Preliminary Exploration of the Ecological Habits of Odorrana lipuensis
Odorrana lipuensis occurs in an entirely dark karst cave in Lipu County, Guangxi Province, China. This paper discusses the ecological habits of the type locality of O. lipuensis, including the cave environment, daily activity rhythm, annual activity pattern, food, and predators. The results show that the life cycle of O. lipuensis is completed entirely in dark caves; cave temperature fluctuates around 19℃; there is no hibernation phenomenon, and it is active throughout the year. The daily activity rhythm does not differ between day and night, and breeding seasons range from March to June. Given the limited distribution range and small population size of O. lipuensis, it is recommended for inclusion in the national second-class protection animal list.
Introduction
Odorrana lipuensis lives in dark karst caves and is currently only distributed in one karst cave in Maling Town, Lipu County, Guangxi Province, China [1] (Figure 1), which is the type locality. Little is known about the distribution and ecological habits of O. lipuensis since its publication. Field surveys were conducted around the type locality between 2013 and 2022, but no new distribution points were found in China. However, Pham, et al. [2] identified a second distribution point of O. lipuensis in Ha Lang District, Cao Bang Province, Vietnam, located near the Vietnam-China border. In the type locality, a population of approximately 20 individuals was found living in completely dark karst caves, and their ecological habits are not well understood.
To improve our understanding of this species and enhance conservation efforts, we conducted preliminary research on its ecological habits from 2013 to 2022, and the results are presented below.
Materials and Methods
Study area and surveys: The type locality of O. lipuensis is Lipu County, Guangxi, China (110°26´ E, 24°38´ N; altitude: 182 m a.s.l.) and it inhabits dark karst caves. From Spring 2013 to September 2022, we conducted surveys of the caves where O. lipuensis resides. These surveys involved visual encounter methods to determine the presence and abundance of tadpoles, juveniles, and adult O. lipuensis, as well as searching for egg clutches on the rock walls, crevices, and ceilings of the caves. The organisms encountered during the survey were recorded, and the tadpole stage was determined using the Gosner method [3].
Monitoring Cave Temperature and Humidity: The Tp- 2200 temperature and humidity recorder was used to record data every two hours, starting from 18:51 on October 15, 2016, and ending at 2:51 on November 13, 2017, covering one natural year. The VICTOR (model: VC230A) was used to record water temperature.
Monitoring Activity Rhythm: The activity rhythm of the O. lipuensis was studied using the Bestguarder (SG-990V) infrared cameras, which were set to take pictures every 30 seconds continuously for 24 hours. The number of times the O. lipuensis appeared in front of the camera within each half- hour period was counted.
Analyzing Food Composition and Natural Enemies: The stomach contents of the O. lipuensis were dissected and observed to determine its food sources. Potential food sources and natural enemies found in the cave were investigated and recorded.
Results
In March, we observed the amplexus behavior of O. lipuensis and collected tadpoles at the 39th stage towards the end of March and the beginning of April. Subsequently, in June, tadpoles at the 43rd stage were observed (Figure 1). Our surveys did not yield any eggs, nor did we find any O. lipuensis outside the caves. However, we did observe the presence of Duttaphrynus melanostictus, Fejervarya multistriata, Hylarana guentheri, Microhyla butleri, Microhyla fissipes, and Rhacophorus mutus outside the caves, but no anuran was found within the caves.

Annual Temperature and Humidity Patterns
Over the course of a natural year, 4721 temperature points were recorded in the cave, with an average temperature of 19.01±0.46℃ (ranging from 18.25℃ to 19.75℃). The cave temperature remained stable throughout the year, and no changes in humidity were recorded due to a malfunctioning humidity sensor. The mechanical hygrometer indicated a humidity level of around 75% (GEMlead, model: TH101B). The water temperature was measured at 19.4℃ (Figure 2).
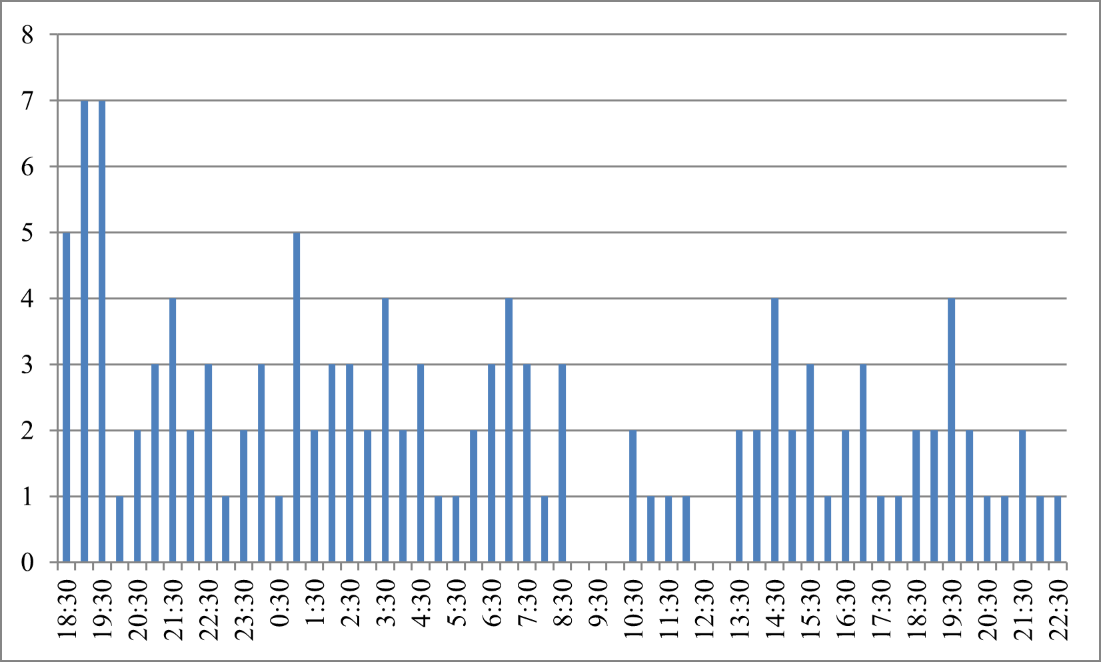
Daily Activity Rhythm
From January 14, 2016 at 18:37 to January 15, 2016 at 22:49, the infrared camera captured a total of 3383 photos, among which 125 photos of O. lipuensis were identified, indicating an occurrence rate of 3.7%. Figure 2 shows that O. lipuensis was active throughout the day with no discernible diurnal rhythm.
Food Composition and Natural Enemies
The stomach contents of two adult frogs were examined, but no identifiable food species were found under the microscope. However, Sinopoda sp., Triphosa sp., Diestrammena sp., and Glyphiulus sp. (Figure 3) were observed in the cave, with the highest encounter rate being Diestrammena sp. These animals could all potentially serve as food sources for O. lipuensis. During the field survey, no direct predators of O. lipuensis were observed, but Myotis chinensis and mammal footprints were present in the cave (Figure 3), suggesting that they could be potential natural enemies.

Discussion
Odorrana lipuensis is exclusively found in a dark karst cave at its type locality and completes its entire life cycle there. The cave has no light and a narrow entrance that can only accommodate one person. It is located 80 meters away from the underground river outlet and has a stable temperature that fluctuates between 19°C. A water pool of approximately 2 square meters is inside the cave, close to the wall, and has many crevices. The water temperature remains at around 19°C throughout the year, with water seeping out of the crevices during the rainy season. O. lipuensis is active all year round, with no hibernation period and no obvious diurnal or nocturnal activity rhythm. The breeding period of O. lipuensis lasts from about March to June. Observations were made on the amplexus behavior of O. lipuensis in March, and tadpoles at 39th stage were collected at the end of March and beginning of April. Tadpoles at 43rd stage were seen in June, as shown in Figure 1 [3]. Additionally, dissection of two female O. lipuensis in March revealed yellow eggs in their abdomen. While Odorrana liboensis and Odorrana wuchuanensis also inhabit karst caves [4, 5, 6], their breeding seasons differ. According to Luo, et al. [6], the living environment and habits of O. liboensis are very similar to those of O. lipuensis, such as no light, stable water temperature and they are sister groups in terms of phylogeny. However, the breeding season of O. liboensis is speculated to occur from late June to mid-August. Meanwhile, O. wuchuanensis has yellowish eggs in their abdomen in July and amplexus behavior is observed in August, leading to an estimated breeding period of July to August [5, 7]. This is later than the breeding period of O. lipuensis and O. liboensis. According to Zhao, et al. [8], the distribution of Odorrana is influenced by the average temperature in July, the annual average temperature, and annual rainfall. However, since the frogs live and complete their life cycle in caves where the environment is stable, external environmental factors do not affect their distribution. Further research is needed to investigate the difference in breeding seasons between O. lipuensis and O. liboensis.
Pham, et al. [2] found that O. lipuensis in Vietnam lives in secondary karst forests, where they were found on branches near the cave entrance, about 0.2-0.5 meters above the ground. However, the life cycle of O. lipuensis is completed entirely in the dark cave environment, as evidenced by observations of morphology and habits at various stages inside the cave. The differences in living environments between the type locality and the Vietnam habitat of O. lipuensis require further study.
Caves have a relatively closed environment, resulting in significantly lower species diversity compared to outside the caves. The analysis of the stomach contents of two O. lipuensis individuals showed almost no visible digested food items. Only a few species of organisms such as Sinopoda sp., Triphosa sp., Diestrammena sp., and Glyphiulus sp. (Figure 3) were observed in the cave with relatively low population numbers. Compared to non-cave species such as Odorrana graminea and Odorrana schmackeri, O. lipuensis is relatively smaller and thinner, which could be attributed to long-term food shortages. Human activities and not just bats and small mammals, also severely impact the survival of O. lipuensis. Evidence of man-made theft of stalactites and the use of bat guano as fertilizer was found in the cave. Moreover, locals often catch O. lipuensis for medicinal purposes, believing it can cure children’s fever, which is an erroneous belief that challenges their conservation. From 2013 to 2019, the O. lipuensis population in the type locality remained stable at around 20 individuals. However, during our September 2022 revisit, we discovered that the population had declined to less than 10 individuals, and the cave environment had been significantly damaged due to heavy soil disturbance. Outside the cave, we observed logging and charcoal burning. Due to O. lipuensis’s unique habitat and extremely small population, Guangxi has listed it as a key protected wildlife species [9]. We recommend the inclusion of O. lipuensis in the second level of national key protected wildlife species to facilitate its protection.
In addition to O. lipuensis, O. liboensis, and O. wuchuanensis, only a few species of frogs inhabit caves. These include Eleutherodactylus cooki [10], E. cundalli [11], Cycloramphus eleutherodactylus [12], and Rana iberica [13]. However, these species do not display troglomorphic characteristics such as degenerate eyes and depigmentation. Some species are occasionally found in caves for breeding or to avoid predators [13], while others exclusively reside in caves and complete their life cycle there. The adaptation mechanisms of these species to the dark karst cave environment require further research.
Funds: This work was supported by Guangxi Natural Science Foundation, China (2020GXNSFDA238022).
References
-
Mo YM, Chen WC, Wu HY, Zhang W, Zhou SC (2015) A New Species of Odorrana Inhabiting Complete Darkness in a Karst Cave in Guangxi, China. Asian Herpetological Research 1: 11-17.
-
Pham CT, Nguyen TQ, Bernardes M, Nguyen TT, Ziegler T (2016) First records of Bufo gargarizans Cantor, 1842 and O. lipuensis Mo, Chen, Wu, Zhang et Zhou, 2015 (Anura: Bufonidae, ranidae) from Vietnam. Russian journal of herpetology 23(2): 103-107.
-
Gosner KL (1960) A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica 16(3): 183-190.
-
Wu L, Xu RH, Dong Q, Li DJ, Liu JS (1983) A new species of Rana and records of amphibians from Guizhou Province. ACTA Zoological Sinica 29(01): 66-70.
-
Xu J, Liu JX (2009) Odorrana wuchuanensis. Forest & Humankind 2009(01): 46-53.
-
Luo T, Wang SW, Xiao N, Wang YL, Zhou J (2021) A New Species of Odorous Frog Genus Odorrana (Anura, Ranidae) from Southern Guizhou Province, China. Asian Herpetological Research 12(4): 381-398.
-
Liu JX, Zhang ZY, Zhang ZP, Liu XY, Zhang L, et al. (2009) Preliminary report on the habitat and current situation of the critically endangered cavity-dwelling frog, Odorrana wuchuanensis. Bulletin of Biology 44(05): 14-16.
-
Zhao YZ, Guo Y, Zhao HB (2013) Study of environmental factors on geographical distribution of Odorrana in Guizhou. Agricultural technology services 30(7): 763-
-
(2022) List of key protected wild animals in Guangxi. Forestry Bureau of Guangxi Zhuang Autonomous Region.
-
Rogowitz LG, Candelaria CL, Denizard LE, Meléndez LJ (2001) Seasonal reproduction of neotropical, the cave coquí (Eleutherodactylus cooki). Copeia 2001: 542-547.
-
Diesel R, Bäurle G, Vogel P (1995) Cave breeding and froglet transport: a novel pattern of anuran brood care in the Jamaican frog, Eleutherodactylus cundalli. Copeia 1995: 354-360.
-
Lima AMX, Araújo CO, Verdade VK (2012) Cycloramphus eleutherodactylus (Alto button frog): Calling among rocks and caves. Herpetological Bulletin 120: 39-42.
-
Rosa GM, Penado A (2013) Rana iberica (Boulenger, 1879) goes underground: subterranean habitat usage and new insights on natural history. Subterranean Biology 11: 15-29.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework