Preliminary Antecedents on the Selected Temperature (Tsel) of Rhinella spinulosa Wiegmann 1834 (Anura: Bufonidae) from the Andean Environments of Chile
The ambient temperature is a decisive factor in many activities of poikilothermic organisms, since it influences the correct development of their vital processes, which implies the existence of an optimal temperature range. The present study describes selected temperatures (Tsel) of R. spinulosa in adult individuals. For this purpose, adults of this species were acclimatized at 10°C and then subjected to a thermal gradient was used in the laboratory. The mean Tsel in the studied individuals was 23.38±2.65°C, with a range from 17.73°C to 28.93°C. Future surveys are suggested to determine the habitat thermal quality and thermoregulatory precision.
Introduction
The global distribution and abundance of amphibians is affected by environmental temperature [1, 2]. Temperature has been considered as a selective factor influencing the survival, growth and spread of these organisms [3]. However, in spite of being ectotherms, amphibians have been able to adapt to and inhabit a wide spectrum of thermal climates, including high latitudes, deserts and mountains, characterized by extreme temperatures [4]. Even so, changes in ambient temperature resulting from global warming can shorten the time frame for biological activities such as foraging and reproduction [5, 6, 7] causing the animals to spend more time in their shelters [8, 9].
To reach their preferred body temperatures, organisms can strategically move between different microhabitats, alter their activity times, adjust posture and body orientation, or take advantage of retreat sites [10, 11, 12, 13, 14]. Most amphibians regulate temperature by selection of microhabitats during the day and night, thus maintaining an optimum temperature (To) range [15]. Amphibians in mountainous areas optimize the body temperature (Tb) by exposing themselves to the sun, process called heliothermy, or by contact with the substrate, process called tigmothermy [16, 17, 18, 19], which allows them to obtain a body temperature similar to that of the environment, that is, to maintain a thermoconformist behavior [3]. However, the time spent on this behavior will depend on an individual’s ability to tolerate water loss [20, 21] and the necessity of being on a humid substrate to avoid desiccation [22, 23].
Natural environmental gradients strongly influence the variation patterns of species, giving rise to great phenotypic diversity in populations, as ambient conditions vary among sites or in controlled experimental conditions under gradients of a particular factor [24, 25]. Comparison of populations exposed to different environments allows us to investigate the extent to which selective pressures lead to adaptive responses and to identify the geographic scale in which these phenomena occur [26].
The experiments called common garden are used to determine whether geographically separated populations respond similarly to warming [27]. This helps to understand whether these responses are produced by local adaptation or by phenotypic plasticity [28].
One way of assessing the restrictions imposed by thermoregulation under natural conditions is to determine the temperatures that organisms voluntarily use for thermoregulation in ideal, unrestricted conditions. These temperatures are called selected body temperatures or Tsel and are usually established under laboratory conditions using a thermal gradient [29, 30]. The selection of this temperature may vary over time due to the high plasticity of amphibians in terms of Tb; their ability to acclimate allows them to move between thermal gradient extremes [31].
Mass and body size are variables that significantly affect individual thermoregulation [32]. Larger individuals have greater thermal inertia or resistance to temperature changes and heat and cool more slowly than smaller individuals, because they have less exposed surface area [33].
Rhinella spinulosa Wiegmann 1834 is an amphibian that is present in Bolivia, Peru, Argentina and Chile and inhabits heterogeneous thermal environments; in Chile it is found at latitudes from 18º to 41º S [34], but it is absent between 25° and 31° S [35]; it thus has a discontinuous distribution on the western slope of the Andes Range. This species inhabits areas near streams, lakes and wetlands at altitudes from 1000 to 4600 m and has been the subject of numerous studies, showing geographic variation in morphological, ecological, genetic and physiological traits [36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46]. It has a strong geographic structure and low levels of gene flow determined by low vagility and physiological limitations [41, 42].
Rhinella spinulosa is more active during warmer periods of the year [47]. It has physiological tolerance to water and temperature stress, which allows it to live at higher elevations [18, 37, 48]. Thermal studies indicate that R. spinulosa uses a variety of strategies to control body temperature [16, 17, 18]. Adults exhibit heliothermic behavior during the morning, which also helps them to improve digestion of prey caught the night before [49]. During the rest of the day, the species has tigmothermic behavior, using damp and shaded sites that reduce the loss of water by evaporation [17]. Larvae and postmetamorphics are active diurnally, while adults are nocturnal, staying hidden under rocks during the day [50]. Adults are more terrestrial than aquatic [47], unlike the postmetamorphics who prefer to stay close to water bodies [45].
The present study aims to determine temperature preferences (Tsel) in R. spinulosa species and establish differences between localities. These results are complementary to those published by the authors in Alveal, et al. [51] where the thermal tolerance of the species was analyzed. Alveal, et al. [51] determined that the species tolerates a wide range of temperatures, so it is considered eurythermic. In addition, significant differences were observed between the northern locality versus the central and southern ones in terms of CTmax, establishing that individuals from the north tolerate higher temperatures. Both, Tsel and thermal tolerance are generally accepted as central to characterize the thermal biology of ectothermic organisms [52].
Materials and Methods
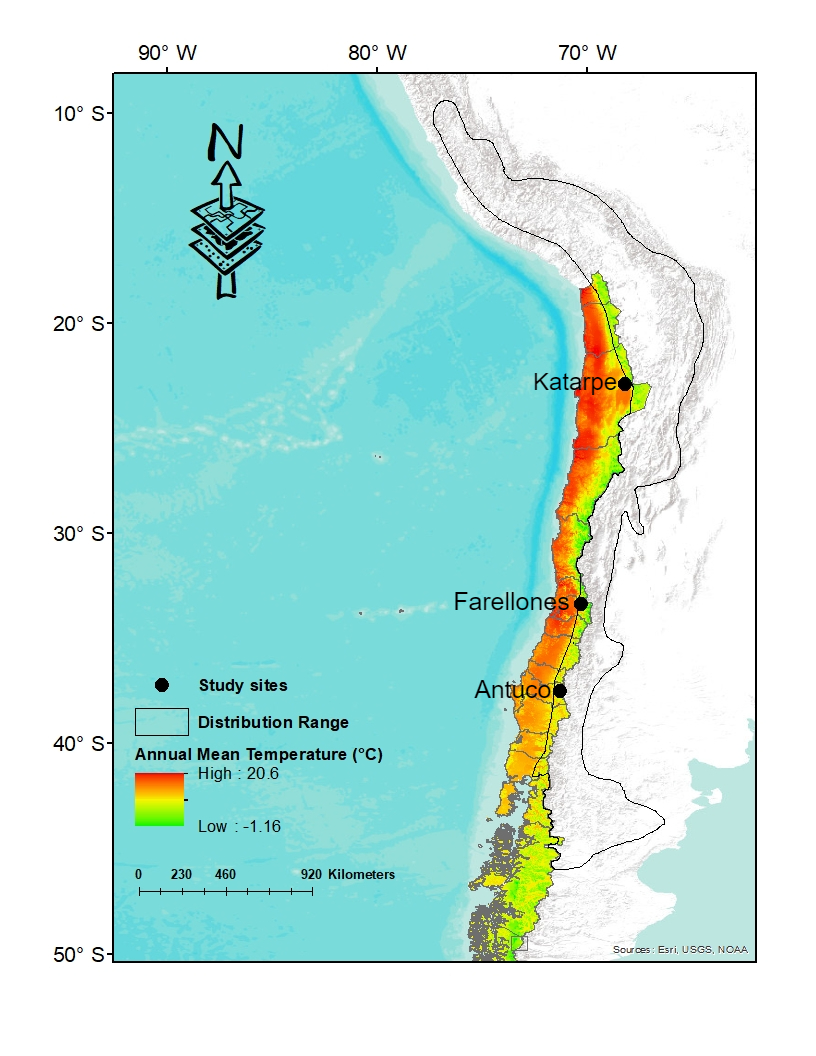
During the spring and summer of 2012 and 2013, three localities on the western slope of the Andes range were visited. These sites are representative of different climatic zones throughout of the range latitudinal of the species in Chile: Katarpe, Antofagasta region: 22°53´22.26´´S-68°12´53.73´´W, 2466 m, Farellones, Metropolitan region: 33º21´27´´S- 70º18´14´´W, 2392 m and Antuco, Bio Bio region: 37°28´32´´S-71°19´10´´W, 1409 m (Figure 1), with an annual average temperature of 12.7°C, 7.5°C and 5.9°C, respectively. In these areas the annual mean temperature decreases in relation to latitude (www.worldclim.org) which implies a different thermal environment in each population.
Captures were made manually using the “Protocol for the control of infectious diseases in amphibians during field surveys” [53]. A total of 19 males adult individuals were used for the experiment, three from the north, four from the center and 12 from the south. The individuals were maintained in the Laboratorio de Ecofisiología y Conservación de Herpetozoos at the Universidad de Concepción in terrariums (40 x 42 x 35 cm) and provided with conditions similar to those of their habitats, maintaining humidity with water containers to prevent dehydration. Individuals were subjected to a temperature of acclimatization of 10 °C for a period of two weeks, which corresponds approximately to the average environmental temperature of the three localities under study. A photoperiod regime of 10H light and 14H dark was used, which resembles the photoperiod of its activity period. They were fed ad libitum with earthworms (Lumbricus terrestris Linnaeeus 1758) and larvae of Chilecomadia moorei Silva 1915 and Tenebrio molitor Linnaeus 1758. Individuals were marked with elastomers in the ventral zone and intermembrane of hind limbs for identification, according to the Northwest Marine Technology protocol [54]. Prior to each experiment, the snout-vent length (SVL, mm) was measured with a digital caliper with 0.01 mm precision, and mass (g) using a digital balance with 0.01 g precision.
To determine the Tsel in the laboratory, an artificial temperature gradient was used. The system of thermal preference used consisted of a heat-sealed metal structure 195 cm long, 60 cm wide and 56 cm high; the internal space was divided into four lanes of about 15 cm each using a porous metal mesh. The base was made of transparent acrylic, perforated and covered with gravel. As the level of dehydration of individual anurans affects the performance [55], a constant moisture system was provided underneath which maintained the ambient humidity at approximately
60%. The gradient cover consisted of a transparent acrylic surface attached to a photoperiod system. The thermal gradient heated at one end and cooled at the other, providing a range of temperature in air and soil. The mean temperature range of the system was 9.27°C (± 2.09) to 38.63°C (± 2.40), increasing by approximately 3°C every 10 cm (Figure 2). The laboratory thermal gradient allowed the animals access to a wide range of environmental temperatures without being affected by other variables such as predators, prey and a number of other factors that could limit the field [56].
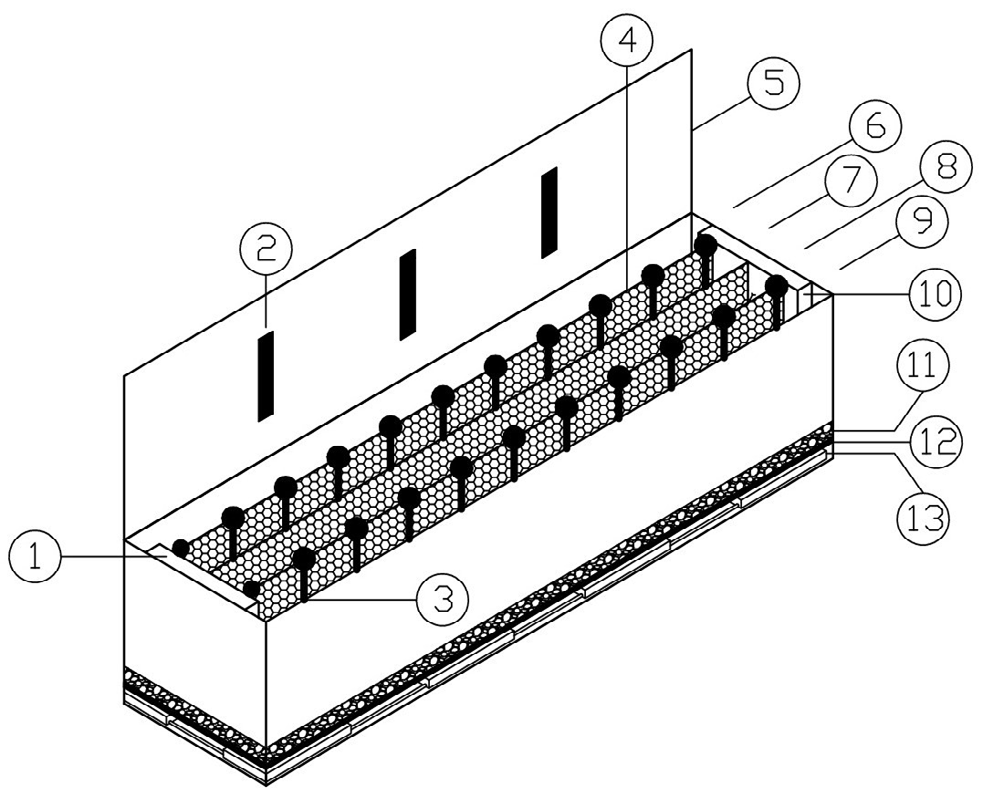
Figure 2: Thermal gradient. (1) Heat Source, (2) Photoperiod System, (3) Thermometer, (4) Lane Separator Mesh, (5) Gradient Cap, (6), (7), (8) and (9) Lanes, (10) Cooling Source, (11) Gravel Substrate, (12) Perforated Acrylic Base, (13) Moisture System. The mean temperature range of the system was 9.27°C (±2.09) to 38.63°C (± 2.40), increasing by approximately 3°C every 10 cm.
Data collection was performed simultaneously for four individuals, one in each lane, where it was not possible for them to come into contact. The animals were placed in the center of each lane and Tb dorsal temperature with a Digital Infrarrojo Laser (380°c/°f) Fluke thermometer was recorded at the beginning and in three subsequent moments: 5, 6 and 7 hours after starting the experiment. The mean of these three values corresponds to the Tsel of the organisms [57]. By measuring with an infrared thermometer it was not necessary to handle the animal, which reduces stress and preserves natural behaviors and eliminates the risk of altering the subject’s Tb [56].
The statistical support was tested in Infostat version 2020e. Kruskal Wallis tests were performed to show significant differences in the parameters mass, size and Tsel among the three localities. A nonparametric ancova analysis was also used, using mass and size as Tsel covariates.
Results
Rhinella spinulosa size and body mass varied significantly among localities. Analysis by locality showed a significantly larger size of individuals from the central locality (northern: 71.97 ± 0.28 mm; central: 89.63 ± 4.50 mm, southern: 75.63 ± 4.01 mm; K-W: H=10.07, P=0.006), which also had higher body mass compared to the other sites analyzed (northern: 45.33 ± 7.77 g; central: 69.18 ± 6.75 g, southern: 55.15 ± 11.87 g. K-W: H=7.72, P=0.021).
The individuals from the three localities preferred Tsel above 23°C (Tsel northern: 23.87 ± 2.52°C, Tsel central: 23.16 ± 2.13°C, Tsel southern: 23.34 ± 2.88°C) (Figure 3); however, there were no significant differences between localities (K- W: H: 0.45; P: 0.80). Therefore, as there were no differences between the three locations, the mean Tsel of R. spinulosa for this study was 23.38 ± 2.65°C (range: 17.73°C-28.93°C). On the other hand, the non-parametric Ancova analysis established that there were no significant differences in Tsel using mass (F: 1.70; P: 0.19) and size (F: 2.47; P: 0.12) as covariates.

Discussion
The regulation of body temperature in ectotherms has an important impact on their physiological and behavioral processes [3]. The selected temperature (Tsel) is the body temperature at which physiological processes such as digestion, swimming, and locomotion are optimized [58, 59]. Tsel can be modified in response to environmental parameters and/or physiological state so understanding the results of experiments with amphibians in thermal gradients is crucial for proper inference in studies of evolution, ecological climate change, niche modeling, and emerging diseases, among others [60, 61].
In our study, R. spinulosa shows to select a wide range of temperatures when subjected to a thermal gradient, which is consistent with its eurythermic behavior, since it tolerates critical maximum temperatures above 36°C and critical minimum temperatures below 0°C [50, 51]. This antecedents confirm that it is a thermogeneralist species, a characteristic that allows the species to survive in adverse microclimatic conditions [51].
Tsel studies in the genus Rhinella for the adult stage show a temperature range fluctuating between 21°C to 28°C [52, 60, 61, 62, 63, 64] so our results are within the range described for the genus.
For R. spinulosa, previous works only address Tsel in larval and postmetamorphic stages, which show a Tsel around 25°C [38, 50, 65]. The results of the present study are consistent with these Tsel showing a difference of 2°C less, which could be attributed to the nocturnal habits of adults.
The effect of activity periods on Tsel is an interesting aspect to address in thermal studies that could explain why adults subjected to a wide thermal supply select temperatures higher than the ambient temperature of their respective habitats. Anguilletta, et al. state that anurans in cold environments do not necessarily select these same temperatures in experimental gradients, so we can hypothesize that the optimal field temperature for processes such as locomotion in R. spinulosa is lower than Tsel in the laboratory. Furthermore, it has been established that in the absence of thermoregulatory opportunities (as might occur during the night), animals experience thermoconformational behavior, allowing time to be spent on other activities such as foraging, reproduction and shelter-seeking for daily torpor [62, 66]. However, for other processes, such as digestion, adults of R. spinulosa exhibit heliothermic behavior during the morning that helps them to improve the digestion of prey captured the previous night [49].
In a temperature gradient, individuals have greater thermal options to thermoregulate and there are no other variables that may hinder their preference such as lack of operative temperatures, risk of predation, water balance, among others [63, 66, 67]. Therefore, their Tsel will adjust to the temperature at which physiological performance is optimal [49], which allows them to compensate for thermal conditions in captivity, selecting temperatures similar to those they would select in shelters in their respective habitats.
These toads are active during the night and quickly equilibrated with the ambient temperature; however, during daytime rest, they bury or hide in volcanic rock shelters (Personal Obs.). In addition to this, since the experimentation was during the day, the Tsel would correspond to that of the resting period, so the individuals thermoregulated behaviorally selecting higher temperatures than those corresponding to the ambient temperatures in the field, which allows them to maintain a high metabolic rate during their resting period, a situation that changes during their period of activity at night. This has already been described in other bufonids, such as Anaxyrus fowleri under natural conditions [68]. These toads are active during the night and maintain a Tb similar to Ta, but during daytime rest, they select and maintain a body temperature around 30°C, in order to maintain a high metabolic rate. Thus, it can be established that these animals thermoregulate behaviorally during their resting hours [68].
In our field studies, it has been observed that during the active season the toads spend the daylight hours in shelters made up of volcanic rock burrows and that they do not explore microhabitats with higher thermal quality, such as areas open to the sun, a situation that also occurs in other species of the genus [64]. It is presumed that these shelters maintain stable temperatures throughout the daily cycle of the species and that night temperatures there are close to Tsel, allowing them to reach an optimal temperature after foraging.
Our results show the need for future studies to contrast experimental results with values obtained in field conditions, such as evaluating thermal precision and thermoregulatory efficiency taking into account the habitat thermal quality.
Although the three sampling localities are widely separated from each other [35, 46], the mean Tsel is similar in the three localities analyzed, without evidencing significant differences, which would indicate that individuals exhibit a conservative Tsel and thus rely mainly on behavioral thermoregulation to cope with cold and variable temperatures, since within the gradient, they tend to avoid low temperatures.
The degree to which physiological traits are phylogenetically conserved has been described primarily in reptiles, which demonstrate that lizards inhabiting different thermal environments have similar body temperatures due to behavioral thermoregulation which helps to reduce the strength of natural selection and reduce rates of trait evolution [69, 70, 71, 72].
However, without further background, it is not possible to confirm this response in our study. On the other hand, previous studies in R. spinulosa that analyzed local adaptation to temperature indicate that those originating from a highly variable environment show a broad transcriptional response to temperature change, thus possessing greater plasticity and tolerance than populations from homogeneous habitats [46].
This is how the selection of a temperature that is optimal for the organism may vary over time, as amphibians are organisms with high plasticity in terms of body temperature, whose acclimatization capacity allows them to move between thermal extremes [31] and Tsel should not necessarily be considered as coadapted with the optimal temperature and replicate field body temperature [73, 60].
Finally, because the amphibians have a permeable skin and both aquatic and terrestrial life stages make them susceptible to changes in temperature, studies related to thermal physiology in species such as R. spinulosa contribute to the understanding of how amphibian species will be affected by climate change [74], which also allows for more accurate predictions of species distribution models [75]. According to Blaustein, et al. [76], species distribution models should be strengthened with laboratory and field experiments, as understanding the causal mechanisms that limit species distribution is likely to improve our ability to anticipate changes in their range in response to climate change [77].
Acknowledgements
The authors wish to thank Lafayette Eaton, for translating the manuscript. Becas Doctorado Nacional 2019-21191272, ANID. We also thank Marcela Vidal, Claudio Correa, Angelo Sandoval, Evelyn Cortés, Pabla Hernández, Cristian Sáez and Juan Leiva.
References
-
Andrewartha H, Birch L (1954) The distribution and abundance of animals. The University of Chicago Press, USA, 520: 01-782.
-
Dunson WA, Travis J (1991) The Role of Abiotic Factors in Community Organization. The American Naturalist 138(5): 1067-1091.
-
Angilletta M (2009) Thermal adaptation a theoretical and empirical synthesis. Oxford University press, USA, pp: 304.
-
Angilletta M, Niewiarowski P, Navas C (2002) The evolution of thermal physiology in ectotherms. Journal of Thermal Biology 27: 249-268.
-
Dunham AE (1993) Population responses to environmental change: physiologically structured models, operative environments, and population dynamics. In: Kareiva PM, Kingsolver JG, et al. (Eds.), Biotic interactions and global.. Sunderland, MA, Sinauer Associates pp: 95-119.
-
Huey RB (1982) Temperature, physiology, and the ecology of reptiles. In Gans, C., Pough, F. H. (Eds.) Biology of the Reptilia. Academic Press. USA, pp: 25-91.
-
Sinervo B, Miles DM, Martínez Méndez N, Lara-Resendiz R, Méndez-De-La-Cruz F (2011) Response to comment on erosion of lizard diversity by climate change and altered thermal niches. Science, 332(6029): 537.
-
Cowles R, Bogert C (1944) A preliminary study of the thermal requirements of desert reptiles. Bulletin of the American Museum of Natural History, Dirección Meteorológica de Chile 83: 265-296.
-
Sinervo B, Lara-Reséndiz RA, Miles DB, Lovich JE, Ennen JR, et al. (2017) Climate change and collapsing thermal niches of Mexican endemic reptiles. White paper for the environmental working group of the UC-Mexico initiative, University of California.
-
Bakken GS (1989) Arboreal perch properties and the operative temperature experienced by small animals. Ecology 70(4): 922-930.
-
Huey R, Peterson CR, Arnold SR, Porter W (1989) Hot rocks and not-so-hot rocks: retreat-site selection by garter snakes and its thermal consequences. Ecology 70(4): 931-944.
-
Hertz PE (1992) Temperature Regulation in Puerto Rican Anolis Lizards: A Field Test Using Null Hypotheses. Ecology 73(4): 1405-1417.
-
Kearney M, Shine R, Porter W (2009) The potential for behavioral thermoregulation to buffer “cold-blooded” animals against climate warming. Proceedings of the National Academy of Sciences of the United States of America 106(10): 3835-3840.
-
Gunderson AR, Leal M (2016) A conceptual framework for understanding thermal constraints on ectotherm activity with implications for predicting responses to global change. Ecology Letters 19(2): 111-120.
-
Brattstrom BH (1963) A preliminary review of thermal requirements of Amphibians. Ecology 44(2): 238-255.
-
Pearson OP, Bradford DF (1976) Thermoregulation of lizards and toads at high altitudes in Peru. Copeia 1976(1): 155-170.
-
Sinsch U (1989) Behavioural thermoregulation of the Andean toad (Bufo spinulosus) at high altitudes. Oecologia 80(1): 32-38.
-
Lambrinos J, Kleier C (2003) Thermoregulation of juvenile Andean toads (Bufo spinulosus) at 4300 m. Journal of Thermal Biology 28(1): 15-19.
-
Sanabria E, Quiroga L, Acosta J (2007) Odontophrynus occidentalis (NCN). Body temperature. Herpetological Review 38: 188-189.
-
Lillywhite HB, Licht P, Chelgren P (1973) The role of behavioral thermoregulation in the growth energetics of the toad, Bufo boreas. Ecology 54(2): 375-383.
-
Sanabria E, Quiroga L, Acosta J (2003) Relación entre la temperatura corporal de adultos de Bufo arenarum (Anura: Bufonidae) y variables ambientales en un humedal de San Juan, Argentina. Multequina 12(2): 49- 53.
-
Tracy CR (1976) A model of the dynamic exchanges of water and energy between a terrestrial amphibian and its environment. Ecological Monographs 46(3): 293-326.
-
Titon B, Navas CA, Jim J, Gomes FR (2010) Water balance and locomotor performance in three species of neotropical toads that differ in geographical distribution. Comparative Biochemistry and Physiology 156(1): 129- 135.
-
Scheiner M (1993) Genetics and evolution of phenotipic plasticity. Annual Review of Ecology, Evolution, and Systematics 24: 35-68.
-
Pigliucci M, Murren CJ, Schlichting CD (2006) Phenotypic plasticity and evolution by genetic assimilation. Journal of Experimental Biology 209(12): 2362-2367.
-
Gómez Mestre I, Tejedo M (2003) Local adaptation of an anuran amphibian to osmotically stressful environments. Evolution 57(8): 1889-1899.
-
Pelini ShL, Diamond S, Mac Lean H, Ellison AE, Gotelli N, et al. (2012) Common garden experiments reveal uncommon responses across temperatures, locations, and species of ants. Ecology and Evolution 2(12): 3009- 3015.
-
Johnston I, Bennett A (1996) Animals and temperature: phenotypic and evolutionary adaptation. Society of Experimental Biology. Seminar Series. Cambridge University Press, Cambridge, USA, 59: 1-419
-
Pough FH, Gans C (1982) The vocabulary of reptilian thermoregulation. In: Gans C, Pough FH (Eds.), Biology of Reptilia, Academic Press, UK, 12: 17-23.
-
Licht P, Dawson WR, Shoemaker VH (1966) Heat resistance of some Australian lizards. Copeia, 1966(2): 162-169.
-
Brattstrom B, Lawrence P (1962) The rate of thermal acclimation in anuran amphibians. Physiological Zoology 35(2): 148-156.
-
Stevenson R (1985) The relative importance of behavioral and physiological adjustments controlling for body temperature in terrestrial ectotherms. American Naturalist, The University of Chicago Press, 126(3): 362- 386.
-
Hillman S, Withers P, Drewes R, Hillyard S (2009) Ecological and environmental physiology of amphibians. Oxford University Press, USA, pp: 464.
-
Correa C, Cisternas J, Correa-Solís M (2011) Lista comentada de las especies de anfibios de Chile (Amphibia: Anura). Boletín de Biodiversidad de Chile 6: 1-21.
-
Salinas H (2013) Patrones de diversidad genealógica en el anuro Rhinella spinulosa; conservación intraespecífica de sus linajes y posibles refugios pleistocénicos. Tesis de Magister, Universidad de Chile.
-
Veloso A, Navarro J (1988) Lista sistemática y distribución geográfica de anfibios y reptiles de Chile. Bollettino del Museo Regionale di Scienze Naturali, Torino 6: 481-539.
-
Ruiz G, Rosenmann M, Veloso A (1989) Altitudinal distribution and blood values in the toad, Bufo spinulosus Wiegmann. Comparative Biochemistry and Physiology, Chile 94(4): 643-646.
-
Benavides L, Gonzalo A (2003) Biología térmica de Bufo spinulosus: efecto de la temperatura sobre el desarrollo larval, una comparación intraespecífica. Doctoral Thesis, Universidad de Chile, Santiago, Chile.
-
Méndez M, Soto ER, Correa C, Veloso A, Vergara E, et al. (2004) Diferenciación morfológica y genética entre poblaciones chilenas de Bufo spinulosus (Anura: Bufonidae). Journal of Thermal Biology 77: 559-567.
-
Benavides G, Veloso A, Jimenez P, Mendez M (2005) Assimilation efficiency in Bufo spinulosus tadpoles (Anura: Bufonidae): effects of temperature, diet quality and geographic origin. Revista Chilena de Historia Natural 78: 295-302.
-
Jara FG, Simpson V, Úbeda C (2006) Bufo spinulosus papillosus (NCN). Tadpole behavior. Herpetological Review 37: 201.
-
Méndez M, Correa-Solís M (2009) Divergence in morphometric and life history traits in two thermally contrasting Andean populations of Rhinella spinulosa (Anura: Bufonidae). Journal of Thermal Biology 34(7): 342-347.
-
Correa C, Pastenes L, Sallaberry M, Veloso A, Méndez M (2010) Phylogeography of Rhinella spinulosa (Anura: Bufonidae) in northern Chile. Amphibia-Reptilia 31: 85- 96.
-
Navas C, Úbeda C, Logares R, Jara F (2010) Thermal tolerances in tadpoles of three species of Patagonian anurans. South American Journal of Herpetology 5(2): 89-96.
-
Gallardo C, Hernández H, Diniz Filho J, Palma E, Méndez M (2011) Relationship between the genetic structure of the Andean toad Rhinella spinulosa (Anura: Bufonidae) and the northern Chile landscape (21°- 24° S). Revista Chilena de Historia Natural 84(3): 391-406.
-
Pastenes L, Valdivieso C, Di Genova A, Travisany D, Hart A, et al. (2017) Global gene expression analysis provides insight into local adaptation to geothermal streams in tadpoles of the Andean toad Rhinella spinulosa. Scientific Reports.
-
Cei J (1962) Batracios de Chile. Ediciones Universidad de Chile, Santiago, Chile.
-
Espinoza RE, Quinteros S (2008) A hot knot of toads: aggregation provides thermal benefits to metamorphic Andean toads. Journal of Thermal Biology 33(2): 67-75.
-
Sinsch U (1991) Análisis radio-telemetrico de la regulación térmica del sapo Andino, Bufo spinulosus. Boletín de Lima 73: 65-73.
-
Sanabria E, Rodríguez C, Vergara C, Ontivero E, Banchig M, et al. (2015) Thermal ecology of the post-metamorphic Andean toad (Rhinella spinulosa) at elevation in the monte desert, Argentina. Journal of Thermal Biology 52: 52-57.
-
Alveal N, Díaz Páez H, Ortiz JC (2016) Thermal tolerance in the Andean toad Rhinella spinulosa (Anura: Bufonidae) at three sites located along a latitudinal gradient in Chile. Journal of Thermal Biology 60: 237-245.
-
Oliveira-Anderson RC, Parelli R, Vieira D (2018) Seasonal variation in the thermal biology of a terrestrial toad, Rhinella icterica (Bufonidae), from the Brazilian Atlantic Forest. Journal of Thermal Biology 74: 77-83.
-
Lobos G, Vidal M, Labra MA, Correa C, Rabanal F, et al. (2011) Protocolo para el control de enfermedades infecciosas en Anfibios durante estudios de campo. Red Chilena de Herpetología.
-
Northwest Marine Technology (2008) Visible Implant Elastomer Tag Project Manual.
-
Moore FR, Gatten JR (1989) Locomotor performance of hydrated, dehydrated and osmotically stressed anuran amphibians. Herpetologica 45(1): 101-110.
-
Taylor EN, Diele Viegas LM, Gangloff EJ, Hall JM, Halpern B, et al. (2020) The thermal ecology and physiology of reptiles and amphibians: A user’s guide. J Exp Zool 335(1): 13- 44.
-
Castañeda L, Lardies M, Bozinovic F (2004) Adaptive latitudinal shifts in thermal traits of a terrestrial isopod. Evolutionary ecology Research 6: 570-593.
-
Wells D (2007) The ecology and behavior of amphibians. The University of Chicago Press pp: 1146.
-
Navas C, Gomes F, Carvalho J (2008) Review: thermal relationship and exercise physiology in anuran amphibians: integration and evolutionary implications. Comparative Biochemistry and Physiology 151(3): 344- 362.
-
Clemente AC, Senzano LM, Gavira RSB, Andrade DV (2020) Feeding alters the preferred body temperature of Cururu toads, Rhinella diptycha (Anura, Bufonidae). Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 249: 110771.
-
Navas CA, Gouveia SF, Solano-IguaránJJ, Vidal MA, Bacigalupe LD (2021) Amphibian responses in experimental thermal gradients: Concepts and limits for inference. Comp Biochem Physiol B Biochem 254: 110576.
-
Sanabria E, Quiroga L (2011) Thermal parameters changes in males of Rhinella arenarum (Anura:Bufonidae) related to reproductive periods. Revista de Biología Tropical 59 (1): 347-353.
-
Sanabria E, Quiroga L, Martino A (2011) Seasonal changes in the thermoregulatory strategies of Rhinella arenarum in the Monte desert, Argentina. Journal of Thermal Biology 36(1): 23-28.
-
Noronha-de-Souza CR, Bovo RP, Gargaglioni LH, Andrade DV, BícegoKC (2015) Thermal biology of the toad Rhinella schneideri in a seminatural environment in southeastern Brazil, Temperature 2(4): 554-562.
-
Pastenes L, Valdivieso C, Di Genova A, Travisany D, Hart A, et al. (2017) Global gene expression analysis provides insight into local adaptation to geothermal streams in tadpoles of the Andean toad Rhinella spinulosa. Scientific Reports 7(1): 1966.
-
Huey R, Slatkin M (1976) Cost and benefits of lizard thermoregulation. The Quarterly Review of Biology 51(3): 363384.
-
Tracy CR, Christian KA, O’Connor MP, Tracy CR (1993) Behavioral thermoregulation by Bufo americanus: the importance of the hydric environment. Herpetologica 49(3): 375-382.
-
Forget Klein E, Green DM (2021) Toads use the subsurface thermal gradient for temperature regulation underground. Journal of Thermal Biology 99: 102956.
-
Cowles R, Bogert C (1944) A preliminary study of the thermal requirements of desert reptiles. Bulletin of the American Museum of Natural History, Dirección Meteorológica de Chile 83(5): 265-296.
-
Bodensteiner B, Warner D, Iverson J, Milne-Zelman C, Mitchell T. et al. (2019) Geographic variation in thermal sensitivity of early life traits in a widespread reptile. Ecology and Evolution 9(5): 2791-2802.
-
Huey RB, Hertz PE, Sinervo B (2003) Behavioral drive versus behavioral inertia in evolution: a null model approach. The American Naturalist 161(3): 357-366.
-
Hutchison VH, Dupré RK (1992) Thermoregulation. In Environmental Physiology of the Amphibians. In: Feder ME, Burggren WW (Eds.), University of Chicago Press, Chicago, Illinois, USA, pp: 472.
-
Piasečná K, Pončová A, Tejedo M, Gvoždík L (2015) Thermoregulatory strategies in an aquatic ectotherm from thermally-constrained habitats: An evaluation of current approaches. Journal of Thermal Biolog 52: 97- 107.
-
Oyamaguchi HM, Vo P, Grewal K, Do R, Erwin E, et al. (2017) Thermal sensitivity of a Neotropical amphibian (Engystomops pustulosus) and its vulnerability to climate change. Biotropica 50(2): 326-337.
-
Winter M, Fiedler W, Hochachka WM, Koehncke A, Meiri S, et al. (2016) Patterns and biases in climate change research on amphibians and reptiles: a systematic review. R Soc open sci 3.
-
Blaustein AR, Walls SC, Bancroft BA, Lawler JJ, Searle KL, et al. (2010) Direct and Indirect Effects of Climate Change on Amphibian Populations. Diversity 2(2): 281- 313.
-
Enriquez‐Urzelai U, Kearney MR, Nicieza AG, Tingley R (2019) Integrating mechanistic and correlative niche models to unravel range‐limiting processes in a temperate amphibian. Global Change Biology, 25(8): 2633-2647.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework