The Finding and Description of A Diverse Population of the Southern Cestode Isoglaridacris agminis Williams and Rogers, 1972 (Caryophyllaidae) from Lake Chub Sucker Erimyzon sucetta (Lacépède, 1802) in a Northern Land-Locked Lake in Wisconsin, with a Special Reference to its Disjunct Post-Glacial Distribution in North America
Specimens of Isoglaridacris agminis Williams and Rogers, 1972 (Caryophyllidae) were first described from the lake chubsucker Erimyzon sucetta (Lacépède, 1802) in a small tributary of the Tallapoosa River at NE central Alabama. We have collected and examined I. agminis from the same host species in the land-locked Silver Lake, SE Wisconsin. We update our first and only 1986 account of this Wisconsin population, with new observations and measurements, and compare with those in the original and other descriptions. The morphology of I. agminis is revisited using light microscopy revealing new features. Emphasis is placed on the morphological variability of its unusual scolex and reproductive structures with variable ovarian shapes throughout development using many microscopic images. Measurements of 17 specimens and the re-examination of 15 more specimens from Silver Lake provided an opportunity to review the taxonomy and description of the species, distinguish locality differences, and correct and update other statements and concepts by other observers. The disconnected distribution of worms from the same host species in separate waters of various states is attributed to the disjunct post-glacial distribution of their primary hosts.
Omar M Amin* and Nataliya Yu Rubtsova
of their primary hosts.
Keywords: Isoglaridacris agminis; Zoogeography; Fishes
Introduction
Mackiewicz [1] reviewed the morphology, anatomy, development, zoogeography, and systematics of the Caryophyllidea van Beneden, 1893 (Cestoidea) and indicated the existence of 89 species and 37 genera in these monozoic cestodes. Mackiewicz [2] provided perspectives on the evolution, biology and zoogeography of 128 species in 45 genera of caryophyllids from the Nearctic, Neotropical, Palearctic, Ethiopian, Oriental, and Australian regions. Mackiewicz [3] later recognized about 150 species in 41 genera worldwide (except for the Neotropical Region). Of these, 14 genera and 90 species belonging to 3 families were described from the Indo-Malayan region from Bagridae, Heteropneustidae, Schilbeidae and Siluridae, cypriniformes, and cobitid fishes [3]. Ash, et al. [4, 5] invalidated a large number of inadequately described species from clariid and catfishes in India and neighboring countries. Most recently, however, Scholz, et al. [6] and Scholz and Kuchta [7] recognized only 117 species and 46 genera in order Caryophyllidea Carus, 1863 parasitizing teleost fish (Cypriniformes, Siluriformes, and some catfish). Chubb [8] reviewed the seasonal ecology of the Caryophyllidea in the various climate zones of the world.
Amin [9, 10, 11, 12] recognized 15 species in 8 families of caryophyllid cestodes mostly from suckers (Catostomidae) in lake and river systems in Arizona and Wisconsin. These included Isoglaridacris agminis Williams and Rogers, 1972. It is important to note here that the order Caryophyllidea includes 4 families (Balanotaeniidae, Capingentidae, Caryophyllaeidae, Lytocestidae) discussed by Mackiewicz [1, 3] and reviewed in Amin and Rubtsova [13].
In Wisconsin, Amin [12] gave an account of I. agminis from E. sucetta in Silver Lake, and distinguished between it and that of Williams and Rogers’ original description [14]. Williams, et al. [15] followed up with additional measurements of 20 more specimens, corrected and accounted for a larger collection of 46 additional specimens of I. agminis from a new host species, Minytrema malanopis (Rafinesque, 1820) in additional collection sites on 3 other river systems in eastern Alabama. The only other known records from recognizable collecting sites in Arkansas in the western creek chubsucker Erimyzon claviformis (Girard, 1856) were recorded by AcAllister, et al. [16]. Elsewhere, I. agminis was reported from the creek chubsucker Erimyzon oblongus Mitchill, 1814 in Wake County, North Carolina by Grimes, et al. [17] who did not name the collecting site but noted 56-77 (69) testes in gravid specimens. Similarly, Mackiewicz [18] reported 60 specimens of I. agminis from E. sucetta in unspecified geographical locations in Alabama, Florida, Mississippi, and North Carolina, and expressed apprehension about its generic assignment for inconsequential reasons. Notwithstanding reports from unknown collecting sites in North Carolina, Florida, Mississippi, and Alabama [17, 18], the only recognizable collecting sites are known from Alabama [14, 15] and Wisconsin (this paper) from unconnected waters, albeit from the same hosts. I. agminis appears to have highly specific host specificity and distribution and its geographical distribution appears to be limited and of disjunct nature.
Materials, Collections, and Methods
Collections
Seasonal collections of 139 specimens of I agminis from 28 of 116 (24%) examined lake chubsuckers, E. sucetta (mean 1.2, maximum 38), in the land-locked Silver Lake, Kenosha County, SE Wisconsin (42°32′58″N,88°10′9″W) were made between 1976 and 1984. In the nearby Tichigan Lake, a relatively large eutrophic impoundment on the Fox River which is a tributary of the Mississippi River in Racine County (42°48′20″N,88°13′00″W), only 2 lake chubsuckers were captured that harbored no parasites.
Methods
After capture and transfer to the laboratory in coolers on ice, the fish were dissected within a few hours after capture. Upon recovery, worms were placed at once in warm 70% ethanol then processed and stained with Semichon’s carmine. Worms were then processed in ascending concentrations of ethanol for dehydration reaching 100%. Worms were ultimately cleared in oil of winter green before whole mounting in Canada balsam. Measurements are in mm or micrometers as noted in Table 1; the range is followed by the mean values between parentheses. Width measurements represent maximum width. Microscope images were created using 10X and 40X objective lenses of a BH2 light Olympus microscope (Olympus Optical Co., Osachi- shibamiya, Okaya, Nagano, Japan) attached to an AmScope 1000 video camera (United Scope LLC, DBA AmScope, Irvine, California), linked to an ASUS lab top equipped with HDMI high-definition multimedia interface system (Taiwan- USA, Fremont, California). Images from the microscope were transferred from the lab top to a USB and stored for subsequent processing on a computer. We found microscope images to be considerably more informative than schematic line drawings as they depict the natural appearance of anatomical structures.
| Locality | Silver Lake, Wisconsin | Uphapee Creek, Macon County & Coosa, Chattahoochee, Tallapoosa rivers, Alabama |
|---|---|---|
| Specimens measured | 17+ 15 | 10 + 20 (22 + 46 studied) |
| Hosts | Erimyzon sucetta | Erimyzon sucetta, Minytrema malanopis |
| Authority | Amin (1986) and this paper | Williams & Rogers (1972), Williams (1975) |
| Characters | ||
| Body length (mm) | 3.00-13.20 (6.28) | 3.77-7.73 (5.52) |
| Body width at gonopore (mm) | 0.40-0.77 (0.51) | 0.47-0.78 (0.57) |
| Scolex | Barely wedge-shaped, ovoid or lantern- shaped with indistinguishable loculi | Wedge shaped with 3 pairs of shallow or deeper loculi |
| Scolex length (mm) | 0.37-1.07 (0.68) | --- |
| Scolex width (mm) | 0.32-0.75 (0.52) | --- |
| Neck | Long, slender, tapering at both ends | Moderate-long |
| Neck length (mm) | 0.50-2.60 (1.29) | --- |
| % Neck length to worm length | 21% | --- |
| Narrowest Neck width (mm) | 0.17-0.40 (0.27) | --- |
| Vitellaria | From posterior neck to anterior ovary | From scolex to ovary + post-ovarian vitellaria |
| Vitellaria length (µm) | 125-200 (148) | 34-66 (54) corrected to 85-182 (116) |
| Vitellaria diameter (µm) | 73-135 (104) | 32-43 (34) corrected to 36-143 (70) |
| First vitellarium to anterior tip | 0.68-2.80 (1.56) | 0.63-2.23 (1.16) |
| % of worm length | 11-38 (26%) | 21% |
| Testes | Usually in 1 dorsal & 1 ventral medial rows, in smaller field than vitellaria | In 1 dorsal and 1 ventral layer occasionally in irregular or random rows; posterior from anterior vitellaria to cirrus sac |
| Testes number | 17-68 (39) | 28-40 |
| Testis length (µm) | 94-218 (156) | 101-221 (166) |
| Testis diameter (µm) | 80-192 (124) | 91-188 (134) |
| First testis to ant. tip (mm) | 0.80-4.20 (2.02) | 1.42-2.93 (1.99) |
| % of worm length | 32% | 36% |
| Post-gonopore distance (mm) | 0.84-1.52 (1.07) | 0.68-1.29 (0.90) |
| % of worm length | 10-17 (15)% | 17% |
| Cirrus sac | Large, oblong-round, between anterior ends of ovarian arms | Oblong |
| Cirrus sac diameter (µm) | Round, 140-198 (171) | 120-162 (138) X 85-138 (118) |
| Post-ovarian vitellaria number | 5-17 (9.7) | 4-11 clustered or in V-shape (Figs. 1, 4, 5) |
| Ovary shape | Lobate, H-shaped, arms’ length variable | Elongated, inverted A-shaped with open apex |
| Ovarian arms’ length (µm) | 350-840 (524) | 420-840 (580) |
| Distance between ovarian arms (µm) | 375-750 (499) | --- |
| Shape of posterior end | Bluntly pointed | Bluntly pointed (Figs. 4, 5) |
| Egg shape | Ovoid, operculate, smooth | Operculate with smooth shell |
| Egg length (µm) | 33-54 (44) | 34-66 (45) |
| Egg diameter (µm) | 25-38 (31) | 32-43 (34) |
Table 1: Comparative morphometrics of Isoglaridacris agminis from Erimyzon sucetta in Silver Lake Wisconsin and Uphapee Creek, Al
Bolded figures indicate numbers markedly different among relevant populations. Table 1: Comparative morphometrics of Isoglaridacris agminis from Erimyzon sucetta in Silver Lake Wisconsin and Uphapee Creek, Alabama.
Results
Amin [12] gave morphological notes of I. agminis from the Wisconsin population collected from lake chubsuckers E. sucetta captured in Silver Lake, SE Wisconsin between 1976 and 1984. He [12] made initial taxonomic comparisons with specimens from other collections in Alabama and [14, 15]. We have restudied specimens in our personal collection. Measurements of about 17 specimens were made in 1967- 1968 [12] and new measurements and observations of 15 additional specimens from the original collection were recently made and added to the earlier measurements creating combined measurements included in Table 1. Amin [12] noted that the percent gravid worms of the originally reported 139 worms collected from E. sucetta examined from Silver Lake was 22 in the autumn, 84 in the spring and 100 in the summer suggesting a seasonal reproductive cycle. No juveniles were recovered in the autumn, and prevalence and intensity of infection progressively decreased in the spring and summer indicating accelerated development early in the infectious cycle and earlier loss of gravid worms in the spring [12]. During all seasons, I. agminis infected the post- gastric region of the gut. “The short-lived Neoechinorhynchus robertbaueri Amin, 1985 shared the same anterior intestinal locations with I. agminis during the spring; it is not present in its fish hosts during the autumn” [12, page 54].
We provide below a more elaborate revised description of our qualitative morphological findings and a detailed comparative morphometrical data (Table 1) high-lightening extreme variations from other descriptions by Williams and Rogers [14] and Williams [15].
Brief morphological Description of Adults of I. agminis from Wisconsin
Body small-medium with long neck averaging about one fifth worm length (Figures 6-14), Widest at gonopore (Figures 19, 20). Body usually dorso-ventrally flattened, covered with thin cuticle enclosing prominent cortical parenchyma and inner and weaker outer longitudinal muscles. Measurements and counts in Table 1. In adults, scolex barely wedge-shaped, usually ovoid or lantern-shaped, longer than wide, tapering anteriorly, cut by 6 usually barely noticeable loculi (Figures 6, 7-10, 13, 14). Younger specimens with anteriorly cone- shaped proboscis with indistinguishable loculi (Figures 1-3) as well as with underdeveloped testes and vitellaria, and immature ovary and uterus (Figures 4 & 5). Adults with seventeen to 68 large spheroidal testes usually in 1 dorsal and 1 ventral medial rows occasionally appearing in partially irregular or random rows (Figures 15- 17,19 & 20). Mature adults with testicular field extending from 32% of anterior worm tip to just anterior to cirrus sac. Paired osmoregulatory canals in testicular field. Testes occupying less space than vitellaria anteriorly and posteriorly (Figures 11,14-16). Cirrus sac large, oblong- round, near ventral surface at tip of anterior ovarian arms (Figure 25, arrow). Vitelline glands large, only slightly smaller than testes, ovoid-elongate, multi- layered in 2 lateral fields (Figures 21 & 22), extending from 11-38% of anterior worm tip to anterior end of ovarian arms. Vitellaria extend dorsally and ventrally about the edge of testicular field (Figures 21 & 22). Post-ovarian vitellaria always present, especially large, not continuous with pre- ovarian vitellaria, with 5-17 glands clustered or in V-shaped pattern (Figures 19,24,28-29). Male and female reproductive systems open into common gonopore with post gonopore distance of 10-17% of worm length. Vas deference detectable about middle of testicular field ventrally. Distinctly lobate ovary H-shaped in varying length and robustness (Figures 19,20,23-29). Ovarian commissure fanning at juncture with ovarian arms and tapering towards center (Figure 25). Vagina without seminal receptacle and running posteriorly to anterior aspect of ovarian commissure. Uterine coils usually not extending anterior to cirrus sac but posteriorly reaching to posterior tip of posterior ovarian arms (15-29). Posterior end bluntly conical (Figures 19,24,28 & 29) with slight terminal middle invagination where excretory vesicle and pore open tremino-ventrally. Eggs small, ovoid, thin- shelled, operculate (Figure 30).
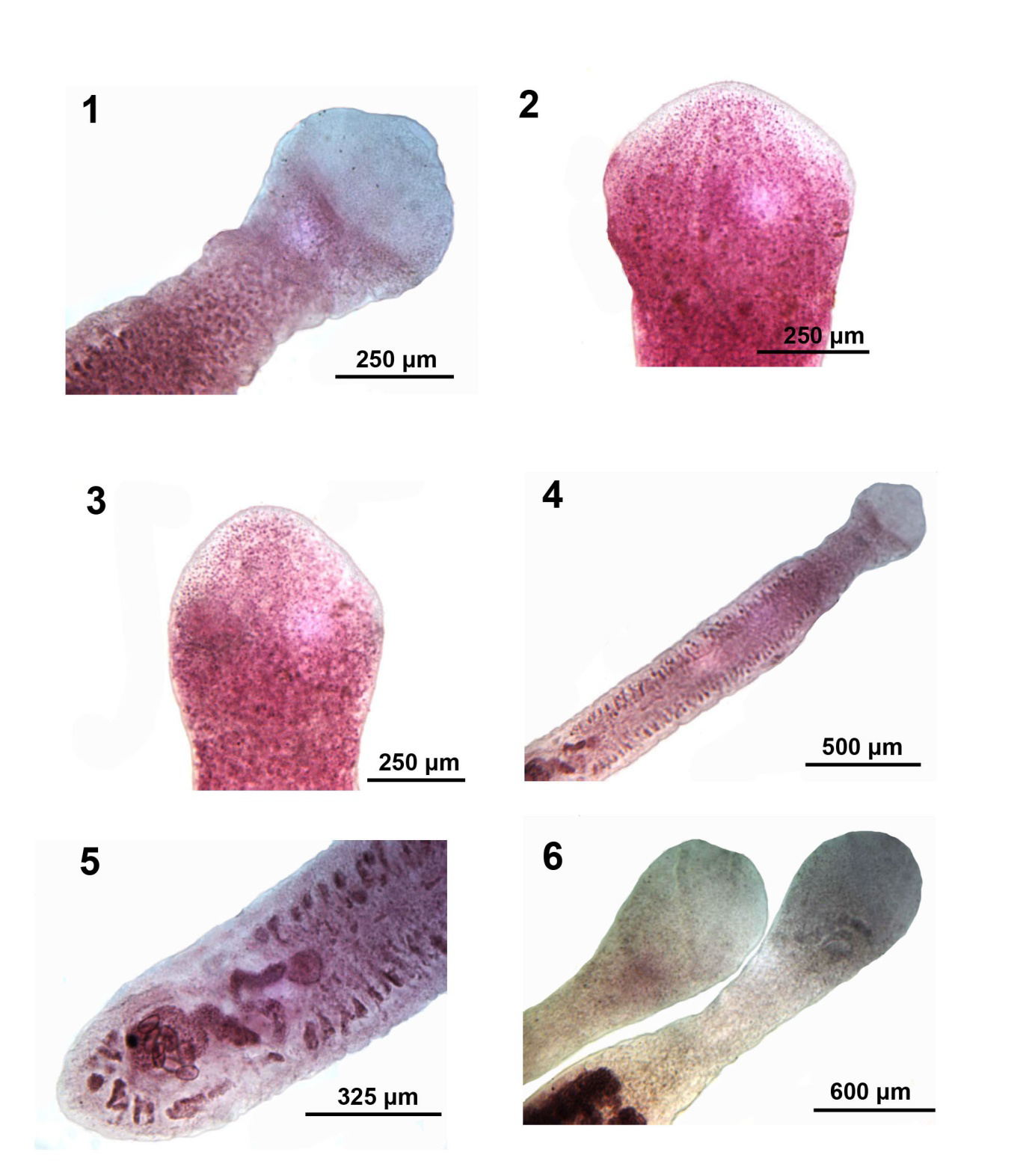
Figures 1-6: Microscope images of developing and maturing adults of Isoglaridacris agminis from Erimyzon sucetta in Silver Lake, Wisconsin. 1-3. The bluntly conical scolex of juveniles showing some longitudinal muscle fibers. 4, 5. The anterior to mid-section and posterior portion of one young maturing specimen showing underdeveloped vitellaria and incipient female reproductive system with barely developing uterus and eggs (Figure 5). 6. Developed scolexes of 2 older specimens showing their characteristic ovoid-balloon shape, undefined loculi and no terminal disc.
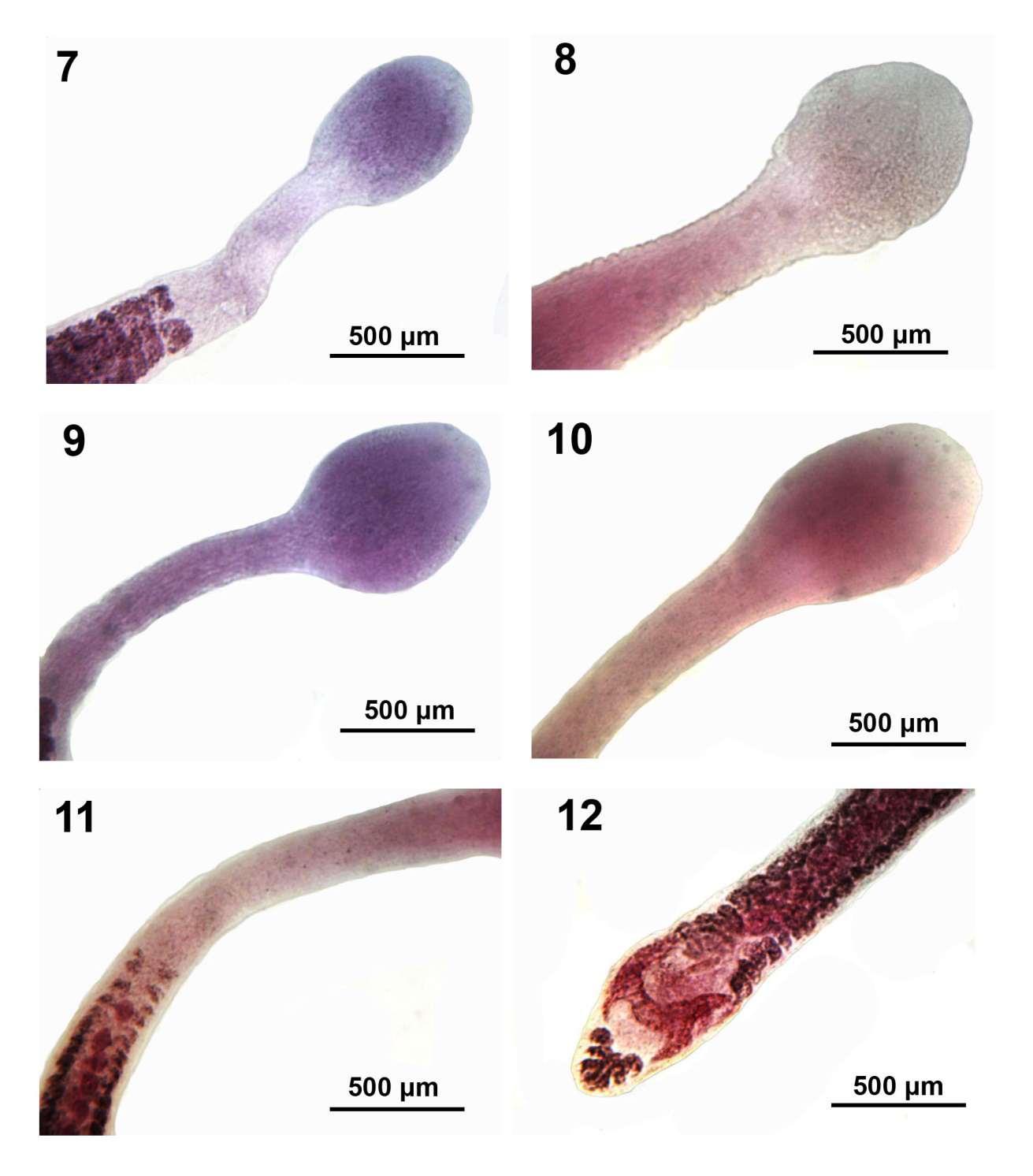
Figures 7-12: Microscope images of adults of Isoglaridacris agminis from Erimyzon sucetta in Silver Lake, Wisconsin. 7-9. Variations in the rounded-oblong shape of the scolexes of 3 specimens typical of the Wisconsin material. 10-12. The scolex, neck and posterior portion of the same specimen. Note the more posterior position of the anterior-most testis compared to the anterior vitellaria (Fig. 11) and the shape of the ovary (Fig. 12).
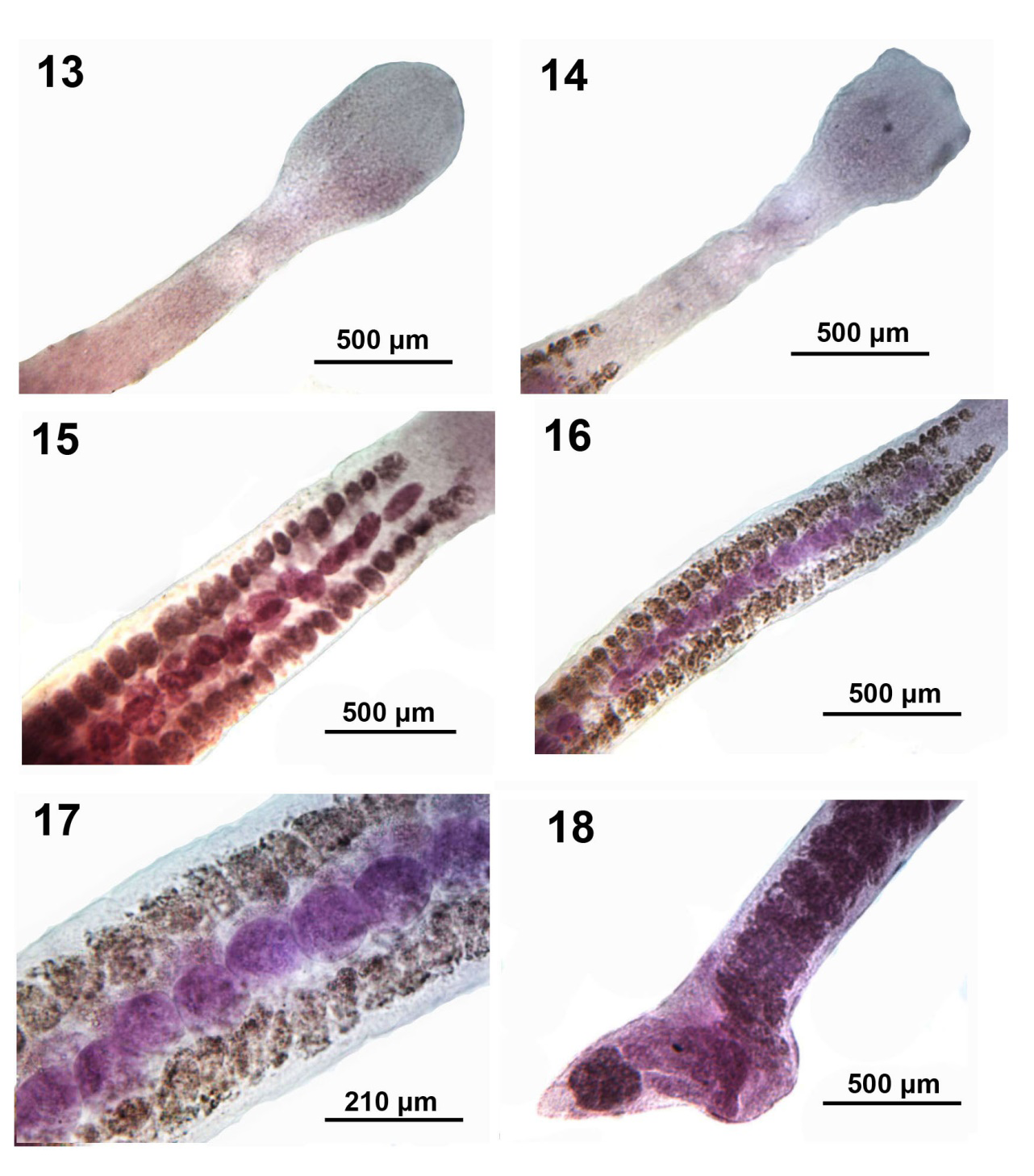
Figures 13-18: Microscope images of adults of Isoglaridacris agminis from Erimyzon sucetta in Silver Lake, Wisconsin. 13, 14. Additional variations in the shape of the scolexes of 2 more worms. Note the lantern-like scolex pattern in Fig. 14. 15-17. A ventral view of the variations in the organization of the single ventral medial row of testes. Note the large size of the vitelline glands especially in the more magnified Fig. 17 compared to the testes. 18. A lateral view of the posterior portion of a breeding worm with the ventral expansion of the cirrus pouch and adjacent area.
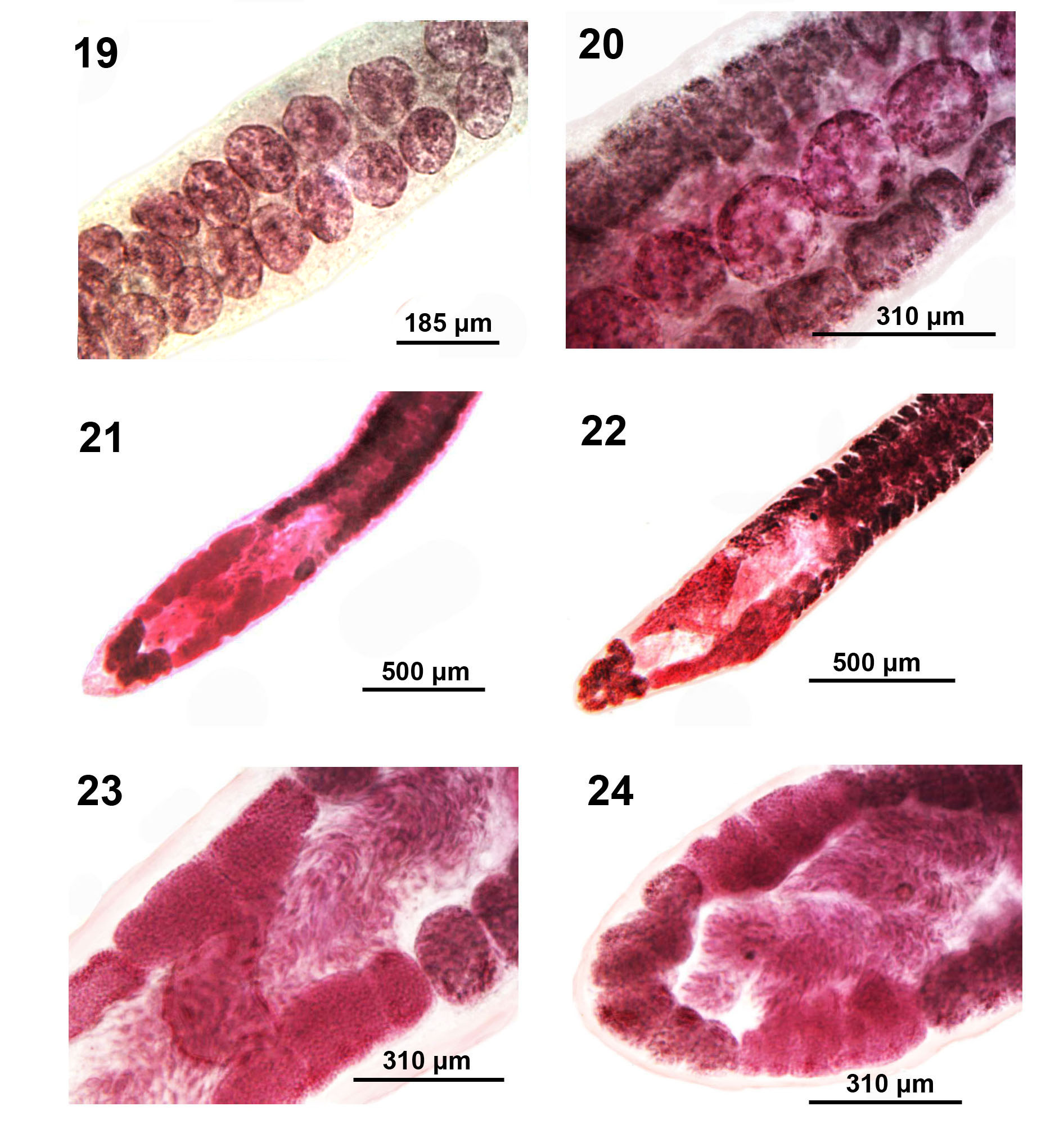
Figures 19-24: Microscope images of adults of Isoglaridacris agminis from Erimyzon sucetta in Silver Lake, Wisconsin. 19. A lateral view showing and dorsal and ventral single rows of testes. 20. A higher magnification of a ventral aspect of one row of testes showing their size and shape compared to those of the vitellaria. 21-24. Variations in the size, shape, and robustness of the ovarian arms. Note also the robustness of the V-shaped post-ovarian vitellaria (Fig. 21) compared to the cluster-shaped post-ovarian vitellaria (Fig. 22).
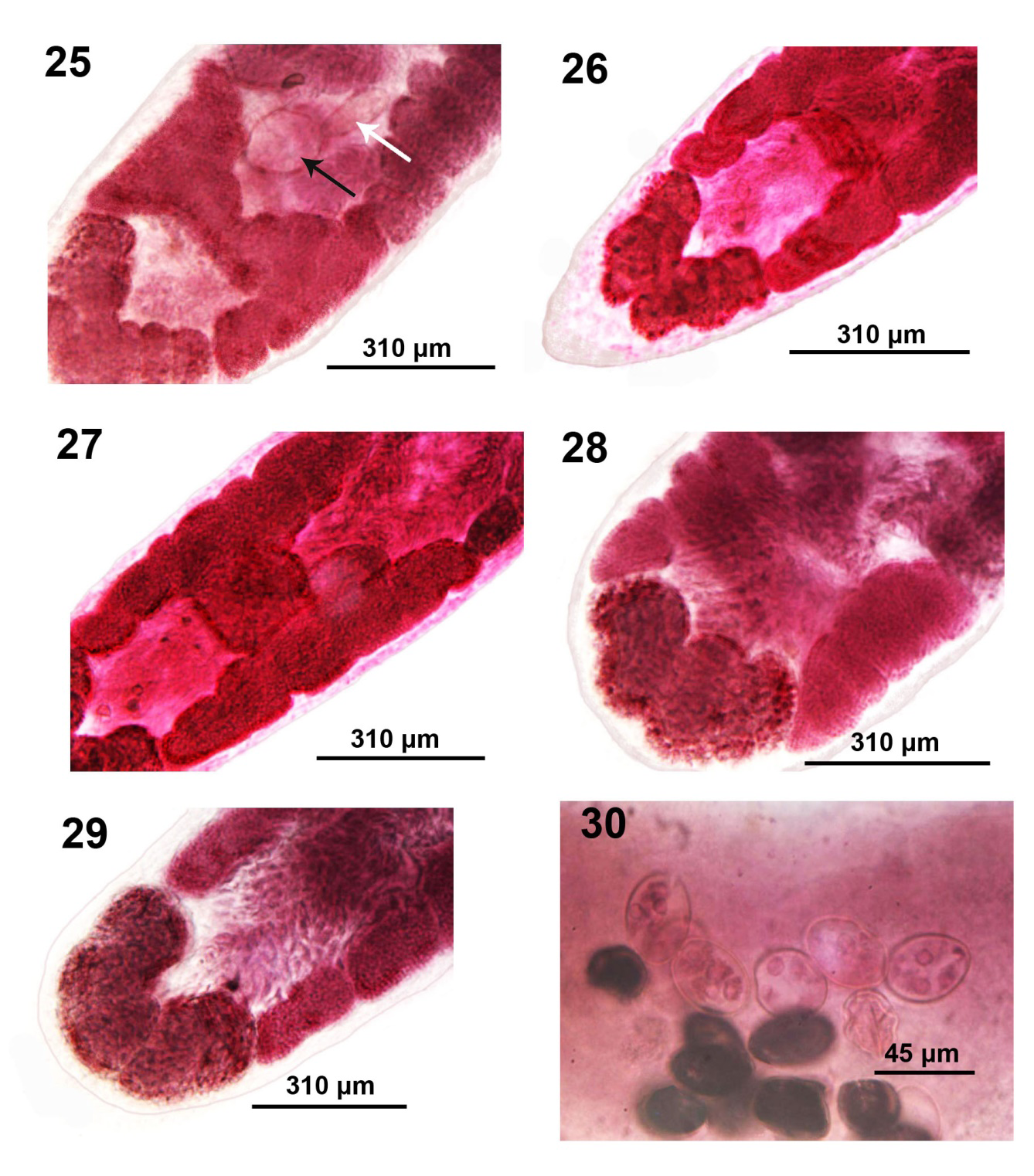
Figures 25-30: Microscope images of adults of Isoglaridacris agminis from Erimyzon sucetta in Silver Lake, Wisconsin. 25-29. More variations in shape and size of the ovary. Note the cirrus sac (black arrow), the external seminal vesicle (white arrow), and the commissure fanning out at juncture with the ovarian arms (Fig. 25). Also note the extreme shortness and broadness of the ovary arms and the extreme size of the few post-ovarian vitellaria in Figs. 28, 29. 30. Eggs; note the few yolk cells.
Taxonomic Summary
- Type and current host: Lake chubsucker Erimyzon sucetta Lacépède
- Other hosts: Creek chubsucker E. oblongus, spotted sucker Minytrema melanops (Rafinesque).
- Type locality: Uphapee Creek near Tuskegee, Macon County (32°48’N, 85°70’W), Alabama.
- Other identifiable localities: Chattahoochee, Coosa, and Tallapoosa River systems, Alabama [15], Caddo River and Big Fork Creek, Arkansas [16], and Silver Lake, SE Wisconsin (this paper).
- Other states from which specimens have been collected but no names of water bodies were given: Florida, North Carolina, Mississippi [17].
- Deposited specimens from our Wisconsin material: US National Museum Helminthological collections nos. 78653, 78654 & Milwaukee Public Museum coll. no. IZ985-01A.
Remarks
The original description of I. agminis from E. sucetta in Alabama [14] was brief but well documented. Williams [15] supplemented and corrected errors in the original description from another host species, M. melanops, also from Alabama. Only Grimes, et al. [16] mentioned that their gravid specimens from E. oblongus in unspecified waters in North Carolina had 56-77 (69) testes. Another descriptive account of that species by Amin [12] from E. sucetta in the disconnected waters of the Wisconsin land-locked Silver Lake produced a brief comparison between the Wisconsin and the Alabama populations. The only other morphological account of I. agminis and the most recent is that in McAllister, et al. [16] who tentatively identified a few specimens of I. agminis from the western creek chubsucker E. clavirormis in Arkansas. The Arkansas specimens were noted to be similar to those described by Williams and Rogers [14] but varied in the number of testes (67), shorter neck, and uterine loops reaching slightly anterior to the cirrus sac. We have observed the same variations in our Wisconsin specimens. McAllister, et al. [17] made no reference to variations in the scolex and provided no figures or illustrations of their specimens. We here, however, resolve doubts about the identity of the Arkansas specimens and propose transferring its tentative identity to a genuine one. We also provide herein a considerably more detailed and expanded comparison between these Alabama and Wisconsin populations (Table 1) supplemented by microscope images for the first time.
Adults of our specimens from Wisconsin were longer than the Alabama specimens from the same host and with more testes and larger vitelline glands and cirrus sac. The scolex in our specimens, unlike the wedge -shaped scolex of the original description; was hardly wedge shaped but mostly ovoid or less frequently lantern-shaped with barely detectable loculi (Figures 6-10,13,16). The neck was comparatively long, about 21% of worm length, the distribution of testes (in one dorsal and one ventral medial rows) and vitellaria, and the lobulated ovarian arms were comparable to those in the original description (Table 1). Other features presented in Table 1 that were not highlighted were comparable among the two populations. The shape, size of H-shaped ovarian arms and their robustness showed the most variation (Figures 19,20,23-29).
In his description of Isoglaridacris calentinei Mickiewicz 1974, Mackiewicz [18] made a special reference to I. agminis (misspelled as I. agaminis) in which he stated on page 147 that “After the examination of the holotype and a single paratype, as well as 60 specimens from the same host in Alabama, Florida, Mississippi, and North Carolina, I am unable to accept the generic allocation of I. agaminis, by Williams and Rogers (1972) because of scolex shape (often with a small disc) and ovary condition (posterior arms well separated forming a distinctly H-shaped ovary).” There was no disc, large or small, on the scolex of any of our specimens from Wisconsin and none was mentioned in the original description [14]. Mackiewicz [19] originally diagnosed the genus Isoglaridacris with an inverted A-shaped ovary although “an H-shaped one also occurred.” He then considered the inverted A-shaped one as the “normal” one. “However, descriptions of five other species indicate that the genus is characterized by having a high proportion of both ovarian types (joined and not joined posterior arms) in the same species” [18, page 146]. All Mackiewicz’s [18] seven line-drawings of I. calentinei displaying ovaries showing posterior ovarian arms not joining as in H-shaped ovaries, occasionally with posterior arms approaching. It appears then that the H-shaped ovary in I. agminis does not represent a serious departure from Mackiewicz’s generic diagnosis which could have been emended to accommodate this ovarian variability. Newly described species of Isoglaridacris appear to agree with our opinion. For example, in Isoglaridacris brevicollis Scholz and Pérez-Ponce de León, 2021 from Catostomus nebuliferus Garman in Mexico, the ovary was described as “inverted A-shaped, with long and narrow ovarian wings, (with) posterior wings approaching to each other (Fig. 2A, C) but never connected posteriorly” [20], in contrast to Mackiewicz’s [19] strict definition of the genus. In both Isoglaridacris chetekensis Williams, 1977 and Isoglaridacris wisconsinensis Williams, 1977 from Moxostoma macrolepidotum (LeSueur) and Hypentelium nigricans (LeSueur), respectively in Wisconsin, the ovaries were similarly described as “H-shaped, lobate; posterior ovarian arms converge medially but do not join” [21] (page 93, Figures 1, 2, 8).
Discussion
The generic assignment of I. agminis has not been challenged since Mackiewicz [17] stated his reservations prompting him not to include it in his key to 5 species of Isoglaridacris. We have noted above the reasons why we did not agree with Mackiewicz’s arguments. What remains to be solved is the geographical distribution of I. agminis. The only identified bodies of water where it is known to have been collected from are in the states of Alabama and Wisconsin, as well as in Arkansas.
(1) In Wisconsin, I. agminis is known only from E. sucetta in the land-locked Silver Lake in Kenosha County [12]. It remains an enigma how it got there. In addition, E. sucetta in the adjacent Tichigan Lake which is connected to The Fox River, a tributary of the Mississippi River in Wisconsin is not infected with I. agminis [12].
(2) In Alabama, I. agminis was described from E. sucetta in Uphapee Creek, in Macon County [14], a tributary of the Tallapoosa River which drains into the Alabama River, a major tributary of the Mobile River in southern Alabama. Later on, Williams [15] reported a larger collection of specimens from the spotted sucker M. melanops in the Chattahoochee, Coosa, and Tallapoosa River systems in central and eastern Alabama. The Coosa River, along with the Tallapoosa River, are tributaries of the Alabama River that flows southwards in the Mobile River which discharges in the Mobile Bay at the Gulf of Mexico [22]. The Chattahoochee River, forming the southern half of the Alabama and Georgia border, is a tributary of the Apalachicola River which flows southwards from Florida into the Apalachicola Bay in the Gulf of Mexico [23]. These waterways have been listed by Williams [15] but the specific collecting sites along these two independent River basins (the Mobile and the Chattahoochee basins) were not identified.
(3) In Arkansas, specimens were collected from the western creek chubsucker Erimyzon claviformis in the Caddo River at Caddo Gap, Montgomery County, Arkansas (34°23’ 55.0212’’N; 93°37’ 17.346’’W) and from Big Fork Creek, Polk County, Arkansas (34°28’ 5.109’’N; 93°56’ 52.1772’’W). The Caddo River is a tributary of the Ouachita River whose drainage basin lies in the upper Coastal Plain of Arkansas and Louisiana and in the alluvial valley of the Mississippi [24]. It is not connected to any larger river basin from which I. agminis has been collected and is not associated with the Alabama rivers reported by Williams and Rogers [14] or Williams [15].
(4) Other geographical records not identifying collecting sites in different waterways include site(s) in Wake County, North Carolina reported by Grimes, et al. [16]. There are about 100 streams in Wake County but the major bodies of water include Lake Crabtree, Crabtree Creek, Lake Johnson, the Neuse River, and portions of Falls Lake and Jordan Lake. There are no natural lakes in Wake County but there are several large reservoirs including Harris Lake on Buckhorn Creek and Falls [25]. None of these water bodies is connected to a larger River basin or can be associated with sites in Alabama further south from which I. agminis was reported.
(5) Additional geographical records not identifying collecting sites mentioned in Mackiewicz [18] include the states of Alabama, Florida, Mississippi, and North Carolina. Sites in Alabama from which infected fish were collected could have included the Tallapoosa River, the Alabama River or the Mobile River in southern Alabama reported earlier by Williams and Rogers [14] or in the Chattahoochee or Coosa River systems reported later by Williams [15] in central and eastern Alabama. Sites in Florida may have included portions of the Chattahoochee River which forms the southern half of Alabama and Georgia border, and a portion of the Florida and Georgia border [26]. Sites in Mississippi may have been associated with the Mobile River watershed from whose tributaries the Coosa and Tallapoosa rivers infected fish were collected [14, 15]. Sites in North Carolina may have been in Wake County from where Grimes, et al. [16] made his collection from unknown waters. In North Carolina, most major river systems drain eastward to discharge into the Atlantic Ocean and are not connected with any of the major inland river basins such as the Mississippi.
The North Carolina river basins include the Chow and Lower Roanoke, Pamlico and Neuse, Cape Fear, Yadkin-Pee Dee, Santee, French Broad, and New River drainage basins [25]. No infected fish are known to have been collected from any of these rivers.
Conclusion
Morphology
We conclude our treatment of I agminis from Wisconsin fishes with our acknowledgement of the morphological variability of 5 other Arizona and Wisconsin caryophyllid cestodes originally collected and reported by Amin. Other species in this series include Isoglaridacris hexacotyle (Linton, 1897) Mackiewicz, 1968 from Arizona, and Biacetabulum biloculoides Mackiewicz & McCrae, 1965, Glaridacris catostomi Cooper, 1920, Pseudoglaridacris confusus (Hunter, 1929) Oros, Uhrovič, Scholz, 2018, and Pseudoglaridacris laruei (Lamont, 1921) Oros, Uhrovič, Scholz, 2018 from Wisconsin. In all cases, we have reported the expanded range of qualitative variations in the morphology of the scolex and reproductive structures that varied with host species and/ or developmental stages as has been well documented for I. agminis. Scolexes in developing juveniles assume immature anteriorly tapering morphology usually lacking defined terminal disc or loculi different from the scolexes of adults reported in original descriptions. The degree of variations in the size or number of testes by host species or geographical locations varied in many species. In addition, we included among the pivotal features of taxonomic importance in our descriptions of all species such characters as the distance between the male and female gonopores, the earlier development of the testes compared to vitellaria, the lateral vs. the medullary fields of distribution of vitellaria, size of cirrus sac and eggs and whether the latter are operculated, the degree of development of inner vs. outer longitudinal muscles, the number of post-ovarian vitellaria, the distance between anterior-most vitellaria and scolex tip, the shape and makeup of the ovary (whether lobate or granular), and the anterior and posterior extensions of uterine coils. Some of these characters are of taxonomic importance at the generic level such as the shape makeup of ovarian tissue, the organization of the posterior ovarian arms, the distance between the male and female gonopores and whether they are in a common genital atrium. Measurements of some of these structures were not reported in many original descriptions and other descriptive accounts but are all included and tabulated for our reported Arizona and Wisconsin specimens.
Distribution
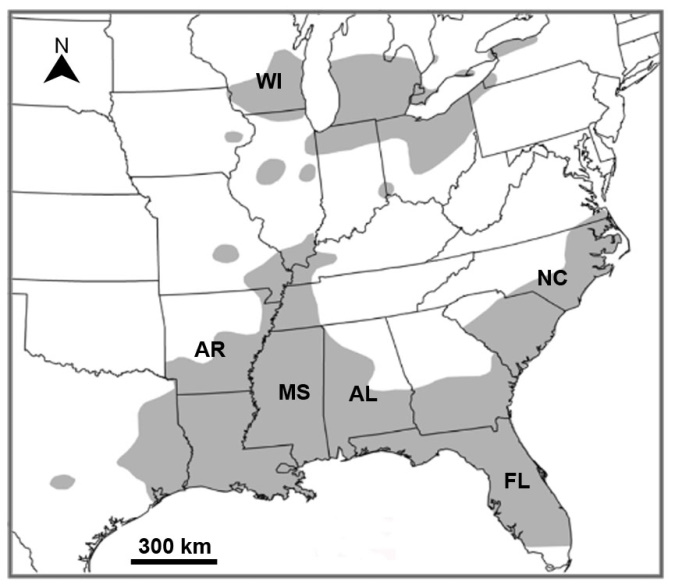
The enigmatic sporadic distribution of I. agminis in independent waters of Wisconsin (land-locked Silver Lake, Kenosha County), West and east Alabama (the Mobile River drainage and the Chattahoochee River), North Carolina, and Arkansas are hard to explain but they appear to be related to the disjunct distribution of its hosts. Isoglaridacris agminis appears to be restricted mostly to suckers of the genus Erimyzon Jordan, especially the lake chubsucker, E. sucetta, the type host. The distribution of E. sucetta in North America has been well documented by Mandrak, et al. [27]. Lake Chubsucker exhibits a disjunct distribution in North America (Fig. 31). It remains the “only representative of the genus …. in Canada (as it is) declining throughout most of its North American range” [27, page 19]. A southern element is centered on the Gulf States and extends northward from the Arkansas River through the Mississippi Valley to southern Illinois; east of the Mississippi River to the Atlantic Seaboard northward to southern Virginia; and west of the Mississippi River to eastern Texas. A northern element encompasses the southern Great Lakes drainage. Distribution is fragmented [27]. Trautman [28] hypothesized that this “fragmentation was the result of northeastern range expansion during the warm Hypsithermal Period (ca. 7000 to 5000 years before present), and subsequent range contraction and fragmentation during the wane of this Period” (page 5). This fragmentation left residual pockets of suckers in isolated streams along with their parasites including I. agminis. Fig. 31 shows the extension of E. sucetta into the SE third of Wisconsin including Kenosha County where the land-locked Silver Lake is located in the SE corner of the state. Central and western Wisconsin have never been glaciated and extensive 3-year parasitological data from NW Wisconsin by Fischthal [29] indicate that among 375 suckers of 6 species examined, E. sucetta was not found. A similar pattern appears to occur in other species of fish parasites. For instance, Amin [30] demonstrated that the present distribution of 3 species of acanthocephalans of the genus Acanthocephalus Koelreuter 1771 in North America, especially Acanthocephauls dirus (Van Cleave, 1931) in the post-glacial streams of SE Wisconsin “formed after the withdrawal of the Lake Michigan lobe of the Wisconsin ice sheet from the area between 15,000 and 12,000 B.P. (page 216).” Future research may produce new records of I. agminis from E. sucetta in states within its range where infections in chubsuckers have not been reported, i.e., Georgia, Louisiana, or Michigan [31].
Acknowledgments
This project was supported by an Institutional Grant from the Parasitology Center, Inc. (PCI), Scottsdale, Arizona.
Declarations
- Compliance with ethical standards
- Conflict of Interest. The authors declare no conflicts of interest or competing interests.
- Ethical Approval. The authors declare that they have observed all applicable ethical standards.
- Availability of Data. All presented and related data are available by contacting the senior author.
References
-
Mackiewicz JS (1972) Caryophyllidea (Cestoidea): a review. Exper Parasitol 31(3): 417-512.
-
Mackiewicz JS (1982) Caryophyllidea (Cestoidea): Perspect Parasitol 84: 397-417.
-
Mackiewicz JS (1994) Order Caryophyllidea van Beneden in Carus, 1863. In: Khalil LF, Jones A, et al. (Eds.), Keys to the cestode parasites of vertebrates. CAB Intern Wallingford UK, pp: 21-43.
-
Ash A, Scholz T, Oros M, Kar PK (2011a) Tapeworms (Cestoda: Caryophyllidea), parasites of _Clarias batrachus_ (Pisces: Siluriformes) in the Indomalayan region. J Parasitol 97: 435-459.
-
Ash A, Scholz T, Oros M, Levron C, Kar PK (2011b) Cestodes (Caryophyllidea) of the stinging catfish _Heteropneustes fossilis_ (Siluriformes: Heteropneustidae) from Asia. J Parasitol 97(5): 899-907.
-
Scholz T, Kuchta R, Oros M (2021) Tapeworms as pathogens of fish: A review. J Fish Dis 44(12): 1-18
-
Scholz T, Kuchta R (2017) A digest of fish tapeworms. Vie et milieu - Life and environment 67(2): 43-58.
-
Chubb JC (1982) Seasonal occurrence of helminths in freshwater fishes Part IV. Adult Cestoda, Nematoda and Acanthocephala. Adv parasitol 20: 1-292.
-
Amin OM (1968) Helminth fauna of Suckers (Catostomidae) of the Gila River System, Arizona. PhD Thes Ariz St Univ 202 pp (No 68-1644) Univ Microfilms. Ann Arbor, Michigan; Dissert Abstr 28: 3521.
-
Amin OM (1969) Helminth fauna of suckers (Catostomidae) of the Gila River System, Arizona. II. Five parasites from _Catostomus_ spp. Am Midland Nat 82: 429- 443.
-
Amin OM (1974) Intestinal helminths of the white sucker, _Catostomus commersoni_ (Lacépède), in SE Wisconsin. Proc Helminthol Soc Wash 41: 81-88.
-
Amin OM (1986) Caryophyllaiedae (Cestoda) from lake fishes in Wisconsin with a description of _Isoglaridacris multivitellaria_ sp. n. from _Erimyzon sucetta_ (Catostomidae). Pro Helminthol Soc Wash 53:48-58. Helminthol Soc Wash 53: 48-58.
-
Amin OM and Rubtsova NYu (2022) Reexamining _Pseudoglaridacris confusus_ (Hunter, 1929) nom. nov. (Cestoda: Caryophyllidae) from buffalo fish, _Ictiobus_ spp. in Arizona, USA. Int J Zoo Animal Biol 5(6): 000420.
-
Williams EH and Rogers WA (1972) _Isoglaridacris_ _agminis_ sp. n. (Cestoda: Caryophylleidae) from the lake chubsucker, _Erimyzon sucetta_ (Lacépède). J Parasitol 58: 1082-1084.
-
Williams EH (1975) Two new species of _Isoglaridacris_ (Cestoidea: Caryophylleidae) from Alabama. Trans Amer Micros Soc 94: 340-346.
-
McAllister CT, Choudhury A, Fayton TJ, Cloutman DG, Bursey CR, et al. (2021) Parasites of Western Creek Chubsucker _Erimyzon_ _claviformis_ (Cypriniformes: Catostomidae) from Arkansas and Oklahoma, U.S.A. Comp Parasitol 88(1): 113-129.
-
Grimes LR and Miller GC (1975) Caryophyllaeid cestodes in the creek chubsucker, _Erimyzon oblongus_ (Mitchill), in North Carolina. J Parasitol 61(5): 973-974.
-
Mackiewicz JS (1974) _Isoglaridacris calentinei_ n. sp. (Cestoidea, Caryophyllidea) from catostomid fish in western United States. Trans Amer Micros Soc 93: 143- 147.
-
Mackiewicz JS (1965) _Isoglaridacris bulbocirrus_ gen. et sp. n. (Cestoidea: Caryophyllaeidae) from _Catostomus_ _commersoni_ in North America. J Parasitol 51: 377-381.
-
Scholz T, Pérez-Ponce de León (2021) Caryophyllidean tapeworms (Cestoda), Nearctic parasites of fish in Mexico, including description of a new species of _Isoglaridacris_ and the first report of _Khawia japonensis_, an invasive parasite of common carp. Intern J Parasitol: Parasites & Wildlife 15: 70-78.
-
Williams DD (1977) _Isoglaridacris chetekensis_ sp. n. and _I. wisconsinensis_ sp. n. (Cestoda: Caryophyllaeidae) from Red Cedar River, Wisconsin catostomid fishes. Proc Helminth Soc Wash 44: 91-95.
-
Deutsch WG (2018) Alabama Rivers, A Celebration and Challenge. Florence, Ala: Mind Bridge Press.
-
Chattahoochee-Flint River Basin (2010) River Basin Center.
-
Anonymous (2017) Ouachita-Black River navigation.
-
List of rivers of North Carolina.
-
Anonymous (2010) Chattahoochee-Flint River Basin.
-
Mandrak NE, Cudmore B, Crossman EJ (2008) COSEWIC assessment and update status report on the lake chubsucker _Erimyzon sucetta_ in Canada. Committee on the Status of Endangered Wildlife in Canada, Ottawa, pp: 1-29.
-
Trautman MB (1981) The fishes of Ohio. Ohio State University Press, Columbus, pp: 683.
-
Fischthal JH (1953) Parasites of northwest Wisconsin fishes: summary and limnological relationships. Trans Wis Acad Sci 42: 83-108.
-
Amin OM (1985) Hosts and geographic distribution of _Acanthocephalus_ (Acanthocephala: Echinorhynchidae) from North American freshwater fishes, with a discussion of species relationships. Proc Helminthol Soc Wash 52: 210-220.
-
Page LM, Burr BM (1991) A field guide to freshwater fishes, North America; North of Mexico. Houghton Mifflin Co Boston, Massachusetts pp: 432.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework