): 417-512.
-
Mackiewicz JS (1982) Caryophyllidea (Cestoidea): Perspect Parasitol. 84: 397-417.
-
Mackiewicz JS (1994) Order Caryophyllidea van Beneden in Carus, 1863. In: Khalil LF, Jones A, Bray RA, et al. (Eds.), Keys to the cestode parasites of vertebrates. CAB International, Wallingford, UK, pp: 21-43.
-
Ash A, Scholz T, Oros M, Kar PK (2011a) Tapeworms (Cestoda: Caryophyllidea), parasites of _Clarias batrachus_ (Pisces: Siluriformes) in the Indomalayan region. J Parasitol 97: 435-459.
-
Ash A, Scholz T, Oros M, Levron C, Kar PK (2011b) Cestodes (Caryophyllidea) of the stinging catfish _Heteropneustes fossilis_ (Siluriformes: Heteropneustidae) from Asia. J Parasitol 97(5): 899-907.
-
Scholz T, Waeschenbach A, Oros M, Brabec J, Littlewood DTJ (2021) Phylogenetic reconstruction of early diverging tapeworms (Cestoda: Caryophyllidea) reveals ancient radiations in vertebrate hosts and biogeographic regions, Int J Parasitol 51(4): 263-277.
-
Scholz T, Kuchta R (2017) A digest of fish tapeworms. Vie et milieu - Life and environment 67(2): 43-58.
-
Chubb JC (1982) Seasonal occurrence of helminths in freshwater fishes. Part IV. Adult Cestoda, Nematoda and Acanthocephala. Adv Parasitol 20: 1-292.
-
Amin OM (1969) Helminth fauna of suckers (Catostomidae) of the Gila River System, Arizona II Five parasites from _Catostomus_ spp. Am Midland Nat 82(2): 429-443.
-
Amin OM (1974) Intestinal helminthes of the white sucker, _Catostomus commersoni_ (Lacépède), in SE Wisconsin Proc Helminthol Soc Wash 41: 81-88.
-
Amin OM (1986) Caryophyllaiedae (Cestoda) from lake fishes in Wisconsin with a description of _Isoglaridacris multivitellaria_ sp. n. from _Erimyzon sucetta_ (Catostomidae). Proc Helminthol Soc Wash 53: 48-58.
-
Hoffman GL (1999) Parasite of North American freshwater fishes. Comstock Publishing Assoc, Cornell Univ Press, Ithaca, New York, USA, pp: 539.
-
Mackiewicz JS (1965) Redescription and distribution of _Glaridacris catostomi_ Cooper, 1920 (Cestoidea: Caryophyllaeidae) J Parasitol 51: 554-560.
-
Amin OM (1975) Intestinal helminthes of some southeastern Wisconsin fishes. Proc Helminthol Soc Wash 42: 43-46.
-
Amin OM (1977) Distribution of fish parasites from two southeast Wisconsin streams. Trans Wis Acad Sci, Arts, Lett 65: 225-230.
-
Cooper AR (1920) _Glaridacris catostomi_ n g, n sp, a cestodarian parasite. Trans Am Microscop Soc 39: 5-24.
-
Hunter III GW (1930) Studies on the Caryophyllaeidae of North America. Ill Biol Monogr 11(1927). pp: 186.
-
Anthony JD (1963) Parasites of eastern Wisconsin fishes. Tr Wisc Acad Sci, Arts Let 52: 83-95.
-
Bangham RV (1946) Parasites of Northern Wisconsin fish. Tr Wisc Acad Sci, Arts Let 36: 291-325.
-
Fischthal JH (1947) Parasites of Northwest Wisconsin fish I. The 1944 survey. Tr Wisc Acad Sci, Arts Let 37: 157-220.
-
Fischthal JH (1950) Parasites of Northwest Wisconsin fish. II. The 1945 survey. Tr Wisc Acad Sci, Arts Let 40: 87-113.
-
Fischthal JH (1952) Parasites of Northwest Wisconsin fish. III. The 1946 survey. Tr Wisc Acad Sci, Arts Let 41: 17-58.
-
Mitchum DL (1995) Parasites of fishes in Wyoming. 1995. Wyom Game and Fish Dept Cheyenne pp: 304.
-
Bauer ON (1987) Key to the parasites of freshwater fish of the USSR, Volume 3, Parasitic Metazoa (part 2) Lenigrad, Nauka pp: 583.
-
Bachevskaya LT, Pereverzeva VV, Ivanova GD, Agapova GA, Primak AA (2014) Genetic structure of the Siberian Sucker (Catostomus catostomus rostratus) according to data on sequence variation of the mtDNA cytochrome b gene. Biol Bull Russ Acad Sci 41: 306-311.
-
Oros M, Uhrovič D, Scholz T (2018) A new classification of Glaridacris Cooper, 1920 (Cestoda: Caryophyllidea), parasites of suckers (Catostomidae) in North America, including erection of Pseudoglaridacris n gen. J Parasitol 104(1): 60-69.
-
Olson PD, Scholz T, Poddubnaya LG, Littlewood DTJ (2008) On the derived position of _Archigetes_ and the early evolution of the tapeworms (Platyhelminthes: Cestoda). J Parasitol 94: 898-904.
-
Brabec J, Scholz T, Králová-Hromadová I, Baszalovicsová E, Olson PD (2012) Substitution saturation and nuclear paralogs of commonly employed phylogenetic markers in the Caryophyllidea, an unusual group of non- segmented tapeworms (Platyhelminthes). Int J Parasitol 42: 259-267.
-
Scholz T, Oros M (2017) Caryophyllidea. In Caira JN, Jensen K eds, Planetary Biodiversity Inventory (2008- 2017): Tapeworms from Vertebrate Bowels of the Earth. University of Kansas, Natural History Museum, Special Publication No 25, Lawrence, Kansas, pp: 47-64.
-
Waeschenbach A, Littlewood DTJ (2017) A molecular framework for the Cestoda. In: Caira JN, Jensen K, et al. (Eds.), Planetary Biodiversity Inventory (2008-2016): Tapeworms from Vertebrate Bowels of the Earth. The University of Kansas Natural History Museum pp: 431- 451.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework