Bee Diversity Associated with Canavalia rosea (Sw.) DC. (Fabaceae, Papilionoideae) in an Peri-Urban Patch of “Restinga”
The Restinga is one of the physiognomies of the Atlantic Forest biome with the highest degree of anthropic impact, mainly due to disordered urbanization. The impacts generate habitat loss and threaten the animals and plants that inhabit this ecosystem. In the Restinga, the psammophilous vegetation have an important role in stabilizing the sandy substrate and maintaining the pollinator community. In this study, we analyzed the floral resources offered by Canavalia rosea to floral visitors between October 2019 and September 2020, in the Municipal Natural Park of Barreto’s Restinga, in Macaé, Rio de Janeiro, Brazil. Sampling was carried out from 8:00 am to 4:00 pm, covering a 250-meter transect in the sandy cord, counting the total number of open flowers and collecting visiting bees. The morphology of C. rosea flowers was described to relate to the functional traits of visitors, related to pollen and nectar collection. The pollen load of visitors was analyzed to estimate the degree of specialization in relation to C. rosea. A total of 192 individuals of 14 bee species were captured. There was a significant relationship between the availability of floral resources, measured by the number of open flowers and the richness and abundance of bees. The greatest richness of bees was observed between January and March and the greatest abundance in January. The most frequent visiting genera in C. rosea were Eufriesea, Centris and Xylocopa. The greatest richness of pollen types was observed in Eufriesea species. Most Xylocopa individuals showed C. rosea pollen. In this study, the importance of C. rosea as a source of floral resources for fourteen species of solitary bees was demonstrated. The data generated increases the knowledge about the ecological patterns of C. rosea and its floral visitors and may contribute to the maintenance and expansion of Restinga areas in urban environments.
Introduction
The Atlantic Forest is a diverse biome, with a high rate of endemism and formed by different physiognomies [1, 2]. Among them, the Restinga, a geologically recent environment, formed by processes of sandy sediment deposition during the Quaternary period [3, 4]. Restinga areas, as they are adjacent to the coast, harbor vegetation constantly exposed to salinity, light and high temperatures, in addition to the scarcity of nutrients and water in the soil, mobility of the dunes and the action of the winds [5].
The psammophilous vegetation in the Restinga occupies the sand area between the upper limit of high tide and the dunes, mitigating the action of erosive agents [6] and protecting the substrate from the action of the winds [4]. The genus Canavalia, one of the groups that compose the psammophilous vegetation, has a wide distribution on the coast of Brazil, with the species Canavalia rosea being one of the most abundant.
Individuals of C. rosea grow in a creeping way and form long mats along the sand cord of the beaches [7, 8]. The long stolons, roots that attach to the substrate, and rapid growth contribute to soil stabilization, preventing erosion and reducing the direct impact of wind and wave action [9]. In addition, the flowers of C. rosea provide food resources such as pollen and nectar for several species of bees, mainly Bombus, Epicharis, Centris and Xylocopa, which are able to forage even under the strong winds [10, 11, 12].
Despite the importance of the ecosystem service of bees as pollinators of several plant species, several studies have reported worldwide declines in bee populations, the main causes being contamination by pesticides, fungi and habitats loss [13, 14, 15, 16, 17, 18, 19]. Habitat loss reduces the diversity and abundance of pollinating species [16], making resources such as nesting sites, food and other physical conditions scarce [20]. Estimates indicate that half of the Earth’s surface has been altered by human actions, mainly agriculture and urbanization [21], causing loss of natural habitats for several species. Thus, natural habitats in urban environments are considered refuges [22] that can shelter part of the biodiversity, including communities of pollinators such as bees, butterflies and birds [23, 24, 25].
On the Brazilian Atlantic coast, Restingas are mainly threatened by tourism and disorderly human occupation [26, 27]. Works aimed at surveying the fauna and flora in these environments are important for understanding the communities that inhabit the Restingas and can support conservation actions or management plans. Very little is known about the invertebrate species that live in the Restingas, and even less about the interactions in which they participate [28]. In this sense, this work is the first developed in the newly created Municipal Natural Park of Barreto’s Restinga, the second park in Brazil in extension of Restinga in urban area. The general objective was to survey the floral visitors of C. rosea, to answer the following questions: (1) do the plants provide floral resources throughout the year? (2) does the richness of visiting bees vary throughout the year? and (3) how is the foraging behavior of bee species visiting C. rosea? Are these bees generalists or specialists in resource collection?.
Material and Methods
Study Area
The study was carried out in the Municipal Natural Park of Barreto’s Restinga (22º20’22”S and 41º44’28”W) located in the municipality of Macaé, State of Rio de Janeiro (Figure 1). The park has an area of approximately 32 hectares, composed of shrubby vegetation and a sandy cordon with psammophilous vegetation. The climate of the region is tropical, classified as Aw according to [29]. The average temperature is 22.9°C and the average annual precipitation is 1126 mm, with the highest concentration of rainfall during the summer [30].
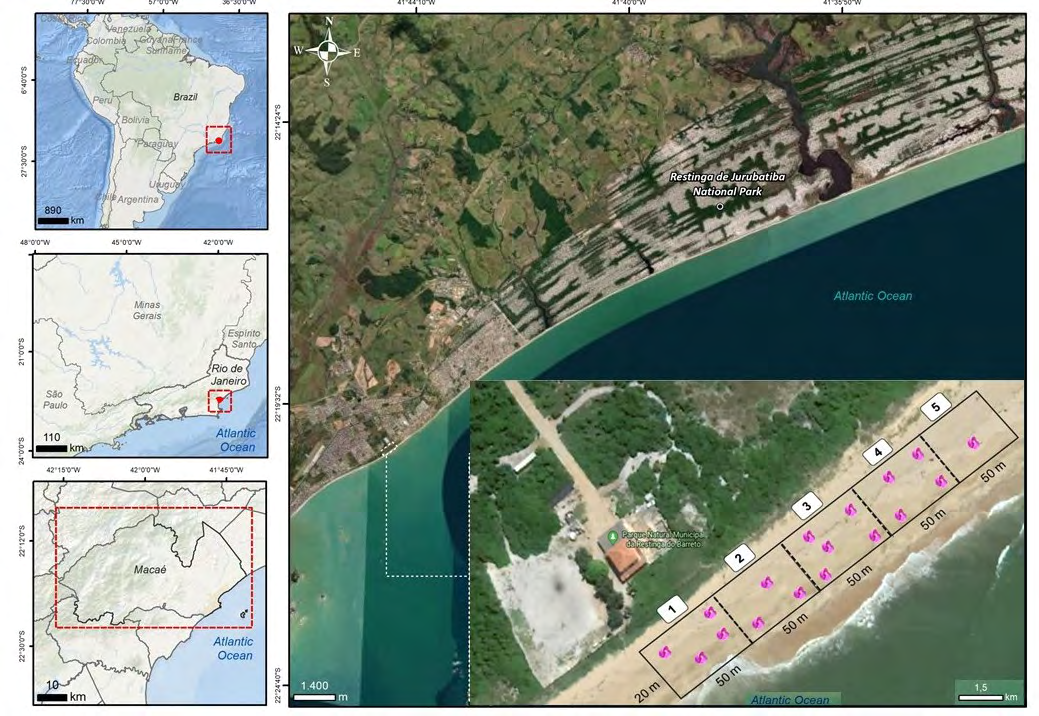
Figure 1: Geographic map of the study area showing the municipality of Macaé on the coast of Rio de Janeiro State, Brazil. The Municipal Natural Park of Barreto’s Restinga (PNMRB) is separated from the Jurubatiba’s Restinga National Park by an urban area. The white dashed indicates the sampling area and the plots within the 250 m transect in the sandy cord.
Sampling
During one year, the study area was inspected fortnightly to assess whether C. rosea flowered. In the months with flowering, the collections were carried out monthly (from October 2019 to September 2020), between 08:00 am to 16:00 pm, along a 250 m transect divided into 5 plots (50 x 20 m) for better results in the flower count.
During the sampling, the total number of open flowers of C. rosea were quantified. Flowering duration was categorized according to [31] in short (< 1 month), intermediate (between 1 and 5 months) or long (> 5 consecutive months).
Floral visitors were actively collected with entomological nets by two collectors, who covered the entire transect in the morning (8:00 to 12:00 h) and in the afternoon (13:00 to 16:00 h). The collectors remained for about 50 minutes in each sample plot, observing the behavior of the visitors in the flowers of C. rosea to describe the dynamics of insect- plant interaction. Floral visitors were classified as potential pollinators when their body size and behavior allowed the floral reproductive structures to come into contact with some part of the body, during visits to collect nectar or pollen, and as robbers when visitors did not contact the reproductive parts of the flower.
Captured bees were sacrificed in a killing chamber (ethyl acetate), mounted on entomological pins and identified to the highest possible taxonomic level based on the keys of [58] and [59]. Each individual was analyzed under an optical stereomicroscope to find the location of pollen grains. Subsequently, they were stored in the entomological collection of the Integrated Laboratory of Invertebrate Biology of the Institute of Biodiversity and Sustainability of the Federal University of Rio de Janeiro (NUPEM/UFRJ).
Floral Biology and Specialization of Floral Visitors
The floral morphology data of C. rosea were obtained from the analysis of fresh flowers in an optical stereomicroscope, using specialized literature as a reference [32, 33]. The duration of anthesis was determined by monitoring flowers from pre-anthesis to petal fall.
To assess whether the floral visitors had pollen types other than C. rosea pollen, the pollen grains present in the floral visitors’ bodies were removed with the aid of a sterile needle. The pollen present at each site of the body was recorded as an individualized sample, from which fresh slides were made using gelatin [34]. The identification of pollen grains was made by comparison with a reference slide made with C. rosea pollen. Pollens from other plants were categorized into morphotypes in order to quantify the number of plants used by floral visitors. For analysis purposes, the pollen present in the three most representative genera were evaluated.
Statistical Analysis
Differences were compared using the Student-Newman- Keuls post-hoc test and correlations were made using Pearson’s coefficient (r). Data were processed and analyzed using Sigma Plot v. 2.1.
Results
The flowering period of Canavalia rosea was considered long, flowering for six consecutive months, from October 2019 to March 2020. Flowering occurred during spring and summer, with the highest number of open flowers observed in November (n = 3155), January (n = 4365) and March (n = 3578) (Table 1).
| Species/month | Oct-19 | Nov-19 | Dec-20 | Jan-20 | Feb-20 | Mar-20 | Total |
|---|---|---|---|---|---|---|---|
| Apidae | |||||||
| Centris sp. 1 | 1 | 5 | 5 | 4 | 4 | 2 | 21 |
| Centris sp. 2 | - | - | - | - | 1 | - | 1 |
| Centris sp. 3 | - | - | - | 1 | - | - | 1 |
| Centris sp. 4 | - | - | 1 | 4 | 1 | 2 | 8 |
| Centris sp. 5 | - | - | - | - | 2 | - | 2 |
| Centris sp. 6 | - | - | - | - | 2 | 1 | 3 |
| Centris sp. 7 | - | - | 1 | 12 | 7 | 8 | 28 |
| Epicharis sp. 1 | - | - | 1 | - | - | - | 1 |
| Epicharis sp. 2 | - | - | - | 1 | 5 | 2 | 8 |
| Eufriesea dentilabris | 1 | 40 | 19 | 12 | 3 | - | 75 |
| Eulaema nigrita | - | 1 | 0 | 1 | - | 8 | 10 |
| Mesoplia sp.1 | - | 1 | - | 2 | 1 | - | 4 |
| Xylocopa sp. 1 | 1 | - | - | 12 | 1 | 4 | 18 |
| Xylocopa truxali | - | - | - | 1 | 2 | 9 | 12 |
| Total abundance per month | 3 | 47 | 27 | 50 | 29 | 36 | 192 |
| Bee richness per month | 3 | 4 | 6 | 10 | 11 | 8 | 14 |
| Number of open flowers | 478 | 3155 | 1938 | 4365 | 2078 | 3578 | 15592 |
| Open flowers/m2 | 1,0 | 6,3 | 3,9 | 8,7 | 4,2 | 7,2 | - |
Table 1: Richness and abundance of bees and number of open flowers of C. rosea collected during the sampling months (2019/2020) i
C. rosea presents a racemose inflorescence (Figure 2A&2B), with specialized flowers that present differentiated petals in a vexillum (the lowest petal of the corolla), an external pair called wings and an internal pair of petals united by the margin forming the keel (Figure 2C-2E). At the base of the vexillum, an integumented region with a greenish-white color is observed and the rest of the petal has a different color pattern (Figures 2B-D). The petals of the wing and keel present a spatulate to falciform morphology and an acute auricle present in each petal, with a leathery and dilated texture (Figure 2C). The reproductive organs remain included in the keel and wing (Figure 2D-2I), being exposed only during contact with the floral visitor (Figure 2J-K).
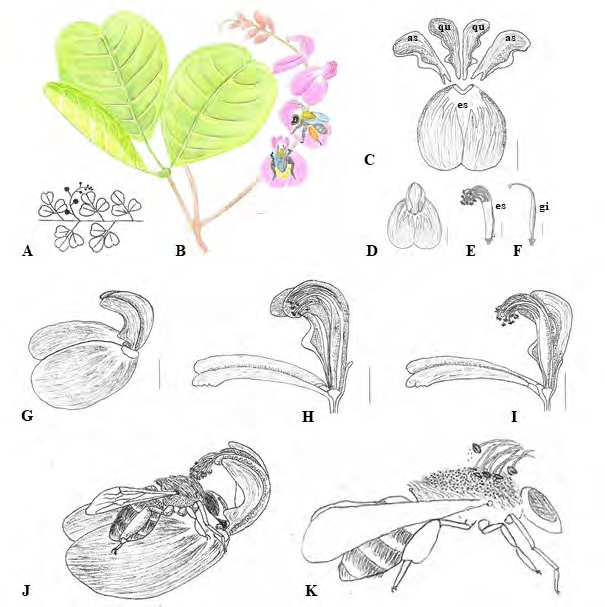
Figure 2: Floral morphology of Canavalia rosea. A-B. General characteristics of the plant: alternate phyllotaxis, trifoliate compound leaf, racemose inflorescence. C. Corolla petals: vexillum (es), wings (as), keel’s petals (qu). D. Flower in front view. E. Monadelphic stamens(es) with free anthers. F. Gyneceus: ovary, style and stigma. G. Flower section in side view, wings and keel in natural position. H-I. Flower section in side view, wings and keel in displaced position with reproductive whorls exposed. J-K Out of scale diagram, in (J) flower in side view with floral visitor (model Eulaema sp.), wings and keel in displaced position due to bee visit and in (K) detail of pollen transfer from the anthers of C. rosea for the bee’s dorsal scope. Bars: 1 cm.
Floral anthesis is diurnal, starting between 6 and 7 am, characterized by the distension and reflection of the vexillum. The duration of the flowers is short, approximately 9 hours. The floral senescence process is initially characterized by the drying of the vexillum, followed by the fall of the other petals.
During anthesis, bees were observed visiting the flowers to collect nectar. In field observations, the bees exhibited behavior of flying over the flower until they landed on the vexillum, from where they force the dilated region of the auricle of the wing and keel with their heads, pushing the petals back and guiding the tongue along the base of the vexillum until reach the nectary region (Figure 2J). The stamens and stigma, previously included and hidden, become apparent and touch the dorsal scopa of the bee, enabling the transfer of pollen (Figure 2K).
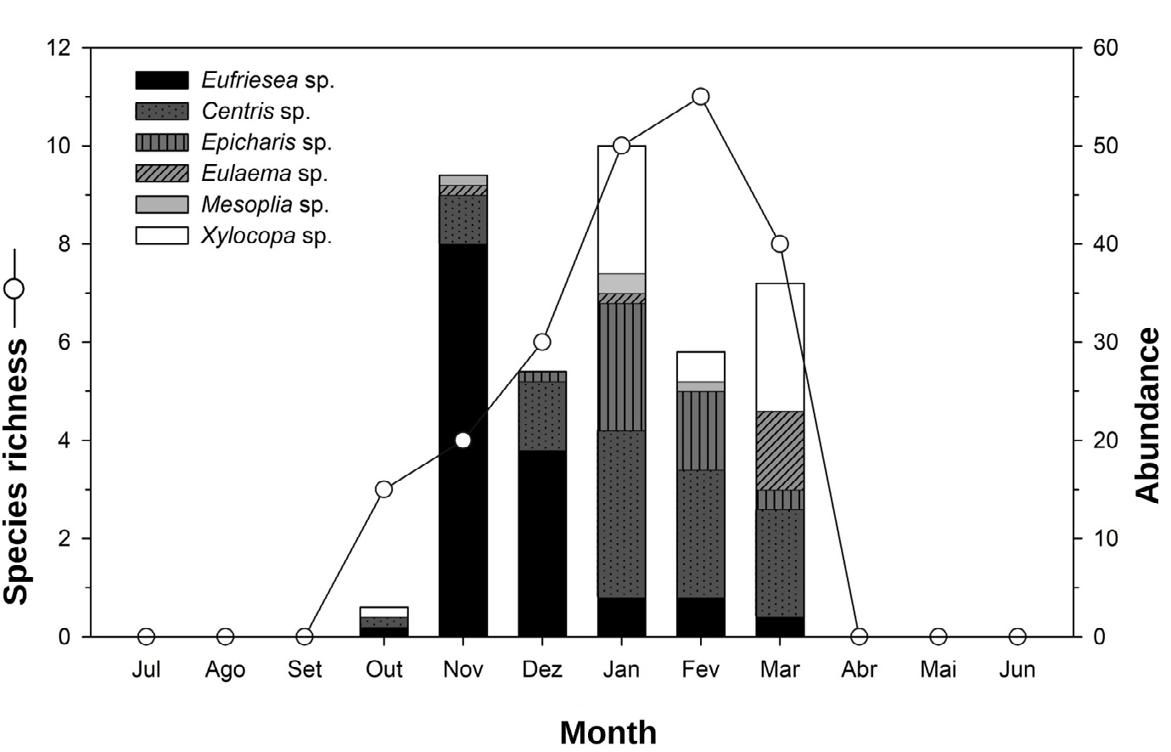
A total of 192 bees were collected, belonging to the Apidae family, distributed in 06 genera and 14 species (Table 1). The months with the highest richness of bee species were during the summer period, January (n = 10), February (n = 11) and March (n = 8) and the highest peaks of abundance were observed in spring, November (n = 47) and in summer, January (n = 50) (Table 1; Figure 3). It was observed from October to December a greater abundance of Eufriesea dentilabris and in the period from January to March an increase in the occurrence of Xylocopa sp (Figure 3). The Centris bees genus were observed throughout all flowering months (Figure 3).
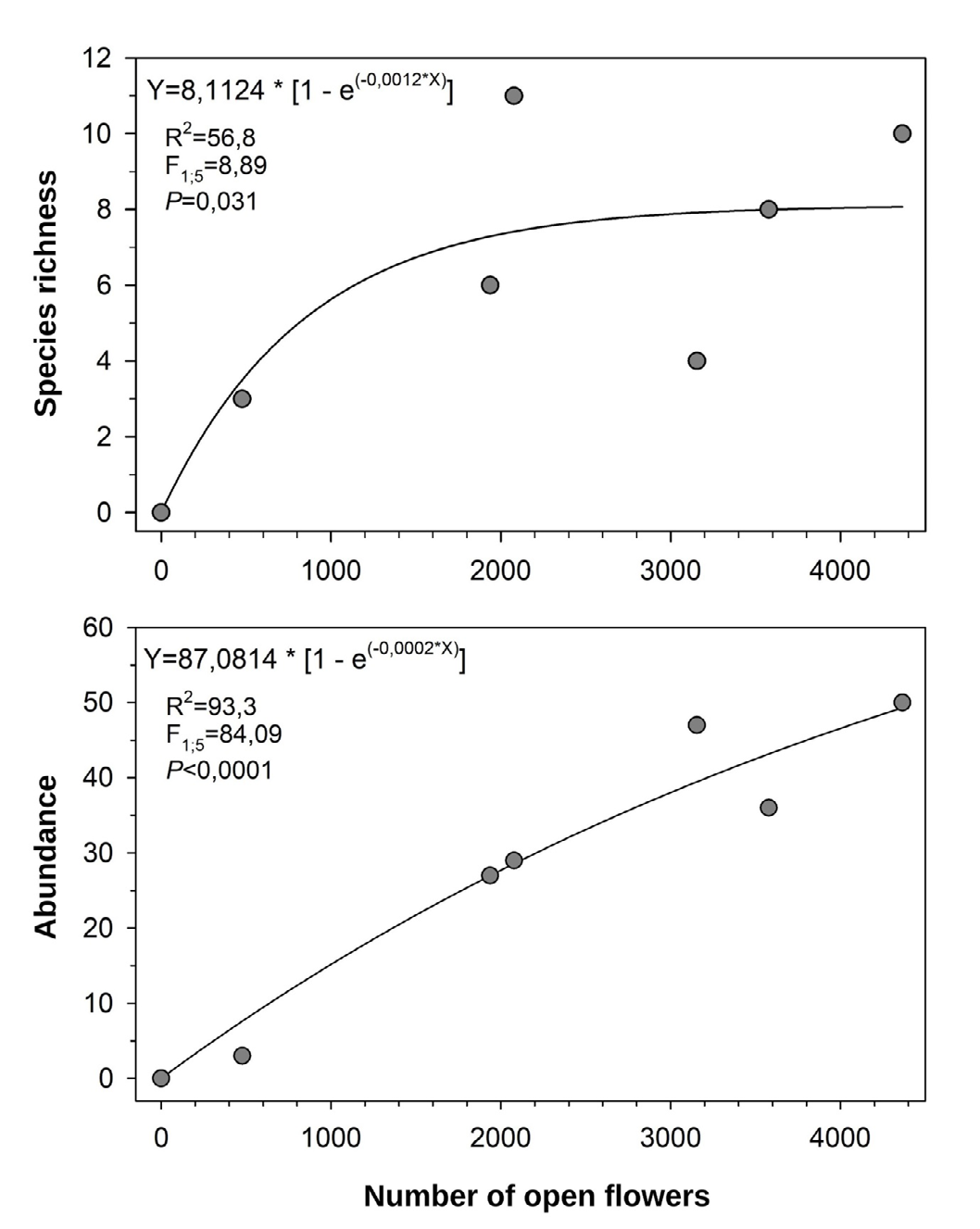
Linear regression analysis showed that the richness (F1;5 = 8.89; P < 0.031; Figure 4) and the abundance (F1;5 = 84.09; P < 0.0001; Figure 4) of species of bees were positively related to the number of open flowers of C. rosea.
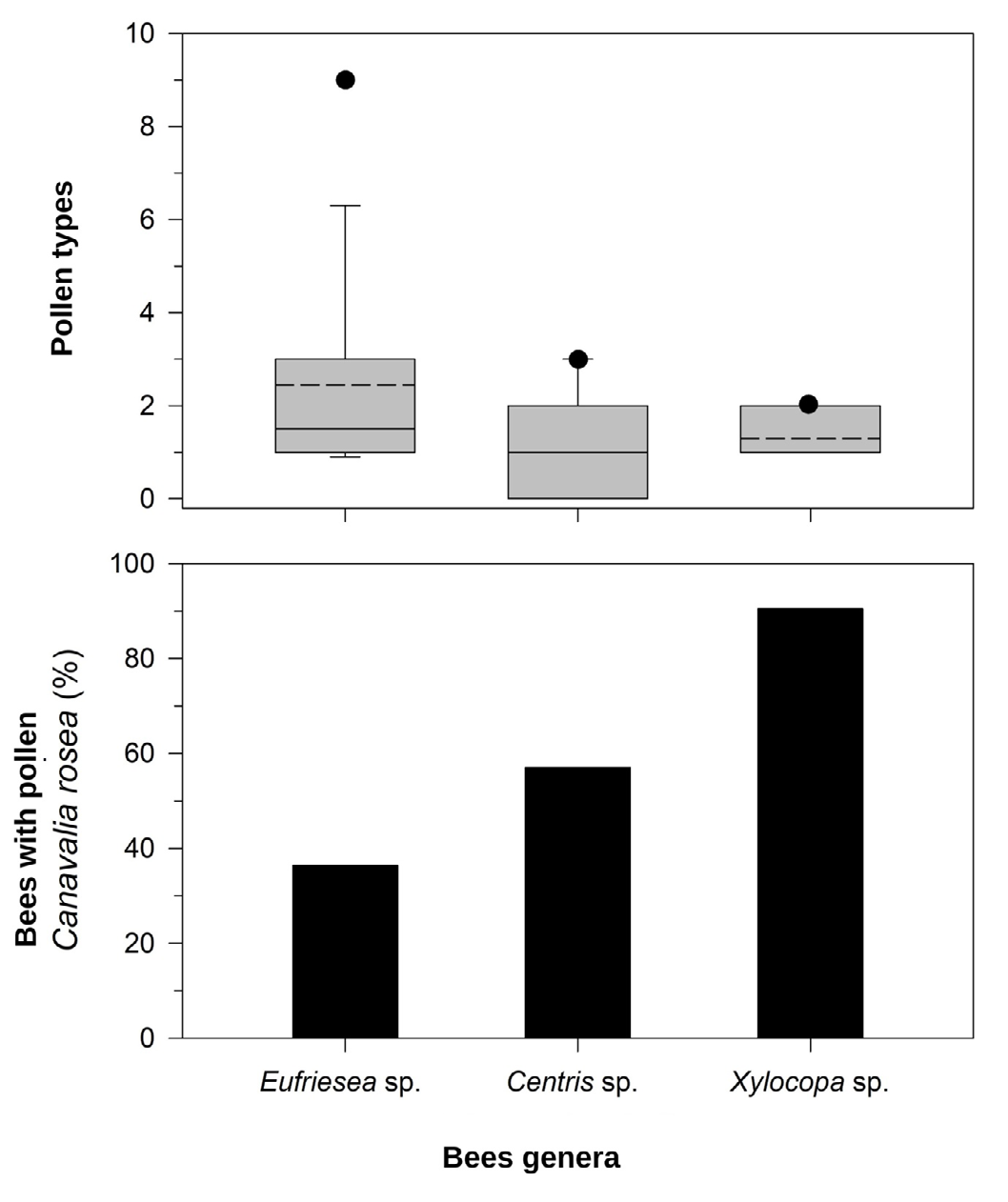
The analyzed pollen grains were collected from species of the genera Centris, Eufriesea and Xylocopa. The genus with the greatest range of pollen types collected was Eufriesea (n = 9; Figure 5). For the genus Xylocopa, pollen grains of C.
rosea were found in 90% of the specimens and located in the dorsal scopa, with variation of one and two pollen types among the individuals sampled (Figure 5).
Discussion
The flowering pattern of C. rosea is long and continuous, as described in other studies [33, 35, 36]. Flowering occurred in spring/summer, months with the highest averages of temperature and rainfall. In the Restinga of Maricá (RJ) a short flowering period was observed between the months of March and May [37], while in the state of Maranhão, despite a continuous flowering, a flowering peak was observed in the months August and September [11]. The different flowering patterns of C. rosea may be associated with regional and annual climatic differences, since precipitation and temperature can directly affect the phenology of plants [38, 39, 40].
The floral morphology of C. rosea has already been described in the literature, highlighting the presence of floral structures such as the vexillum that favors interaction with the visitor and maximizes pollination, thus configuring a specialized pollination system [33, 35]. Also, the color pattern from the base to the apex of the vexillum possibly signals the location for the bees to land and serves as a guide to direct the visitor’s tongue to the flower nectary [41, 42].
Among the floral visitors of C. rosea, species of the genus Xylocopa are identified as the most abundant [33, 43, 44, 45, 46]. In the present study, of the 14 species sampled, the Xylocopa species were the most representative as possible pollinators, since 90% of the analyzed individuals had C. rosea pollen grains in the dorsal scopa. The high frequency of Xylocopa visiting C. rosea has already been demonstrated in Brazilian Restingas in Maranhão [44] and in Grumari, Rio de Janeiro [46]. This pattern has also been reported in other countries such as Costa Rica [43], Indonesia [33] and Puerto Rico [45]. The high frequency of Xylocopa was also observed visiting other species of Canavalia, such as Canavalia brasiliensis [47, 48], Canavalia ensiformis [49], Canavalia lineata [50] and Canavalia paraguariensis [51].
The body size of Xylocopa species allows them to fly even in strong winds and withstand high radiation and temperatures in the Restinga [10, 11, 12]. In addition, the body of these bees is robust, which makes it possible to efficiently push the specialized petals of the flower, leading to the precise deposition of pollen in the bee’s dorsal scope, while in smaller bees the pollen is dispersed throughout the body, which can cause a smaller pollination efficiency [44].
Some studies have already described the species of floral visitors, aspects of pollination and nutritional value of C. rosea fruits [9, 52, 53, 54]. However, these studies did not evaluate the richness and abundance during the flowering months of C. rosea, and it was not possible to establish a comparison between the Restinga areas, considering the size of the fragments or the degree of human threats. Specimens of bees of the genera Xylocopa, Centris and Eufriesea, described in this study, are also among the frequent floral visitors in plant communities associated with the Restinga [55, 56, 57].
The results showed that the richness and abundance of bees associated with C. rosea responded positively to the availability of floral resources (open flowers). Bees are sensitive to environmental responses associated with resource offerings [58, 62]. The effects of seasonality influence the availability of floral resources, such as the amount of flowers and the reward to visitors (nectar and pollen), changing the structure of the bee community [62, 63, 64].
Bees associated with C. rosea, in general, are solitary and their populations fluctuate throughout the year according to the species of flowering plants, with a positive relationship between the richness of plant and bee species [62, 64, 65, 66]. Times of the year with a greater variety of resources can support species with different ecological needs [67], which may explain the greater richness in the summer months, where higher rainfall and temperature can result in a greater amount of available floral resources. The supply of nectar, for example, is considered an important factor in structuring the composition of the pollinator community [62]. The decline in the abundance of Eufriesea dentilabris in the flowering period of C. rosea from January to March may be related to the increase in the supply of floral resources by other plant species during the summer, which would explain the lower abundance of C. rosea in this period.
In the sandy cord C. rosea forms resource patches associated with other plant species, such as Ipomoea sp. [43; 44, 33), which offer attractive resources for bees. Analyzes of pollen types showed that bees visiting C. rosea are generalists [35]. However, medium to large pollinators are characteristic of the Canavalia genus, since floral biology selects visitors who are able to reach the nectaries, which also restricts the occurrence of a greater diversity of associated bees.
The floral structure of Canavalia species is similar to each other [35, 60] which explains this pattern of bees associated with the genus. The floral visitors of C. rosea by pushing the wing and keel petals with their head to reach the nectary, consequently, promote the exposure of the flower’s reproductive organs, causing the deposition of pollen grains in the bee’s body, allowing further pollination to occur [44]. This pattern of flower behavior explains the fact that bees collected in C. rosea have pollen grains deposited mainly on the dorsal scopa [33]. In this study, we observed that the pollen grains of C. rosea were adhered to the dorsal scopa of the species of Centris, Eufriesea and Xylocopa, different from the pattern observed in Japan for Megachile bees, where the pollen was mainly in the scopa of the hind legs [61].
Conclusion
The present study is pioneering research in the Restinga area of the recently created Municipal Natural Park of Barreto’s Restinga and the data generated are relevant for future management plans for the area, since they contribute with ecological data to the understanding of the dynamics of flowering, availability of floral resources of C. rosea and the associated bee fauna.
References
-
Myers N, Mittermeier RA, Mittermeier CG, DaFonseca GA, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403: 853-858.
-
Santos JFC, Mendonça BAF, De Araújo EJG, De Andrade CF (2017) Fragmentação florestal na Mata Atlântica: o caso do município de Paraíba do Sul, RJ, Brasil. Revista Brasileira de Biociências 15(3): 151-158.
-
Freire MSB (1990) Levantamento florístico do Parque Estadual das Dunas do Natal. Acta Botanica Brasilica 4: 41-59.
-
Assumpcão J, Nascimento MT (2000) Estrutura e composição florística de quatro formações vegetais de Restinga no complexo lagunar Grussaí/Iquipari, São João da Barra, RJ, Brasil. Acta Botanic Brasilica 14: 301-315.
-
Dillenburg LR, Waecjter JL, Porto ML (1992) Species composition and structure of a sandy coastal plain forest in northern Rio Grande do Sul, Brazil. Coastal plant communities of Latin America Academic Press pp: 349- 366.
-
Lamêgo AR (1974) The Man and the Sandbank. Lidador, Rio de Janeiro, pp: 307.
-
Araújo DSD (1992) Vegetation types of sandy coastal plains of tropical Brazil: a first approximation. Coastal plant communities of Latin America Academic Press pp: 337-347.
-
Mendoza-González G, Martínez ML, Lithgow D (2014) Biological flora of coastal dunes and wetlands: Canavalia rosea (Sw.) DC. Journal of Coastal Research 30: 697-713.
-
Sridhar KR, Seena S (2006) Nutritional and antinutritional significance of four unconventional legumes of the genus Canavalia – A comparative study. Food Chemistry 99: 267-288.
-
Viana BF, Kleinert AMP (2005) A community of flower- visiting bees (Hymenoptera: Apoidea) in the coastal sand dunes of northeastern Brazil. Biota Neotropica 5: 1-13.
-
Albuquerque PMC, Camargo JMF, Mendonça JA (2007) Bee community of a Beach Dune ecosystem on Maranhão Island, Brazil. Braz Arch Biol Tech 6: 1005-1018.
-
Oliveira FS, Mendonça MW, Vidigal M, Rêgo M, Albuquerque P (2010) Comunidade de abelhas (Hymenoptera, Apoidea) em ecossistema de dunas na praia de Panaquatira, São José de Ribamar, Maranhão, Brasil. Revista Brasileira de Entomologia pp: 82-90.
-
Allen-Wardell G, Bernhardt P, Bitner R, Burquez A, Buchmann S, et al. (1998) The potential consequences of pollinator declines on the conservation of biodiversity and stability of food crop yields. Conserv Biol 12: 8-17.
-
Biesmeijer JC, Roberts SP, Reemer M, Ohlemüller R, Edwards M, et al. (2006) Parallel declines in pollinators and insectpollinated plants in Britain and the Netherlands. Science 313(5785): 351-354.
-
Gallai N, Salles JM, Settele J, Vaissière BE (2009) Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecological Economics 68(3): 810-821.
-
Potts SG, Biesmeijer JC, Kremen C, Neumann P, Schweiger O, et al. (2010) Global pollinator declines: trends, impacts and drivers. Trends in Ecology & Evolution 25(6): 345- 53.
-
Goulson D, Nicholls E, Botías C, Rotheray EL (2015) Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 347(6229): 1435-1446.
-
Pettis JS, Lichtenberg EM, Andree M, Stitzinger J, Rose R (2013) Crop pollination exposes honey bees to pesticides which alters their susceptibility to the gut pathogen Nosema ceranae. PloS One 8: 1-9.
-
Vanbergen AJ (2013) Threats to an ecosystem service: pressures on pollinators. Frontiers in Ecology and the Environment 11(5): 251-259.
-
De Marco P, Coelho FM (2004) Services performed by the ecosystem: forest remnants influence agricultural cultures’ pollination and production. Biodiversity & Conservation 13: 1245-1255.
-
Vitousek PM, Mooney HA, Lubchenco J, Melillo JM (1997) Human domination of Earth’s ecosystems. Science 277: 494-499.
-
Mcfrederick QS, Lebuhn G (2006) Are urban parks refuges for bumble bees Bombus spp. (Hymenoptera: Apidae)? Biological conservation 129(3): 372-382.
-
Marzluff JM, Ewing K (2001) Restoration of fragmented landscapes for the conservation of birds: a general framework and specific recommendations for urbanizing landscapes. Restoration Ecology 9: 280-292.
-
Koh LP, Sodhi NS (2004) Importance of reserves, fragments, and parks for butterfly conservation in a tropical urban landscape. Ecological Applications 14(6): 1695-1708.
-
Cane JH, Minckley RL, Kervin LJ, Roulston TAH, et al. (2006) Complex responses within a desert bee guild (Hymenoptera: Apiformes) to urban habitat fragmentation. Ecological Applications 16: 632-644.
-
Holzer W, Crichyno J, Pires AC (2004) Sustentabilidade da urbanização em áreas de Restinga: uma proposta de avaliação pós-ocupação. Paisagem e ambiente 19: 49-65.
-
Fantinato E (2019) The impact of (mass) tourism on coastal dune pollination networks. Biological Conservation 236: 70-78.
-
Silva SM (1999) Diagnóstico das Restingas no Brasil. Avaliação e ações prioritárias para a conservação da zona costeira e marinha. In: Panorama da conservação dos ecossistemas costeiros e marinhos no Brasil. Ministério do Meio Ambiente (MMA), Brasília pp: 157.
-
Köppen W, Geiger R (1928) Klimate der Erde. Gotha: Verlag Justus Perthes. Wallmap 150cmx200cm.
-
(2020) Instituto Nacional de Meteorologia.
-
Newstrom LE, Frankie GW, Baker HG (1994a) A new classification for plant phenology based on flowering patterns in lowland tropical rain forest trees at La Selva, Costa Rica. Biotropica 26: 141-159.
-
Percival M (1965) Floral Biology. Pergamon Press, Oxford pp: 243.
-
Gross CL (1993) The reproductive ecology of Canavalia rosea (Fabaceae) on Anak Krakatau, Indonesia. Australian Journal of Botany 41: 591-599.
-
Barth OM (1989) O pólen no mel brasileiro. Instituto Oswaldo Cruz, Rio de Janeiro.
-
Sauer J (1964) Revision of Canavalia. Brittonia 16: 106- 181.
-
Vatanparast M, Takayama K, Sousa MS, Tateishi Y, Kajita T (2011) Origin of Hawaiian endemic species of Canavalia (Fabaceae) from sea-dispersed species revealed by chloroplast and nuclear DNA sequences. Journal of Japanese Botany 86: 15-25.
-
Gonçalves IP, Gama MC, Correia MCR, De Lima HA (2008) Caracterização dos frutos, sementes e germinação de quatro espécies de leguminosas da Restinga de Maricá, Rio de Janeiro. Rodriguésia 59: 497-512.
-
Talora DC, Morellato LPC (2000) Fenologia de espécies arbóreas em floresta de planície litorânea do sudeste do Brasil. Revista Brasileira de Botânica 23(1): 13-26.
-
Pedroni F, Sanchez M, Santos FAM (2002) Fenologia da copaíba (Copaifera langsdorffii Desf. Leguminosae, Caesalpinioideae) em uma floresta semidecídua no sudeste do Brasil. Revista Brasileira de Botânica 25: 183- 194.
-
Marques MCM, Roper JJ, Salvalaggio APB (2004) Phenological patterns among plant life-forms in a subtropical forest in southern Brazil. Plant Ecology 173: 203-213.
-
Silva MF, Carmo MNL, Gurgel ESC (2015) Phaseoleae (Leguminosae - Papilionoideae) nas Restingas do Estado do Pará, Brasil. Biota Amazônia 5: 14-22.
-
Queiroz LP, Snak C (2016) Flora da Bahia: Leguminosae – Canavalia (Papilionoideae: Diocleae). Sitientibus série Ciências Biológicas 6: 1-9.
-
Sage RD (1968) Observations on feeding, nesting, and territorial behavior of carpenter bees genus Xylocopa in Costa Rica. Annals of the Entomological Society of America 61(4): 884-889.
-
Gottsberger G, Camargo JMF, Silberbauer-Gottsberger I (1988) A bee pollinated tropical community: The beach dune vegetation of Ilha de São Luís, Maranhão, Brazil. Botanische Jahrbucher fur Systematik 109: 469-500.
-
Vargas MM (2000) Características de las comunidades de plantas y sus polinizadores en la Isla de Mona, Puerto Rico. Acta Científica 14: 29-44.
-
Verçoza FC, Do Nascimento EC, Côrtes IMR (2010) Melitofilia em Canavalia rosea (Sw.) DC. (Leguminosae: Papilionoideae). EntomoBrasilis 3(3): 73-76.
-
Guedes RS, Quirino ZGM, Gonçalves EP (2009) Fenologia reprodutiva e biologia da polinização de Canavalia brasiliensis Mart. ex Benth (Fabaceae). Biotemas 22: 27- 37.
-
Amaral-Neto LP, Westerkamp C, Melo G (2015) From keel to inverted keel flowers: functional morphology of “upside down” papilionoid flowers and the behavior of their bee visitors. Plant Systematics Evol 301: 2161- 2178.
-
Silva CI, Freitas BM (2018) Rearing carpenter bees (Xylocopa spp.) for crop pollination: a case study with passionfruit (Passiflora edulis) In: Roubik (Ed.), The pollination of cultivated plants: a compendium for practitioners. FAO, Rome.
-
Sugiura S (2008) Male territorial behaviour of the endemic large carpenter bee, Xylocopa (Koptortosoma) ogasawarensis (Hymenoptera: Apidae), on the oceanic Ogasawara Islands. European Journal of Entomology 105: 153-157.
-
Schlindwein C, Schlumpberger B, Wittmann D, Moure JS (2003) O gênero Xylocopa Latreille no Rio Grande do Sul, Brasil (Hymenoptera, Anthophoridae). Revista Brasileira de Entomologia 47: 107-118.
-
Seena S, Sridhar KR, Bhagya B (2005) Biochemical and biological evaluation of an unconventional legume, Canavalia maritima of coastal sand dunes of India. Tropical and subtropical Agroecosystems 5: 1-14.
-
Bhagya B, Sridhar KR, Raviraja NS, Young CC, Arun AB (2009) Nutritional and biological qualities of the ripened beans of Canavalia maritima from the coastal sand dunes of India. Comptes Rendus Biologies 332(1): 25-33.
-
D’Cunha M, Sridhar KR, Bhat, R (2009) Nutritional quality of germinated seeds of Canavalia maritima of coastal sand dunes. In: Food processing: methods, techniques and trends. Nova Science Publishers Inc., New York, USA, pp: 363-384.
-
Mendes GA (2014) As abelhas e a floração sequencial em área de Restinga. Universidade Federal do Espírito Santo, Vitória, Brasil pp: 90.
-
Patricio RS (2015) Abelhas e suas plantas visitadas em uma área de Restinga no Extremo Sul de Santa Catarina. Universidade do Extremo Sul Catarinense (UNESC), Santa Catarina, Brasil, pp: 63.
-
Receputi ALM (2015) Fauna de abelhas nativas de duas áreas de Restinga do Parque Estadual de Itaúnas, Espírito Santo, Brasil. Universidade Federal do Espírito Santo, São Mateus, Brasil, pp: 114.
-
Michener CD (2000) The Bees of the World. Johns Hopkins Press, Washington.
-
Silveira FA, Melo GAR, Almeida EAB (2002) Abelhas brasileiras. Sistemática e Identificação. Fundação Araucária, Belo Horizonte.
-
Stirton CH (1977) The pollination of Canavalia virosa by Xylocopid and Magachilid bees. Bothalia 12(2): 225-227.
-
Matsumura SI, Tateishi Y, Yokoyama J, Maki M (2004) Expansion of the distribution range and pollinators of Canavalia rosea (Fabaceae) in the Ryukyu Archipelago, Japan. Acta Phytotaxonomica et Geobotanica 55: 207- 212.
-
Potts SG, Vulliamy B, Roberts S, O’Toole C, Dafni A, Ne’Eman G, Willmer PG (2004) Nectar resource diversity organizes flower‐visitor community structure. Entomologia Experimentalis Et Applicata 113(2): 103- 107.
-
Klein A, SteffanDewenter I, Tscharntke T (2004) Foraging trip duration and density of megachilid bees, eumenid wasps and pompilid wasps in tropical agroforestry systems. Journal of Animal Ecology 73(3): 517-525.
-
Araújo VA, Araújo APA, Antonini Y (2010) Impact of resource availability on bee diversity. Sociobiology 55: 1-13.
-
Klein A, SteffanDewenter I, Tscharntke T (2003) Fruit set of highland coffee increases with the diversity of pollinating bees. Proceedings of the Royal Society of London 270(1518): 955-961.
-
Ricketts TH (2004) Tropical forest fragments enhance pollinator activity in nearby coffee crops. Conservation biology 18(5): 1262-1271.
-
Tilman D, Pacala S (1993) The Maintenance of Species Richness in Plant Communities. In: Ricklefs RE, Schluter D, et al. (Eds.), Species Diversity in Ecological Communities: Historical and Geographical Perspectives. University of Chicago Press, Chicago, USA, pp: 13-25.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework