Asymmetric Plastral Allometry between Sexes in Captive Yellow- Bellied Slider Turtles (Trachemys scripta scripta Schoepff 1792)
This study aimed to contribute to a better understanding of the role of asymmetric shape changes related to variation in size in the yellow-bellied slider turtles (Trachemys scripta scripta), imported in Spain as pets. Values were obtained from geometric morphometric analysis of 17 anatomical landmarks on plastron (ventral aspect of the body) from 33 males and 63 females. The allometry was described by a multivariate regression where the independent variable was size, expressed as centroid size, and dependent variables was the set of shape coordinates. The results demonstrated the existence of sex-related differences in plastron asymmetries, but, interestingly, the allometric slopes were not sexually dimorphic. The expression of these allometric trajectories may be the reflection of some functional features, such as feeding or movement.
Introduction
Turtles show a very large difference from hatchling to adult size [1], ranging from a few centimeters to over three meters in length, and this provides the basis for large ontogenetic variations of shape in this group [1].
Static allometry refers to the relationship between the shape of a trait and body size across individuals at the same developmental stage [2, 3]. Changes in body proportions (length vs. width) have been observed in aquatic turtle species and are explained as an antipredator adaptation [4]. External linear morphometry, mainly head and carapace, have been used to investigate allometry in turtles [5, 6, 7], but for some species, morphological variation, in terms of allometry, is far from being fully understood. For its capacity of evaluating shape [8]. we consider geometric morphometrics (GM) having a great potential to the study of allometry in turtles, without the limitations and the laboriousness of linear morphometrics. GM is based on the Cartesian coordinates of landmarks (measurement points) that are homologous across all measured individuals [9].
The plastron of turtles contains, as general pattern, twelve bilateral scutes (gular, humeral, pectoral, abdominal, femoral and anal) and one central (intergular) [10]. It is ideal for object asymmetry studies due to its bilateral characteristics and relative flat surface, which allows for easy digital measurements from photographs. Sliders, specifically, possess a very dorsoventrally flattened body shape and lives on water but also on land, thus providing an easy model for evaluating the effects of morphological design. Another pro of sliders is that many species are easily available for studies, because they are commonly kept as pets.
The yellow-bellied slider (Trachemys scripta Schoepff 1792) belongs to the family of true pond turtles (Emydidae; subfamily Emydinae) [5, 11]. It is native to the southeastern United States, being the most common turtle species in its range [5], and occupy a wide variety of habitats, including slow-moving rivers, floodplain swamps, marshes, seasonal wetlands, and permanent ponds [5, 10].
Yellow-bellied sliders are very popular as pets [11]. From 1983 to 1997 500,000 to 900,000 specimens were introduced to Spain where many escaped or were released into the wild, where it seems to have established stable populations [12]. But, as the species has negative impacts on native pond turtle population fitness and can act as parasite and disease vectors, non-captive specimens are captured and legally euthanized [12], so these corpses could be used in studies like the presented here with no ethical restriction.
In order to contribute to the current knowledge about some allometric aspects of sliders, we applied a geometric morphometrics approach to yellow-bellied slider plastron of a wide size range, and from two genders, with the aim of illustrating the role of changes in proportions related to variation in size in shaping the external morphology. In particular, the purposes of the research were: (1) quantifying and determining the types of shape asymmetry of plastron; and (2) assess if this asymmetry has an allometric component. Our final goal is to provide some additional information that can facilitate a better understanding of expression of asymmetry of this species, complementing previous studies on this same species [13, 14].
These findings will represent a contribution to the study of growth models in freshwater turtles and serve as a baseline for (1) comparisons with other wild and captive populations and species, and (2) future ecomorphological investigations aimed at understanding the adaptive meaning of different developmental trajectories in this group of testudines. This study is part of a larger project to study morphology in exotic and local land and water tortoises by means of geometric morphometric techniques.
Materials and Methods
Specimen Collection
A sample of 33 males and 63 females belonging to Trachemys scripta scripta were obtained from the CRARC. They had been collected as invasive species and so euthanized. Sex was determined visually by: 1) general size -females being larger-, 2) position of cloaca and thickness and length of tail -males having enlarged tails-, 3) claws -males having elongated foreclaws [5, 11]. All specimens used in the study were adult turtles without any detectable abnormality, such as injuries or unusual additional scutes or plates. As study was done on specimens euthanized according to the European law, authors considered not necessary the obtention of an ethical agreement.
Data Collection and Geometric Morphometric Analyses
Turtles were labelled and levelled on a horizontal plane, and then photographed on the ventral view (plastron). Image capture was performed with a Nikon® D70 digital camera (image resolution of 2,240 x 1,488 pixels) equipped with a Nikon AF Nikkor® 28-200 mm telephoto lens. The camera was placed parallel to the ground plane so the focal axis of the camera was parallel to the horizontal plane and centered on each plastron. A scale was included in the images to standardize each specimen size (mm unit). Pictures were posteriorly digitized using tpsDig version 2.04 [15]. In total, 17 two-dimensional (2D) landmarks (LMs, i.e., discrete and homologous loci in the space characterized by their x- and y-coordinates, and identified without ambiguity as the same loci in all investigated specimen) were used on the ventral side of plastron (Figure 1), excluding exterior marginal scutes. Ten of them were bilateral and seven were midline. These landmarks corresponded to the intersections between bony plates or epidermal scutes, and thus were type 1 or 2 according to Bookstein’s nomenclature [16]. A double digitalization was done for assessing the measurement error.

Cartesian x-y coordinates were then extracted with a full Procrustes fit in order to remove information about position, orientation and rotation and standardize each specimen to unit centroid size (CS -computed as the square root of the summed squared Euclidean distances from each landmark to the specimen centroid, and providing the estimation of the size [16]. Following the Procrustes fit, we used Procrustes ANOVA (ANalysis Of VAriance), to quantify the amount of asymmetric variation; results are reported as sum of squares (SS) and means squares (MS) [17]. The coordinates of the superimposed landmarks to assess asymmetry of shape were in relation to individuals (symmetric component of variation), plastron sides (directional asymmetry, DA, occurring when there is a greater development of a character at one side of the body plan of symmetry than at the other, with most individuals being asymmetrical in the same direction), their interaction (fluctuating asymmetry, FA, small and random deviations in the symmetry of bilateral characters of organisms, with a mean equal to zero and variation normally distributed around that mean) [18], and measurement error. To avoid making the assumption of equal, independent and isotropic variation of landmarks, we added a MANOVA approach to test for asymmetries; significance was assessed with the Pillai trace statistic [17].
Then, we used PCA (Principal Component Analysis) to summarize the sample shape variation into fewer components. The first principal component (PC) is the shape pattern (linear combination of shape coordinates) with maximum variance in the sample and it can be visualized as a shape deformation or a series of shapes, and a score along the PC could be computed for each individual. The second PC is geometrically orthogonal (perpendicular) to the first one and accounts for the second most variance, and similarly for all subsequent components.
Then we performed a multivariate regression of the Procrustes coordinates as shape variables (being used to calculate the regression score) on the log-transformed centroid size values (CS, the square root of the sum of squared distance between each landmark and the skull centroid) as a size variable to analyze the allometry trend of each gender [16]. Results are reported as a percentage value of the explained total shape variation from the size variation. Allometric trajectories were compared using the angular vectors. Results are reported as angular values of the pairwise angular comparisons. Levels of statistical significance were computed by permutation tests, using 9,900 random permutations. Analyses were done using MorphoJ version 1.05 [19] and PAST version 2.17 c [20] packages.
Results
The studied turtles ranged from 92.8 to 193.6 mm in total length for males (mean=143.4 mm; SD=19.72; n=33; mean CS=3318.69), and from 95.2 to 256.5 mm in total length for females (mean=183.3; SD=33.22; n=63; mean CS=3862.81).
The ANOVA tests whether individual variation is significantly larger than error. This could be potentially a very serious issue because random measurement error can inflate the amount of variance and, since many statistical analyses are based on the amount of “explained” relative to “residual” variance, can result in loss of statistical power. However, it should be noted that it was found a highly significant individual variation (mean squares=0.0001346881, p<0.001) larger than measurement error (mean squares= 0.0000455997). Procrustes ANOVA for the carapace were statistically significant, showing individual variability, DA and FA (Table 1). Similar results were obtained by analyzing the two sexes separately (data not shown here). DA accounted for the bulk of morphological variation (mean squares=0.0004735205, 71.0 % of the total variance). MANOVA tests confirmed only DA (Table 2).
| Effect | SS | Df | MS | F | p |
|---|---|---|---|---|---|
| Individual | 0.1411436 | 930 | 0.00015177 | 9.89 | <0.0001 |
| Sides | 0.0050113 | 15 | 0.00033409 | 21.8 | <0.0001 |
| Individual*Sides | 0.0142745 | 930 | 1.5349E-05 | 0.34 | 1 |
| Measurement error | 0.0865653 | 1890 | 4.5802E-05 |
Table 1: Output for Procrustes ANOVA for plastron asymmetry. Sums of squares (SS) and mean squares (MS) are in dimensionless unit
| Effect | Pillai trace | p |
|---|---|---|
| Side | 0.87 | <0.0001 |
| Individual*Sides | 8.9 | <0.0001 |
Table 2: Output for Procrustes MANOVA for plastron asymmetry.
In PCA, PC1 and PC2 explained a 28.7% and a 25.4%
of plastron asymmetric shape variance, respectively; they did not discriminate between males and females, thus corroborating the independent sexual asymmetric dimorphism. The graphical representation of PC1 and PC2 revealed the existence of directional differences mainly on gular and humeral plaques, indicating a greater development of the right side (Figure 2).
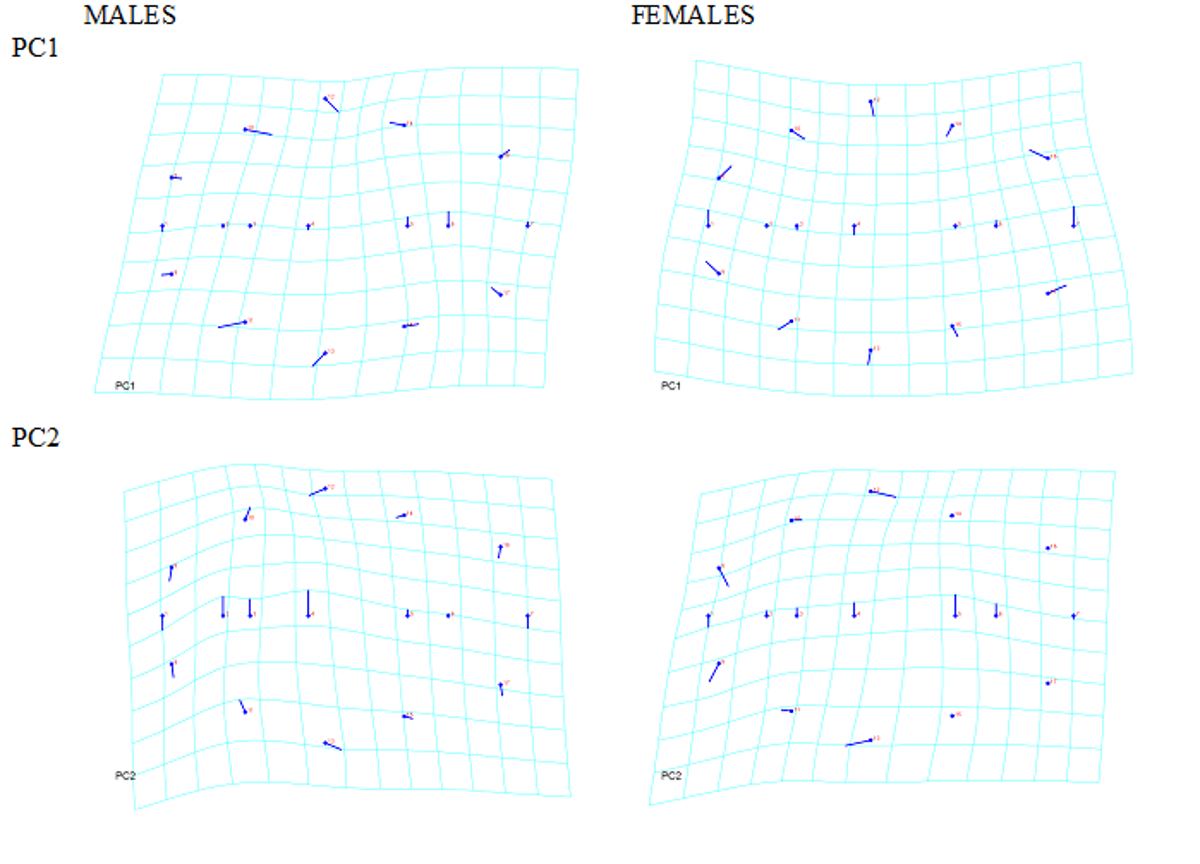
Figure 2: First Principal Component (PC1) and second Principal Component (PC2) of joint variation in landmark positions for individual directional asymmetry for males (n=33) and females (n=63), using the set of 17 landmarks. PC1 and PC2 explained a 28.7% and a 25.4% of plastron asymmetric shape variance, respectively; they did not discriminate between males and females. The graphical representation of PC1 and PC2 revealed the existence of directional differences between the left and right body shell sides mainly on gular and humeral plaques, indicating a greater development of the right side.
Regression of shape of size for symmetric component was not significative for males (P=0.652, 1.97 % of shape variation explained by size variation) but significative for females (P=0.0008), although it explained only a 6.80 % of shape variation, due probably to the larger shape variation. Correlation coefficients R2 for both genders were low (0.019 and 0.6 for males and females respectively). So relationships between centroid size of carapace and the ventral body parts suggest possible isometry in plastron for both genders. The angular comparisons between genders showed no significant distinction between their ontogenetic trajectories (P=0.813), with an angular value of 103.81, indicating similar patterns of joint displacements of landmarks (Figure 3). As it is thought that traits under sexual selection exhibit positive allometric relative to traits that are not under sexual selection [21, 22]. detected isometry in sliders could indicate that DA cannot be interpreted as conferring sexual advantages, but rather being adaptative traits.
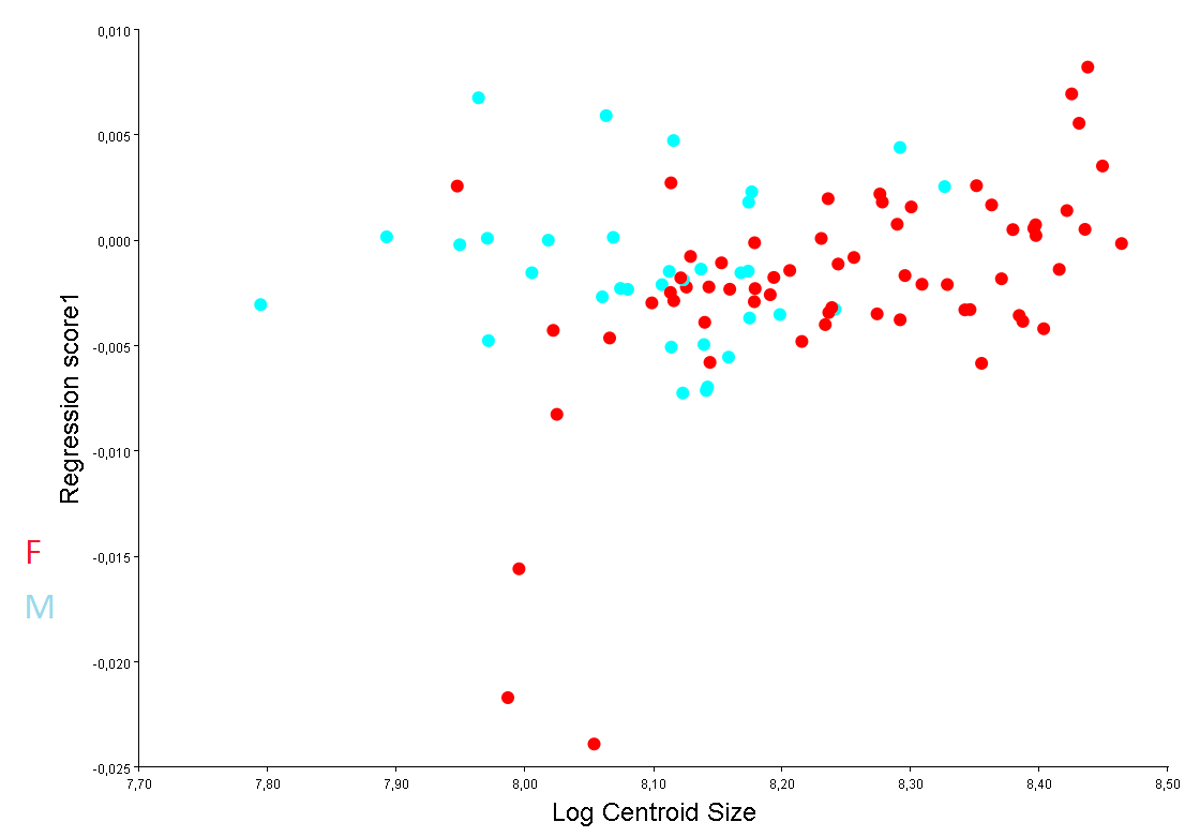
Figure 3: Regression trajectories for both genders of yellow-bellied slider turtle (Trachemys scripta scripta) by relating the centroid size Log-transformed values (Log Centroid Size) vs. the shape scores (that is. the regression score obtained from the Procrustes coordinates). Regression of shape of size for symmetric component was not significative for males (n=33; P=0.652; 1.97 % of shape variation explained by size variation) but significative for females (n=63; P=0.0008; 6.80 % of shape variation explained by size variation).
Discussion
In this study, the body shape asymmetric variation along size was described by comparing individuals of different sizes by multivariate regressions in yellow-bellied slider turtle (Trachemys scripta scripta). The method has allowed us to maintain the relative spatial configuration of landmarks. Here we used the entire body (the entire plastron), which considers the variation in form holistically and allowed us a more realistic view of the variation in shape in a deformation network. Nevertheless, our results differ from those by other authors [13]. Who did find no DA on this species.
Turtles used in the present study were kept in captivity, where they had to adapt to artificial environmental conditions. This factor could cause stress to animals, but the results of statistical analyses, indicating a low presence of FA cannot support the hypothesis that detected asymmetry is interpreted as a reaction to non-optimal environmental conditions [23]. DA was manifested as the main source of variation, and not being an indicator of stress, it must be interpreted as an adaptative trait.
The permutation test indicating the occurrence of ontogenetic allometries for each sex, although being significant, accounted for a low variation for both genders. Proportional changes occurred in several morphometric characters simultaneously from small to big specimens. Interestingly, the allometric slopes were not sexually dimorphic. So, the expression of these allometric trajectories seems to be not associated with age-specific shifts but linked to functional features such as feeding efficiency and movement.
Static allometry could be heavily influenced by management, as sliders have indeterminate growth influenced by variation in local conditions and feeding. Our study has been performed with initially captive (and then released) individuals, so it could be interesting to perform studies including other management conditions (such wild populations). Finally, the information obtained through such studies could be used to plan correct management strategies for local species. This research was not financially supported.
Acknowledgement
Authors wish to thanks to Joaquim Soler (CRARC) for his technical support during the selection of animals.
References
-
Angielczyk KD, Feldman CR (2013) Are diminutive turtles miniaturized? The ontogeny of plastron shape in emydine turtles. Biol J Linn Soc 108(4): 727-755.
-
Gould SJ (1966) Allometry and size in ontogeny and phylogeny. Biol Rev 41(4): 587-640.
-
Mitteroecker P, Gunz P, Windhager S, Schaefer K, Sonja W, et al. (2013) A brief review of shape, form, and allometry in geometric morphometrics, with applications to human facial morphology. Hystrix 24(1): 59-66.
-
Salmon M, Scholl J (2014) Allometric growth in juvenile marine turtles: Possible role as an antipredator adaptation. Zoology 117(2): 131-138.
-
Readel AM, Warner JK, Holberton RL, Phillips CA (2008) Maturational changes in male slider turtles (Trachemys scripta) from Illinois. Herpetol Conserv Biol 3(2): 170- 175.
-
Barros MS, Resende LC, Silva AG, Ferreira PD (2012) Morphological variations and sexual dimorphism in Chelonoidis carbonaria (Spix, 1824) and Chelonoidis denticulata (Linnaeus, 1766; Testudinidae). Brazilian J Biol 72(1): 153-161.
-
Regis KW, Meik JM (2017) Allometry of sexual size dimorphism in turtles: a comparison of mass and length data. PeerJ 5: e2914.
-
Webster M, Sheets HD (2010) A practical introduction to landmark-based geometric morphometrics. In: Hunt G, Alroy J (Eds.), Quantitative Methods in Paleobiology. Paleontological Society, Colorado, USA, pp: 163-188.
-
Zelditch ML, Swiderski DL, Sheets HD (2004) Geometric morphometrics for biologists: a primer. Elsevier Academic Press, Boston, USA, pp: 1-443.
-
Wyneken J, Godfrey MH, Bels V (2008) Biology of turtles. In: 1st (Edn.), CRC Press, New York, USA, pp: 1-404.
-
Martínez-Silvestre A, Hidalgo-Vila J, Pérez-Santigosa N, Díaz-Paniagua C (2015) Galápago de Florida – Trachemys scripta (Schoepff, 1792). In: Salvador A, Marco A (Eds.), Enciclopedia Virtual de Los Vertebrados Españoles. Museo Nacional de Ciencias Naturales, Madrid, Spain, pp: 1-41.
-
Ministerio de Agricultura Alimentacion y Medio Ambiente (2010) Catálogo Español de especies exóticas invasoras.
-
Macchi S, Balzarini LLM, Scali S, Martinoli A, Tosi G, et al. (2008) Assessing shell asymmetry in Trachemys scripta using geometric morphometrics. In: 7o Congresso Nazionale. Societas Herpetologica Italica, Oristano, Italy, pp: 333-337.
-
Parés-Casanova PM (2019) Adaptative directional asymmetric shape in Testudo hermanni hermanni Gmelin, 1789 (Reptilia: Testudines: Testudinidae). Herpetol Notes 12: 743-747.
-
Rohlf FJ (2015) The tps series of software. Hystrix Ital J Mammal 26(1): 9-12.
-
Bookstein FL (1991) Morphometric Tools for Landmark Data Geometry and Biology. Cambridge University Press, England, UK, pp: 1-456.
-
Klingenberg CP, Barluenga M, Meyer A (2002) Shape analysis of symmetric structures: Quantifying variation among individuals and asymmetry. Evolution 56(10): 1909-1920.
-
Laia RC, Pinto MP, Menezes VA, Rocha CFD (2015) Asymmetry in Reptiles: What Do We Know So Far. Springer Sci Rev 3(1): 13-26.
-
Klingenberg CP (2011) MorphoJ: An integrated software package for geometric morphometrics. Mol Ecol Resour 11(2): 353-357.
-
Hammer O, Harper DAT, Ryan PD (2001) PAST v. 2.17c. Palaeontol Electron 4(1): 1-229.
-
Anzelmo M, Sardi ML, Barbeito-Andrés J, Pucciarelli HM (2012) Alometrías ontogénicas y dimorfismo sexual facial en dos poblaciones humanas modernas. Rev Argentina Antropol Biol 14(1): 89-100.
-
Bidau CJ, Taffarel A, Castillo ER (2016) Breaking the rule: multiple patterns of scaling of sexual size dimorphism with body size in orthopteroid insects. Rev la Soc Entomológica Argentina 75(1-2): 11-36.
-
Carter AJR, Osborne E, Houle D (2009) Heritability of Directional Asymmetry in Drosophila melanogaster. Int J Evol Biol 2009: 1-7.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework