Phylogenetic Analysis of Two Species of Procamallanus (Spirocamallanus) (Nematoda: Camallanidae) Parasitizing Indian Freshwater Fishes
Two species of previously known nematodes from the family Camallanidae were found in different fish hosts in India: the Gangetic mystus Mystus cavasius, the Butter catfish Ompok bimaculatus, and the Zig-zag eel Mastacembelus armatus. The material collected during the study was from two different locations of district Poonch, Jammu and Kashmir, and district Muzaffarnagar district of Uttar Pradesh, India. For molecular analyses, the genomic DNA was extracted, and the 18S ribosomal RNA gene was amplified and sequenced. Specimens of nematodes from the genus Procamallanus clearly resembled all previously known species based on morphology and were identified as Procamallanus (Spirocamallanus) mysti Karve, 1952, and Procamallanus (Spirocamallanus) bilaspurensis Gupta et Duggal, 1973. Molecular data of both species are provided for the first time in India, and a phylogenetic analysis based on 18S rRNA is presented. Molecular data depicted that P. (S.) mysti and P. (S.) bilaspurensis clustered with each other and formed a separate lineage in the phylogenetic tree with 97% ML and 1.00 BI posterior probability values showing homogeneity in sequences, respectively. This study expands the genetic information for Procamallanus species in addition to morphology and, contributing to the limited knowledge of Indian camallanid species.
Introduction
Parasitic nematodes belong to the family Camallanidae, mostly infect the digestive tract of marine and freshwater fish (and sometimes amphibians and aquatic reptiles), and are distributed worldwide [1, 2, 3, 4, 5, 6, 7, 8]. Camallanid nematodes can be easily distinguished on the basis of morphological features such as the presence of a well-developed buccal capsule that is further supported by different structures like a basal ring, spiral or longitudinal ridges, tridents, etc. [5]. Until now, the camallanid fauna of Indian fish has been poorly studied, and due to a lack of proper studies, numerous species were erected, most of which lacked proper morphological characteristics and molecular data. Recently, in a study by Moravec, et al. [9], it was pointed out that in spite of the large number of taxonomic and faunistic papers for freshwater fish nematodes in South Asian countries, a gap regarding the knowledge of real fauna still persists. According to Moravec, et al. [9], this situation arises because most of the species from this region were not appropriately identified or were poorly described, as well as because many nematode species identifications or their taxonomic problems in the respective groups were just left unsolved.
During parasitological surveys for fish nematodes in the district Muzaffarnagar of Uttar Pradesh and in the Poonch River of the district Poonch of Jammu and Kashmir, India, in 2021, two species of camallanid nematodes were found: Procamallanus (Spirocamallanus) mysti Karve, 1952, and Procamallanus (Spirocamallanus) bilaspurensis Gupta, et al. [10]. In the present study, we provide molecular characterization based on the 18S gene of the found species.
Materials and Methods
Parasite Collection and Morphology
Fish were brought from local vendors of the fish market in the district of Muzaffarnagar, Uttar Pradesh, and caught in the Poonch River, Jammu and Kashmir, close to the border near the LOC (33°30’N, 75°00’E), India, in 2021. Collected nematodes were washed in saline water and then fixed in hot 70% ethanol. For light microscopy examination, they were cleared with glycerin. The voucher specimens were deposited in the Helminthological Collection of the Institute of Parasitology, Biology Centre of the Czech Academy of Sciences, České Budějovice, Czech Republic (IPCAS) (Coll. No. N-196 for P. (S.) mysti and N-1260 for P. (S.) bilaspurensis).
DNA Isolation and Amplification
For molecular analysis, the nematodes were preserved in 95% molecular-grade ethanol after microscopic identification. DNA was extracted using the DNeasy™ Blood and Tissue Kit (Qiagen, Germany), following the standard protocol method recommended by the manufacturer. The polymerase chain reaction for the 18S gene was performed using the primer pairs Nem 18SF + Nem18SR [11]. The thermocycling profile was as follows: 2 min denaturation at 94 °C for 35 s, 35 cycles of 94 °C for 30 s, 54°C for 35 s, and 72 °C for 1 min, and 72 °C for 10 min for a final extension. PCR products were sent to a commercial sequencing company (Eurofins Genomics India Pvt. Ltd., Bangalore, India). DNA products were sequenced in both directions using the above mentioned PCR primer pairs.
Resulting sequences were assembled and trimmed using MEGA v. 11 [12] software and submitted to Gen Bank under the following accession numbers: ON158670, ON158669, OR195335, and OR195336.
Phylogenetic Analysis
Newly generated gene sequences of 18S obtained during this study were aligned with the other Camallanidae family sequences retrieved from GenBank using MUSCLE implemented in MEGA v.11 [12]. Both maximum likelihood (ML) and Bayesian inference (BI) methods were used for phylogenetic analysis as implemented in MEGA v. 11 and TOPALi v. 2.5 [13], respectively. The best-fitting model for the 18S dataset was estimated prior to analyses using jModelTest (V. 2.1.2) [14, 15], and this was GTR+G for the 18S dataset. A Maximum Likelihood (ML) tree was generated with bootstrap support values of 1,000 repetitions. Bayesian Inference (BI) trees were generated in TOPALi v. 2.5 by running two independent MCMC runs of four chains for 107 generations, sampling tree topologies every 1,000 generations, and setting the burn-in period at 25%. Genetic divergences of the 18S gene were calculated using the uncorrected p-distance model in MEGA v. 11, including transitions and transversions. Cosmocercoides pulcher was used as an out group.
Results
Procamallanus species collected during the present study revealed the presence of Procamallanus (Spirocamallanus) mysti Karve, 1952 and Procamallanus (Spirocamallanus) bilaspurensis Gupta, et al. [10] from Mystus cavasius, Ompok bimaculatus, and Mastacembelus armatus, respectively.
Procamallanus (Spirocamallanus) bilaspurensis comprises the following important characteristics: nematodes thick and small, with a transversely striated cuticle, oval mouth aperture present, covered by six flat crescentic elevations with two circles of eight submedian cephalic papillae, each formed by four papillae, including the external circle, which is distinctively larger than others. Buccal capsule looks yellowish, long, with s well-developed basal ring, lateral amphids small and present in a pair, capsule basal part thick, with inner surface of whole capsule shows many spiral thickenings, oesophagus muscular. Deirids small located in the anterior portion at the level of nerve ring. Excretory pore situated between nerve ring and posterior end of muscular oesophagus, both sexes have conical tails.
Procamallanus (Spirocamallanus) mysti encompasses worms having a thick transverse striated cuticle, a pinkish body colour, mouth large and opening of the mouth covered by a membrane ring, submedian cephalic papillae four in number, and two lateral amphids present. Buccal capsule barrel-shaped comprises two sets of spiral thickenings: with male have 17-19 fragmented ridges while females have 19- 21 fragmented ridges; a basal ring present at distal end of buccal capsule, oesophagus anterior end muscular, while the posterior end glandular. Nerve ring present at muscular oesophagus, deirids also present below the nerve ring with a small spike-like protuberance. Excretory pore, situated at the junction of the muscular and glandular oesophagus.
Sequences obtained in the present study for the 18S gene of Procamallanus species were aligned with other sequences downloaded from the GenBank database. The 18S rDNA phylogenetic reconstruction from ML and Bayesian inference (BI) analyses represented a well-supported Procamallanus lineage supported by high bootstrap and posterior probability values (97/1) for both the presently collected specimens (Figure 1). The phylogenetic tree inferred shows that both, Procamallanus (Spirocamallanus) mysti and Procamallanus (Spirocamallanus) bilaspurensis are sister species (Figure 1). They also share a main clade with P. laeviconchus from Sudan and P. pseudolaeviconchus from South Africa, and the relationships among these four species of Procamallanus received strong bootstrap support (97%) and Bayesian posterior probabilities (0.99) (Figure 1). P. (S.) mysti and P. (S.) bilaspurensis exhibit genetic divergence that averages 1.37%. Additionally, the genetic divergence between our isolates with Procamallanus laeviconchus (JF803934) and Procamallanus pseudolaeviconchus (OP602992- OP602994), ranged from 1.97 to 2.23%. The topology of the tree with another Procamallanus species from India, i.e., Procamallanus spiculogubernaculus (KU292357), represents independently in the tree and shows a genetic divergence 2.92% with our isolates (Figure 1).
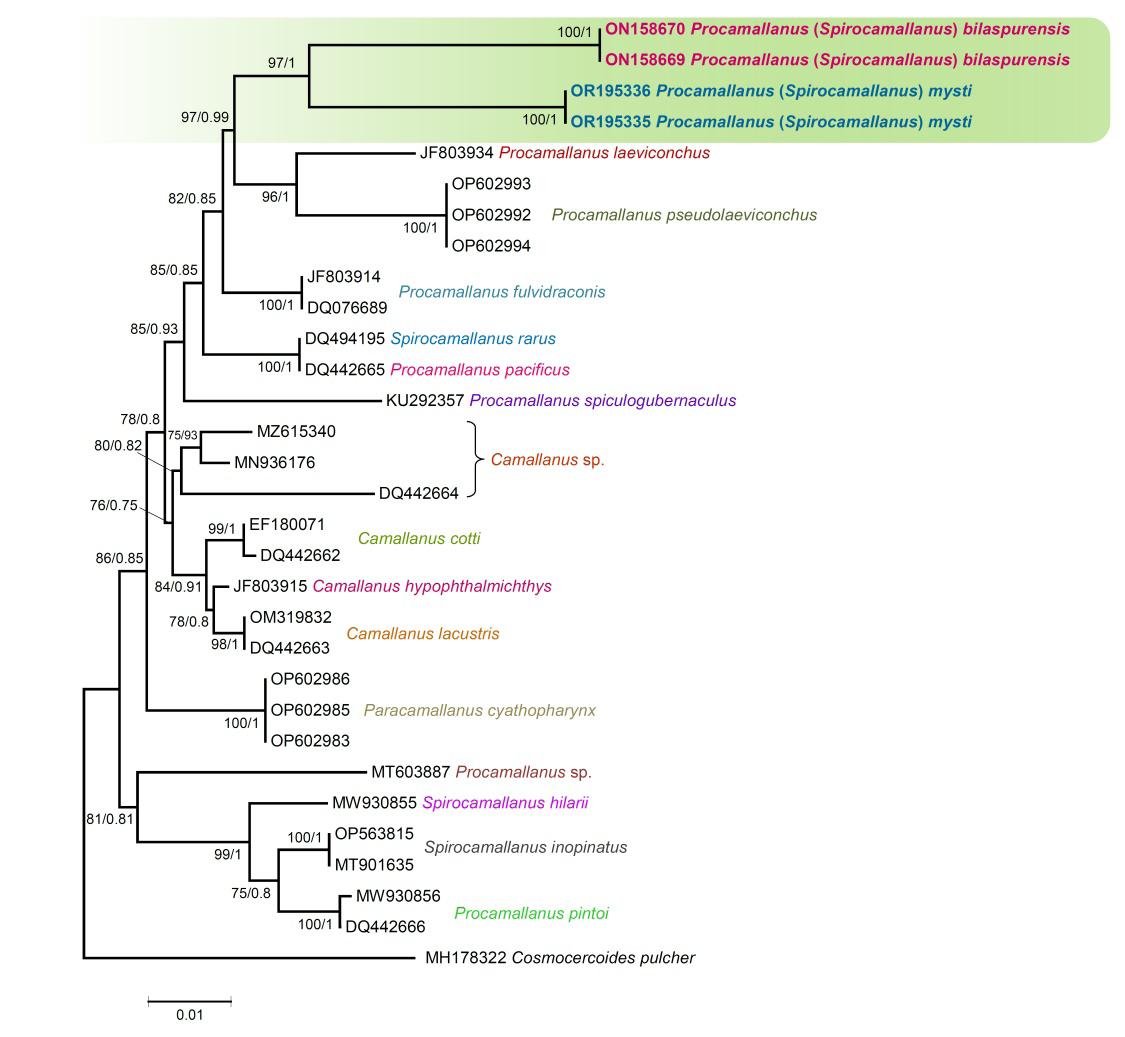
Figure 1: Consensus maximum likelihood and Bayesian inference phylogenetic tree inferred with partial 18S rDNA sequences for species of Procamallanus. Numbers near internal nodes show ML bootstrap and probabilities (BI) values. GenBank accession numbers precede species names. Scale-bar indicates the expected number of substitution per site. Species sequenced in the present study are shown in bold.
Discussion
Of the family Camallanidae Railliet et al. [16] the genus Procamallanus Baylis, 1923, comprises a diverse group of parasites. However, Moravec, et al. [6, 17, 18, 19, 20] cogitated five taxa as subgenera of Procamallanus of the Procamallaninae: Procamallanus Baylis, 1923, Spirocamallanus Olsen (1952), Platicamallanus, Punctocamallanus Moravec et Scholz, 1991; and Denticamallanus Moravec et al. [20]. Furthermore, Batrachocamallanus is also considered a junior synonym of Procamallanus by Moravec, et al. [20], as supported by Rigby, et al. [5]. Procamallanus Baylis, 1923 encompasses a large number of species infected freshwater and marine fishes [21]. From freshwater fishes of southern and south-eastern Asia, especially from India, many nominal species were described [22]. Although, many species of Procamallanus were frequently described from India, often with inadequate morphology, and without molecular data on them, so a thorough comparison of them is impossible [22, 23, 24]. This condition is more complicated when these nematodes are described from similar hosts, creates taxonomic confusions and rarely the type or voucher specimens for most species are available except few by Sood [22].
Although camallanid nematodes have different and sufficient morphological characters to identify them, the differentiation between species is complicated [5]. Though, the use of these characteristics between species is rather confused due to the presence of sexual dimorphism, taxonomy based on morphological structures like the buccal capsule is not stable and therefore requires revision [25, 26]. Consequently, these authors contemplated the subgenera Procamallanus “for practical reasons” [25], to which Rigby and Rigby [5] also diagnosed the Procamallanus subgenera “for the sake of tradition and simplicity”. Besides the buccal capsule, the other trustworthy character for identification was considered the male caudal region. Initially, studies described that camallanid nematodes possess only a right spicule, whereas later descriptions also mentioned that the left spicule is also present in species that were previously described with a right one [7, 20, 27]. The tail tip of adult females of this group of parasites comprises mucrons, and their presence and absence are also treated as reliable characters for identification [28].
According to Sood [22], from the host Mastacembelus armatus, the following species of Procamallanus are listed from India: P. bilaspurensis Gupta, et al. [10], P. fulvidraconis Li, 1935, P. gubernaculus Khera, 1955, and P. meszarosi Arya, 1984. Of these, P. fulvidraconis, P. gubernaculus, and P. kalriai species are in the category of species inquirenda by Sood [22], who predicted that these species were originally described based on misidentifications. The present nematodes collected from M. armatus belong to P. bilaspurensis, which comprises spiral ridges in the capsule, although these ridges are intervallic and not developed properly. The present specimens of P. bilaspurensis are placed in the subgenus Spirocamallanus Olsen, 1952, because of the presence of spiral ridges in the buccal capsule of the males and females, and mentioned as Procamallanus (Spirocamallanus) bilaspurensis [9, 25]. In case of Procamallanus (Spirocamallanus) mysti Karve, 1952, comprises two unequal spicules, the left one is weakly sclerotized while the right spicule is well sclerotized, and distinctly visible. P.(S.) mysti differentiated from other congeners by having the number of spiral thickenings of buccal capsule and the number of caudal papillae in male while the number of mucrons in the female tail tips [21, 29].
Numerous molecular studies were published recently based on 18S rDNA gene data that included camallanid nematodes, but the phylogenetic relationships between the species and the status of different taxa within the family are still in question and not resolved [7, 30, 31, 32]. To date, in India, only a few molecular studies have been conducted that included very few species, therefore, in the present study, we follow the genetic identification of the species, collected herein and their molecular characterization based on the 18S gene. Our phylogenetic analysis supported the presence of two different Procamallanus species: Procamallanus (Spirocamallanus) mysti and Procamallanus (Spirocamallanus) bilaspurensis, respectively. Phylogenetic analysis demonstrates that the two species studied here (P. mysti and P. bilaspurensis) are closely related, while another Indian freshwater species, P. spiculogubernaculus, occupies separate positions in the 18S based tree. However, a lack of molecular data from Indian camallanid species can influence the present phylogenetic position of the species in future studies. Thus, according to our opinion, all the species, whether they are previously well-known or new ones, should be supported by proper descriptions/illustrations, and molecular data.
Conclusion
Two species of the genus Procamallanus are clearly recognized in the Indian region using molecular data- Procamallanus (Spirocamallanus) mysti and Procamallanus (Spirocamallanus) bilaspurensis, and the phylogenetic analysis demonstrates that both species are closely related. Using an amalgamation of morphology and molecular data together is the only way to correct species identification with regard to Camallanid nematodes.
Conflicts of Interest
The authors declare no competing interests.
Acknowledgements
We are grateful to the Head, Department of Zoology, D.A.V. College Muzaffarnagar, India and Chaudhary Charan Singh University, Meerut, Uttar Pradesh India, for providing laboratory facilities.
Funding
This research was funded by Research and Development Scheme of the Higher Education Department, Uttar Pradesh Government, India, grant number 93/2021/1401/ sattar-4-2021-4(28)/2021.
References
-
Santacruz A, Barluenga M, Pérez-Ponce de León G (2021) Taxonomic assessment of the genus Procamallanus (Nematoda) in Middle American cichlids (Osteichthyes) with molecular data, and the description of a new species from Nicaragua and Costa Rica. Parasitol Res 120: 1965- 1977.
-
Moravec F, Justine JL, Rigby M (2006) Some camallanid nematodes from marine perciform fishes off New Caledonia. Folia Parasitol 53(3): 223-239.
-
Mašová Š, Baruš V, Moravec F (2011) New morphological data on the first-stage larvae of two Procamallanus species (Nematoda: Camallanidae) based on SEM studies. Folia Parasitol 58(4): 318-321.
-
Anderson RC, Chabaud AG, Willmott S (2009) Keys to the Nematode Parasites of Vertebrates: Archival Volume. CABI, Wallingford pp: 463.
-
Rigby M, Rigby E (2014) Order Camallanida: Superfamilies Anguillicoloidea and Camallanoidea. In: Schmidt-Rhaesa A (Ed.), Nematoda. Walter De Gruyter GmbH; Berlin/Boston, pp: 637-659.
-
Moravec F, Justine JL (2019) New species and new records of camallanid nematodes (Nematoda, Camallanidae) from marine fishes and sea snakes in New Caledonia. Parasite 26: 66.
-
Svitin R, Truter M, Kudlai O, Smit NJ, du Preez L (2019) Novel information on the morphology, phylogeny and distribution of camallanid nematodes from marine and freshwater hosts in South Africa, including the description of Camallanus sodwanaensis n. sp. IJP: PAW 10: 263-273.
-
Lebanan K, Mohilal N (2021) Camallanus sonaiensis sp.n. (Camallanidae), a new species of nematode from Heteropneustes fossilis collected from Assam, India. J Parasit Dis 45: 664-672.
-
Moravec F, Chaudhary A, Ahmed M, Shanker Singh H (2022) New data on the morphology and taxonomy of some spiruridan nematodes (Spirurida) parasitising fishes in Jammu and Kashmir, India. Folia Parasitol 69: 002.
-
Gupta NK, Duggal CL (1973) On one new and one already known species of the subgenus Procamallanus (Baylis, 1923) Ali 1956 (Nematoda: Camallanidae) from the freshwater fish and a key to the species of the subgenus. Riv Parassitol 34: 295-304.
-
Floyd RM, Rogers AD, Lambshead PJD, Amith CR (2005) Nematode-specific PCR primers for the 18S small subunit rRNA gene. Mol Ecol Notes 5: 611-612.
-
Tamura K, Stecher G, Kumar S (2021) MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol 38: 3022-3027.
-
Milne I, Lindner D, Bayer M, Husmeier D, McGuire G, et al. (2009) TOPALi v2: a rich graphical interface for evolutionary analyses of multiple alignments on HPC clusters and multi-core desktops. Bioinformatics 25(1): 126-127.
-
Guindon S, Gascuel O (2003) A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol 52: 696-704.
-
Darriba D, Taboada GL, Doallo R, Posada D (2012) jModelTest 2: More models, new heuristics and parallel computing. Nat Methods 9: 772.
-
Rigby MC, Rigby E (2013) Order Camallanida: Superfamilies Anguillicoloidea and Camallanoidea. In: Schmidt-Rhaesa A (Ed.), Handbook of zoology, Gastrotricha, Cycloneuralia and Gnathifera. Nematoda. Deutsche Nationalbibliothek, Hamburg, 2: 637-659.
-
Moravec F, Van As L (2015) Procamallanus (Procamallanus) spp. (Nematoda: Camallanidae) in fishes of the Okavango river, Botswana, including the description of P. (P.) pseudolaeviconchus n. sp. parasitic in Clarias spp. (Clariidae) from Botswana and Egypt. Syst Parasitol 90: 137-149.
-
Moravec F, Van As L (2015) Procamallanus (Spirocamallanus) spp. (Nematoda: Camallanidae) from fishes of the Okavango river, Botswana, including P. (S.) serranochromis n. sp. parasitic in serranochromis spp. (Cichlidae) Syst Parasitol 90(2): 151-164.
-
Moravec F, Jirků M (2015) Two Procamallanus (Spirocamallanus) species (Nematoda: Camallanidae) from freshwater fishes in the lower Congo river. Acta Parasitol 60: 226-233.
-
Moravec F, Justine JL, Rigby MC (2006) Some camallanid nematodes from marine perciform fishes off New Caledonia. Folia Parasitol 53: 223-239.
-
De NC, Moravec F (1980) Redescription of the nematode Procamallanus spiculogubernaculus with notes on related forms. Acta Soc Zool Bohemoslov 44: 81-91.
-
Sood ML (2017) Fish Nematodes from South Asia. In: 2nd (Edn.), Kalyani Publishers, Ludhiana, pp. 1039.
-
Sanachaoba RK, Gambhir RK (2018) A new species of Camallanidae (Raillet and Henry, 1917) from a siluroid catfish in Barak River, Jiribam, Manipur (India). J Parasit Dis 42(2): 182-186.
-
Devi NP, Bidyalakshmi TH, Gambhir RK (2020) A new species of Procamallanus (Baylis, 1923) from Mastacembalus armatus from Iril River, Imphal, East, Manipur, India. UP J Zool 41(9): 65-71.
-
Moravec F, Thatcher V (1997) Procamallanus (Denticamallanus subgen. n.) dentatus n. sp. (Nematoda: Camallanidae) from the characid fish, Bryconops alburnoides, in the Brazilian Amazon. Parasite 4: 239-243.
-
Moravec F, Scholz T (1991) Observations on some nematodes parasitic in freshwater fishes in Laos. Folia Parasitol 38(2): 163-178.
-
Moravec F, Gey D, Justine JL (2016) Nematode parasites of four species of Carangoides (Osteichthyes: Carangidae) in new Caledonian waters, with a description of Philometra dispar n. sp. (Philometridae) Moravec & Thatcher 1997. Parasite 23: 40.
-
Moravec F (1975) The development of Procamallanus laeviconchus (Wedl, 1862) (Nematoda: Camallanidae). Acta Soc Zool Bohemoslov 39: 23-38.
-
De NC, Roy R, Majumdar G (1986) Redescription of Spirocamallanus mysti (Karve, 1952) (Nematoda: Camallanidae) with notes on related forms. Folia Parasitol 33: 353-361.
-
Černotíková E, Horák A, Moravec F (2011) Phylogenetic relationships of some spirurine nematodes (Nematoda: Chromadorea: Rhabditida: Spirurina) parasitic in fishes inferred from SSU rRNA gene sequences. Folia Parasitol 58: 135-148.
-
Sardella CJ, Pereira FB, Luque JL (2017) Redescription and first genetic characterisation of Procamallanus (Spirocamallanus) macaensis Vicente & Santos, 1972 (Nematoda: Camallanidae), including reevaluation of the species of Procamallanus (Spirocamallanus) from marine fishes off Brazil. Syst Parasitol 94: 657-668.
-
Chaudhary A, Verma C, Tomar V, Singh H (2017) Procamallanus spiculogubernaculus Agarwal, 1958 (Nematoda: Camallanidae) from Stinging catfish, Heteropneustes fossilis in India: morphological characterization and molecular data. Helminthologia 54(1): 68-76.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind