A New Amphisbaena species from a Restinga Habitat of Southeastern Brazil (Reptilia, Squamata, Amphisbaenidae)
A new species of Amphisbaena is described from the restinga environment of Grumari in Rio de Janeiro municipality, Rio de Janeiro State, Brazil. This small form has a total length of 115.0 mm, a maximum 3.7 mm diameter at midbody, 186 left ventral annuli, 21 caudal annuli, 26 – 28 annuli scales at midbody and four precloacal pores. Autotomic tail annulus is absent. Characteristically, it has an enlarged pair of temporal shields coupled with a distinctive lateral tail compression. The new species is easily recognizable and distinguishable from any of the other Amphisbaena forms. There are only two Amphisbaena species effectively registered for Rio de Janeiro State (A. alba and A. hogei), and the species here described would add the third known Amphisbaena species for Rio de Janeiro state and raise the number of Amphisbaena species registered for Brazil to 71.
Introduction
The existing biodiversity on the planet is immense, especially in tropical areas, but we are still far from knowing how many species actually live on Earth. Every day more and more species are discovered at an increasing rate, and projections show that we are still far from knowing the real biodiversity for different groups of animals and plants [1, 2], among other living forms. For example, just for reptiles, which are currently known to exist 11,940 species, 120 of these species were described just between July and December 2022 [3]. In a negative opposing force, human action on Earth increasingly promotes an erosion of existing biodiversity on land and on landscapes, on all continents and oceans and, every year, many species are extinct, many of them before science register its existence [4].
The Atlantic Rainforest of southeastern Brazil and their associated habitats is a Biome known for its high biological diversity [5, 6]. Each year an increasing number of new species of different groups are formally described [7]. The more you move within the forest the more you experience a considerable chance to stumble upon an undescribed species.
Species richness in the Atlantic Rainforest biome is also expected to be high for cryptic animals such as the Amphisbaenia, frequently abundant but hard to find due to their fossorial habits [8]. The Sub-Order Amphisbaenia includes more than 200 living species, most of them occurring in Brazil [9, 10, 11, 12, 13, 14, 15, 16]. In Brazil, The genus Amphisbaena (Family Amphisbaenidae) has the greatest number of species in the Sub-Order, 70 in total [17], with the number of known species in this genus increasing sharply in the last two decades [9, 11, 12, 16].
In the state of Rio de Janeiro, which is totally included within the Atlantic Forest biome, the Suborder Amphisbaenia is currently represented by six species: Leposternon infraorbitale, L. microcephalum Wagler, 1824, L. scutigerum and L. wuchereri , Amphisbaena alba Linnaeus, 1758 and A. hogei Vanzolini, 1950 [17, 18, 19, 20, 21] (the occurrence of A. prunicolor (Cope, 1885) for Rio de Janeiro state is not confirmed [17].
On 20 September 1997 while giving an Ecology field class in a restinga habitat (one of the associated habitats of the Atlantic Forest) one of us (CFDR) spread the leaf litter on the ground while explaining to the students about the rich fauna that live in such microhabitat, when suddenly an amphisbaenid emerged from the leaf litter and promptly started to move among the leaves. The individual was captured by hand to show and to explain the students about that interesting form of life. Later, after careful examination, it was realized that the individual differed markedly from any other amphisbaenid previously described. Here we describe a new species of Amphisbaena for the Atlantic Forest Biome in the state of Rio de Janeiro.
Material and Methods
The specimen was an unusual small-sized Amphisbaena with four anal pores and a distinctive compressed tail, undoubtedly an intriguing form.
The Grumari restinga (23°05’S; 43°30’W) is located in Rio de Janeiro Municipality, Rio de Janeiro State, at the southeastern Brazilian coast (Figure 1). Restinga habitats are part of the Atlantic Forest Biome, occurring in lowlands along most of the Brazilian coast, and are characterized by sandy soils and xerophyllous vegetation [22]. Restinga habitats are part of the Atlantic Forest Biome, occurring in lowlands along most of the Brazilian coast [22]. The climate in the area is markedly seasonal with a rainy season from November to March and a dry season from May to September [23, 24]. Mean annual temperature in the area averages 23ºC and mean annual rainfall ranges from 1000 to 1350mm [24].
The vegetation in Grumari is dominated by plants of the Families Leguminoseae, Bromeliaceae, Cactaceae and Arecaceae [25], and four main vegetation types can be found: i) the beach area, dominated by herbaceous vegetation; ii) a coastal sand dune dominated by Cactaceae (Cereus and Pilosocereus) and Bromeliaceae; iii) an open restinga, dominated by shrubby vegetation up to 3 m tall; and iv) forested restinga dominated by shrubby-arboreous vegetation up to about 7 m tall [25].
In the Laboratory the specimen was anesthetized with ether, fixed in 4% formalin and preserved in 70% alcohol. The pholidosis nomenclature follows Gans C, et al. [26] and Vanzolini PE, et al. [27] with adaptations.
The measurements were taken with a dial caliper approaching the nearest 0.01 mm, and meristic data were recorded with a ruler, to the nearest millimeter. Sex determination was not possible.
The MNRJ collection acronym follows [28]. The collecting time is the natural one (10:15h), not the Brazilian Summer Time Adjustment - Horário Brasileiro de Verão (9:15h). Species description Amphisbaena carioca sp. nov (Figure 2).

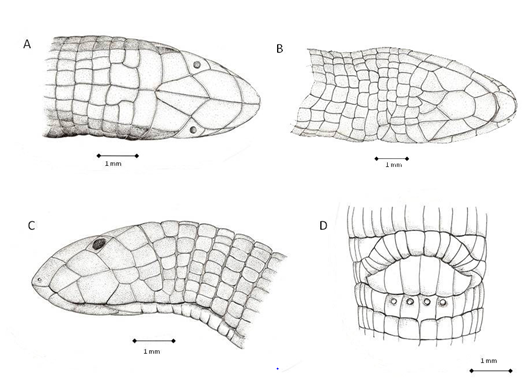
Figure 2: Pictures of the dorsal (A), frontal (B), ventral (C), lateral (D) views of Amphisbaena carioca head and view of the tail (E). F – frontal; G – genial; I – infralabial; MG – medial genials; N – nasal; O – occipital; Oc – ocular; P – parietal; PF – prefrontal; PG – postgenials; PR – postgenial row; R – rostral; SL – supralabial; T – temporal; 1st B – first body annulus. Bars represent 1 mm.
Holotype: MNRJ 18293, Restinga de Grumari (23°05’S; 43°30’W), Rio de Janeiro Municipality, state of Rio de Janeiro, Brazil, collected by C. F. D. Rocha & M. Van Sluys on 20 September 1997.
Diagnosis: A slender small-sized Amphisbaena with 115.0 mm total length and a maximum of 3.7 mm diameter at midbody, without major fusion of the head shields; 186 left ventral annuli, 21 caudal annuli, 26 -28 annuli scales at midbody and four precloacal pores; only the lateral sulci well expressed, autotomic tail annulus absent; and characteristically an enlarged pair of temporal shields coupled with a distinctive tail lateral compression. Dorsal coloration is light brown and ventral coloration is white- yellowish.
Description of the Holotype
Description: A slender, small-sized Amphisbaena, with 115 mm total length and maximum width of 3.7 mm at midbody, and without major head shield fusions (Figures 1 & 2). Measurements and meristic data are as follow: Ventral Annuli, Left side – 186; Ventral Annuli, Right side – 184; Dorsal Annuli, Left side – 183; Dorsal Annuli, Right side – 184; Lateral Annuli, Right/Left sides – 2, 2; Caudal Annuli, counted in the ventral left side from the cloaca to the tip of the tail – 21; Ventral Annuli Scales, counted for five adjacent annuli at midbody - 16; Dorsal Annuli Scales, counted for five adjacent annuli at midbody – 10 – 12; Precloacal Flap Shields – 6; Postcloacal Flap Shields – 13; Maximum Head Width, measured at temporal region – 3.5 mm; Body Width, measured at half of body length – 3.7 mm; Maximum Caudal Width, measured at its base – 3.0 mm; Minimum Caudal Width, measured after its base – 1.5 mm; Snout-Vent Length – 100 mm; Caudal Length – 15 mm; Total Length – 115 mm.
Shields of Dorsal Half of Head
Rostral: an almost triangular shield, scarcely visible from above. There are large posterior sutures with the pair of nasals and small vertical lateral-inferior sutures with the first supralabials.
Nasals: a pair of quadrangular shields, smaller than the (posterior) prefrontals. The rounded, small nostrils open laterally, and are situated in the anterior-lateral part of the nasals, near the rostral. Each nasal is sutured inferiorly with the first supralabial and is in point contact with the second supralabial at the posterior and inferior corner. Nasals, prefrontals and frontals are in broad contact at midline.
Prefrontals: the largest pair of shields atop head, each shaped as an irregular pentagon. They are successively laterally sutured with the second supralabial and with the ocular, and posteriorly with the frontals. The posterior sutures together shape a large “U” with the anterior portion of the frontals.
The nasals and prefrontals have a convex dorsal curvature and are in a conspicuous level above the first and second supralabials and oculars, on a discrete arrangement.
Frontals: a pair of shields shaped individually almost as an elongated scalene triangle, and together resembling a lozenge. However, the corresponding anterior “V” portion of the lozenge, sutured with the prefrontals, is more like a “U” and is smaller than the posterior “V”, which is inserted between the superior temporal shields. The frontals touch the oculars at the angles formed laterally by the point contact of the “U” and the “V” portions. The posterior extremity of the “V” angle contacts the medial parietal shields.
Oculars: a pair of trapezoidal shields, the posterior side the largest, and the anterior portion forming a “V”. The oculars are nearly flattened and lay only on the dorsal, not lateral head level. The rounded dark eyes are clearly visible and anteriorly positioned between the “V” sides. The eyes are oriented in a more frontal than lateral condition.
Superiorly, the oculars touch the frontals, separating in that point the posterior angle of the prefrontal from the anterior angle of the superior parietal. Anterior and superiorly they suture with the prefrontal and anterior and inferiorly with the second supralabial. Posteriorly, the suture is with the superior shield of the temporal row. Inferiorly the oculars suture with the third supralabials at a right angle.
Supralabials: There are three supralabials, the first is trapezoidal and the longest, the second is larger and irregularly pentagonal, the third is the tallest and also irregularly pentagonal. Sutures between supralabials run anteriorly at angles of approximately 45o and 75 o. The first supralabial sutures anteriorly with the rostral and superiorly with the nasal; the second supralabial contacts anteriorly the nasal, sutures superiorly with the prefrontal, and sutures posterior and superiorly with the ocular; and the third supralabial sutures anterior and superiorly with the ocular, touches superiorly the superior temporal, and sutures posteriorly with the two remaining temporals.
Temporal Rows: There are three temporal shields, arranged in a vertical row and decreasing in size from above to below. The superior and medial shields are irregular trapezoids, the inferior is horizontally rectangular. The superior temporals have an area equivalent to the nasals and are separated medially by the frontals; anteriorly they are sutured to the oculars and posteriorly with the medial parietals. The medial temporal sutures anteriorly with the third supralabial and posteriorly with the lateral parietal. The inferior temporal sutures anteriorly with the third supralabial shield, posteriorly with a shield of the row corresponding to the second ventral body annulus, and inferiorly with a shield of the row corresponding to the first ventral body annulus.
The superior temporals are flattened, without the discrete expression of the temporal bulges that are very usual in amphisbaenids at this place. As the oculars, the superior temporals lay totally on the dorsal head level, in almost right angle with the medial temporals.
Parietals: There are two pairs of parietals, medial and lateral. The medial parietals are a pair of transversely, almost rectangular shields (but with more angulations), three times larger than longer. They are very conspicuous atop head, in a diagnostic way. They suture anteriorly with the frontals (at midline) and the superior temporals. At right side, the anterior angle is just in point contact with the medial temporal; at left side, there is a very small suture with the medial temporal. The medial parietals suture at sides with the lateral parietals, and posteriorly with the shields of the first occipital row. The median suture between them is shortened anteriorly by the posterior insertion of the frontals, and posteriorly by the insertion of the medial occipitals. The medial parietals lay totally on the dorsal head level.
The lateral parietals are rectangular but smaller shields, and lay on the lateral head level, with a distinct angulation relative to the medial parietals. They are anteriorly sutured with the medial temporal and posteriorly with the first scale of the occipital row. On the right side, the inferior angle is separated from any contact with the inferior temporal; on the left side, there is a very small suture with the inferior temporal. The inferior suture of the lateral parietals rests on a somewhat enlarged shield, almost rectangular on the right side and somewhat squarish on the left side. This shield is common to the dorsal extra half-annulus of the parietals and to the complete annulus where the first occipitals lay (first occipital row).
Occipitals: There are two rows of occipitals, corresponding dorsally to the ventral part of the second and third complete body annuli (besides the fact that the second “complete” annulus is strangled in a point at its left inferior side by two pair of scales of the first and third body annuli). The medial pair of shields of the first row is posteriorly almost fused with the larger medial pair of the second row, the thin sutures only perceived under magnification.
Dorsal and laterally, the occipital rows also form an acute angle with the lateral head plan, but in a lesser degree than the parietals. In frontal view, the head is characteristically “suqarish” .
Shields of Ventral Half of the Head
Genial: A large shield, with more or less the same area of the postgenial, matching anteriorly with the position of the rostral. Its anterior border is convex, the posterior suture a transversal line with the postgenial. The lateral sutures with the first infralabials have a general vertical orientation but are not straight lines.
Postgenial: A large heptagonal shield, sutured anteriorly with the genial. Anterior-laterally it is sutured with the first infralabials, laterally in longer longitudinal sutures with the second infralabials, and posteriorly in an inserted “V” suture with the medial-genials.
Medial-Genials: There are four medial-genial shields, confined between the postgenial anteriorly, the second infralabials and the lateral-genials laterally, and the postgenial row posteriorly. The general arrangement would be of three shields, two laterals and one medial, but the left lateral one is cut in two. The right lateral shield and the anterior left lateral one are irregular quadrilaterals, sutured anteriorly with the postgenial and laterally touching the first infralabial, then with a longer longitudinal suture with the lateral genials. The medial shield is a smaller irregular quadrilateral, touching median and anteriorly the postgenial “V” extremity, and sutured laterally with the more lateral medial-genial shields. The left and posterior lateral shield is rectangular, sutured laterally with the lateral-genial. The posterior sutures of the medial-genials (the right lateral, the medial and the posterior left ones) are somewhat rounded or pointed, inserted between the shields of the postgenial row.
Lateral-genials: The lateral-genials are a pair of large shields shaped as irregular pentagons. The anterior suture with the second infralabial is a sloping line with the lateral extremity placed posteriorly. The medial suture with the medial-genials is a longitudinal line, as the short line suturing laterally with the third infralabial. A post-medial and very short suture appears with a medial shield of the postgenial row, and there is a transversal posterior suture with the more lateral and larger shield of the postgenial row.
Postgenial row – There are six shields in the postgenial row. The most laterals are large and in fact shaped as irregular pentagons, but seem almost horizontal rectangles: however, the posterior suture with the scales of the first ventral body annulus is not a straight line, having an angulation. Laterally they suture with the third infralabials, and the transversal anterior suture aligns totally with the lateral-genials. The four small and medial postgenial shields are roughly as longitudinal rectangles, but their corners and anterior borders are rounded. These four shields suture anteriorly with the medial-genials shields and are sutured posteriorly with the scales of the first ventral body annulus.
Infralabials: The three pairs of infralabials are quite different in area and shape. The first pair has a trapezoidal shape, is the second in total area (a bit less than the area of the genial), and sutures post-medially with the postgenial shield. The second infralabials are irregular pentagons and the greater ventral shields of the head. They suture medially with the postgenial shield and shortly with the medial-genials. The post-medial suture with the lateral-genial shield is a long sloping line with the anterior extremity medially positioned. The third infralabial shield pair is roughly rectangular, longitudinally oriented, and the smallest in area. They suture medially with the lateral-genial shields and with the more lateral shields of the postgenial row (a short and a longer line respectively) and posteriorly with a scale of the first ventral body annulus.
Body Sulci
The dorsal and ventral sulci are expressed only by an alignment of the body scales at midline. They are discrete in the tail, running to touch the terminal shield. The lateral sulci are better discernible about the 18th body annulus, ending in the last body annulus carrying pores. They are thinner than the neighbor body shields and are characterized in having two small, reduced triangular shields per body scale, with the more acute angle pointing to each other. These triangular shields increase in size gradually from craniad to caudad, so anteriorly they are totally apart and posteriorly nearly touching each other.
Dorsal and Ventral Body Half-Annuli and Body Scales
The meristic data was given above. The half-annuli pattern is regular, without incomplete half-annuli. The first ventral body (half-) annulus, immediately behind the postgenial row, includes the temporal row laterally and the posterior part of the frontals dorsally. The second ventral (half-) annulus is strangled in a point at its left side by two pair of scales of the first and third body annuli; it continues laterally and dorsally as the first occipital row. At the level of the supralabials, it shares a large shield with an intercalated head half-annulus (for the parietal shields), placed between the first occipital row posteriorly and the temporal row plus frontals anteriorly. The third ventral (half-) annulus continues laterally and dorsally as the second occipital row. The posterior dorsal and ventral body half-annuli are regular and well aligned with each other, with rare exceptions. The third to fifth body annuli are somewhat narrowed in diameter, detaching the head from the rest of the body.
Fusions or unusually shaped scales are rare and insignificant for characterization. The scales of the thirty anterior dorsal half-annuli have a general squarish aspect, but backwards they stretch and are clearly longer than wide. The ventral pair of medial scales is larger than longer but ascending to the lateral sulci the shape changes first to squarish and then to a definitively longer than wide rectangle for the scales delimiting the sulci. The sulci delimiting border of these scales are convex, not straight lines.
Lateral Annuli, Cloacal Region and Tail
There are only two rows of lateral annuli, at the sides (and corresponding dorsal position) of the cloacal flaps. Their scales have the same general shape and disposition of the body ones (longitudinal rectangles). The last ventral body annulus has four well expressed rounded pores at the posterior half of its four medial scales. The precloacal flap follows immediately, with three main pairs of shields and a convex posterior border. The medial pair is the longer and longitudinally rectangular as its neighbor; the more lateral pair is trapezoidal and wider than longer. The postcloacal flap has 13 longitudinal rectangular shields placed in arc, the medial pair larger than the other tinny shields.
The tail does not have any identifiable autotomy annulus, nor seems capable of autotomy. Its base (five anterior annuli) doubled the diameter of the rest, because posteriorly to the base the tail has a strong lateral compression towards its tip. The dorsal and ventral sulci are identifiable. The annuli are complete, encircling all the tail without irregularities, but the scales of successive annuli are not always aligned. The scales are longitudinal rectangles, the ventral ones larger, except for the extreme posterior single semispherical smooth large scale.
Coloration
Dorsal coloration of the preserved specimen is light brown, somewhat faded in the head central portion. The pigment is evenly spread in the scales, but a rounded faded central area can be noted in each one. Laterally the color faints in the head and in the body, and mid-ventral coloration is white-yellowish. The whole tail maintains almost the same brown pigmentation, except for the posterior extremity of the last shield, somewhat lighter than the belly.
Differentiation from Other Amphisbaena Species
Most Amphisbaena species with four pores cannot be confused with A. carioca because of the striking differences in annuli counts, presence of autotomic annulus, total size, head pholidosis and geographic [29]. For Rio de Janeiro State, there are only three other Amphisbaena species, easily distinguished from carioca (in brackets) by the following characters (among others).
A. alba: total size up to 800 mm / largest diameter around 10 mm (115 mm / 3.7 mm); relatively small parietal shields (very large parietals); 212-251 body annuli (186); 4-10 pores (4); short, rounded tail (relatively long and laterally compressed); yellowish or dark brown dorsal pigmentation (light brown).
A. prunicolor - small parietal shields (very large parietals); long rounded tail (smaller and laterally compressed); clearly marked autotomy site on the sixth to ninth tail annuli (without autotomy site); checkerboard ventral pattern (yellowish white).
A. hogei: small parietal shields (very large parietals); rounded tail (compressed after base); clearly marked autotomy constriction on the fourth to seventh tail annuli (without autotomy site).
The other Brazilian Amphisbaena species with a similar tail shape is A. ridleyi from Fernando de Noronha Island, off the coast of northeast Brazil [30]. But A. ridleyi has small parietal shields (very large), 13-17 tail annuli (21), 16-18 dorsal / 20-28 ventral segments to a midbody annulus (10- 12 / 16), and dark brown dorsal head pigmentation (faded light brown).
Etymology: The specific epithet “carioca” is a noun in apposition, in allusion to the nickname given to people natural from Rio de Janeiro City (“carioca”) since the new species is currently known only from Rio de Janeiro Municipality.
Distribution: Amphisbaena carioca is known only from the type locality, by sea level, on the western portion of the coastal Restinga habitats (Restinga of Grumari) of Rio de Janeiro Municipality, within the Atlantic Forest Biome.
Natural History: The specimen of Amphisbaena carioca was collected at 10:15h a.m. presumably in activity on the leaf litter of the forested restinga habitat; it emerged from the litter after it was disturbed by one of us (CFDR). Until other specimens are collected, it is not possible to infer if the specimen is an adult or a juvenile. Comprehensibly, the sex cannot be known without damaging the holotype. Since it is a small specimen, there was no attempt at hemipenial evertion.
Discussion
Descriptions of species based on a single specimen are not recommended taxonomic procedure. However, in the case of amphisbaenians, on account of their fossorial habits and consequent collecting difficulties, there are many exceptions to this rule. In fact, not only for old descriptions [18, 31, 32], but also for the more recent descriptions of A .arenaria [33], A. arda [34], A. brevis [16], and Anops acrobeles [15], the distinctiveness of the forms and eventually the unusual habitats and the geographic location are undisputed reasons for valid descriptions. The same applies for Amphisbaena carioca.
There are only two effectively confirmed Amphisbaena species reported for Rio de Janeiro State [17, 21]: Amphisbaena alba [20], [32] and A. hogei Vanzolini, since the occurrence of A. prunicolor (Cope, 1885) for Rio de Janeiro state is just reported but still not confirmed [17]. So, Amphisbaena carioca would add the third effectively registered species for Rio de Janeiro state. It is important to highlight that all these three species were described based on three or less specimens, a situation similar to A. carioca. Of the four Leposternon species known for Rio de Janeiro State, two (L. infraorbitale and L. wuchereri) are referred from just one specimen [19]. Those specimens were not located in scientific collections and there is no information on their specific localities [19]. So, the species description based on one or a few specimens is not a rare condition for living organisms as secretive as amphisbaenians. Only L. microcephalum and L. scutigerum are well represented in collections, with expressive quantitative samples [19]. We recommend that sampling effort focusing on amphisbaenians for Rio de Janeiro State be increased.
The restinga environment is not typical of amphisbaenians [20], yet they are found in almost all major Brazilian Biomes [14], including those with sandy soils (such as the semi-arid Caatingas, Rodrigues, 2002) [22]. In Rio de Janeiro State, Amphisbaena alba (which occurs in different habitats throughout South America, Gans [35] occurs in restingas [20, 36]. At the restinga habitat of Jurubatiba, A. alba is sympatric with L. scutigerum [36]. Therefore, the occurrence of amphisbaenians in restinga habitats may not be as unusual as previously considered. In our opinion, their rarity in such environments may result from the lack of consistent sampling and difficulty of finding these fossorial organisms. The finding of another Amphisbaena species in the restinga of Grumari supports this idea.
The horizontal alignment of the lateral sutures of the medial occipitals, medial parietals, superior temporals, oculars, and an accentuated angle in the shape of the second and first supralabials mould an acute angle between the dorsal and lateral planes of the head of A. carioca. In frontal view, the head is more “squarish”, in opposition to the ordinary amphisbaenid elliptic conformation. This structure favors the more frontal disposition of the eyes, a very unusual pattern for amphisbaenians. The “squarish” shape, coupled with the convexity and different level disposition of the nasals/ prefrontals conjoint, give a very distinct, maybe unique head structure arrangement for A. carioca.
The compressed tail characteristic of A. carioca is unique among amphisbaenians. The only other species with such a tail shape is A. ridley, endemic to Fernando de Noronha Island off northeastern Brazil [30]. Even though fixation procedures can result, in some cases, in some artificial tail narrowness, for A. carioca this seems not to be the case because its tail appears to be naturally compressed. There is no explanation for such an adaptation in both species yet.
Acknowledgements
We are grateful to D. Vrcibradic for kindly revising the manuscript and offering helpful suggestions and to R. Vamos for taking some of the pictures. P. Nascimento kindly made the drawings of the specimen. CFDR received grants from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (Processes Nos. 302974/2015- 6, 424473/2016-0 and 304375/2020-9), Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) through the Cientistas do Nosso Estado Program (processes Nos. E-26/202.803/2018 and E-26/201.083/2022) and Prociência scholarship from UERJ. JDBF received Post- Doctoral fellowship from Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro – FAPERJ. and MVS (Process No 307773/2008-6) received research grants from the Conselho Nacional do Desenvolvimento Científico e Tecnológico – CNPq.
References
-
Pimm SL, Jenkins CN, Joppa LN, Roberts DL, Gareth J Russell (2010) How Many Endangered Species Remain to Be Discovered in Brazil?. Natureza & Conservação 8(1): 71-77.
-
Joppa LN, Roberts DL, Pimm SL (2010) How many species of flowering plants are there?. Proc R Soc B 278: 554-559.
-
Uetz P, Freed P, Aguilar R, Hošek J (2022) The Reptile Database. Electronic database.
-
Living Planet Report (2020) Bending the curve of biodiversity loss. In: Almond REA, Grooten M, et al. (Eds.), WWF, Gland, Switzerland.
-
Morellato LPC, Haddad CFB (2000) Introduction: The Brazilian Atlantic Rainforest. Biotropica 32: 786-792.
-
Myers N, Mittermeier RA, Mittermeier CG (2000) Biodiversity hotspots for conservation priorities. Nature 403: 853-858.
-
Lewinsohn TM, Prado PI (2002) Biodiversidade Brasileira. Síntese do Estado Atual do Conhecimento. Contexto, São Paulo.
-
Gans C (1969) Amphisbaenians, reptiles specialized for a burrowing existence. Endeavour 28: 146-151.
-
Castro Mello C (2003) Nova espécie de Bronia Gray, 1845 do estado do Tocantins, Brasil (Squamata: Amphisbaenidae). Papéis Avulsos de Zoologia 43: 139- 143.
-
Hoogmoed MS, Mott T (2003) On the identity of Amphisbaena hugoi Vanzolini, 1990 (Reptilia: Squamata: Amphisbaenidae). Zoologische Mededelingen 77: 455- 457.
-
Gans C (2005) Checklist and bibliography of the Amphisbaenia of the world. Bulletin of the American Museum of Natural History 289: 1-130.
-
Thomas R, Hedges B (2006) Two new species of Amphisbaena (Reptilia: Squamata: Amphisbaenidae) from the Tiburon Peninsula of Haiti. Caribbean Journal of Science 42: 208-219.
-
Mott T, Rodrigues MT, Freitas MA, Silva TFS (2008) New species of Amphisbaena from Bahia, Brazil with a tuberculate non-autotomic tail. Journal of Herpetology 42: 172-175.
-
Ribeiro SL, Vaz-Lima W, Santos AP (2008) New pored Leposternon (Squamata, Amphisbaenia) from Brazilian Cerrado. Zootaxa 1930: 18-38.
-
Ribeiro S, Castro-Mello C, Nogueira C (2009) New species of Anops Bell, 1833 (Squamata, Amphisbaenia) from Jalapão region in the Brazilian Cerrado. Journal of Herpetology 43: 21-28.
-
Strüssmann C, Mott T (2009) Sympatric amphisbaenids from Manso Dam region, Mato Grosso State, Western Brazil, with the description of a new two-pored species of Amphisbaena (Squamata, Amphisbaenidae). Studies on Neotropical Fauna and Environment 44: 37-46.
-
Guedes TB, Entiauspe-Neto OM, Henrique C, Costa HC (2023) Lista de répteis do Brasil: atualização de 2022. Herpetologia Brasileira 12(1): 56-161.
-
Gans C (1966) Studies on amphisbaenids (Amphisbaenia: Reptilia) 3. The small species from southern South America commonly identified as Amphisbaena darwini. Bulletin of the American Museum of Natural History 134: 185-260.
-
Gans C (1971) Studies on amphisbaenians (Amphisbaenia: Reptilia). 4. A review of the amphisbaenid genus Leposternon. Bulletin of the American Museum of Natural History 144: 379-464.
-
Rocha CFD (1992) Geographic distribution: Amphisbaena alba. Herpetological Review 23: 89.
-
Rocha CFD, Bergallo HG, Pombal JP, Geise L, Van Sluys M, et al. (2004a) Fauna de anfíbios, répteis e mamíferos do Estado do Rio de Janeiro, sudeste do Brasil. Publicações Avulsas do Museu Nacional, Rio de Janeiro 104: 1-24.
-
Suguio K, Tessler MG (1984) Planícies de cordões litorâneos quaternários do Brasil: origem e nomenclatura. In: Lacerda LD, Araújo DSD, et al. (Eds.), Restingas: Origem, Estrutura e Processos, pp. 15-25. Centro Editorial da Universidade Federal Fluminense, Niterói.
-
Nimer E (1972) Climatologia da região sudeste do Brasil. Revista Brasileira de Geografia 34: 3-38.
-
Nimer E (1979) Climatologia do Brasil. IBGE, Rio de Janeiro. In: Oliveira JC, Gonzalez RC, et al. (Eds.), Non- Avian Reptiles of the state of Rio de Janeiro, Brazil: status of knowledge and commented list. Papéis Avulsos de Zoologia, 60: e20206024.
-
IBAM/DUMA/PCRJ/SMAC (1998) Guide to Rio de Janeiro’s Environmental Protection Areas. Municipal Secretariat for the Environment, Rio de Janeiro.
-
Gans C, Alexander AA (1962) Studies on amphisbaenids (Amphisbaenia, Reptilia). 2. On the amphisbaenids of the Antilles. Bulletin of the Museum of Comparative Zoology 128: 65-158.
-
Vanzolini PE (1991a) Two new small species of Amphisbaena from the fossil dune field of the Middle Rio São Francisco, State of Bahia, Brazil (Reptilia, Amphisbaenia). Papéis Avulsos de Zoologia 37: 259-276.
-
Levington AE, Gibss RH, Heal E, Dawson CE (1985) Standards in herpetology and ichthyology: Part I. Standard symbolic codes for institutional resource collections in herpetology and ichthyology. Copeia 1985: 802-832.
-
Vanzolini PE (2002) An aid to the identification of the South American species of Amphisbaena. (Squamata, Amphisbaenidae). Papéis Avulsos de Zoologia 42: 351- 362.
-
Gans C (1963) Notes on amphisbaenids (Amphisbaenia, Reptilia). 9. Redescription of Amphisbaena ridleyi Boulenger. Copeia 1: 182-187.
-
Gans C (1964) Redescription of Amphisbaena dubia Müller (Amphisbaenia: Reptilia). Notes on amphisbaenids, 12. Breviora 205: 1-11.
-
Gans C, Amdur M (1966) Redescription of Amphisbaena vermicularis Wagler, with comments on its range and synonymy (Amphisbaenia: Reptilia). Notes on amphisbaenids 20. Proceedings of the California Academy of Science ser 4(33): 69-90.
-
Vanzolini PE (1991b) Two further new species of Amphisbaena from the semi-arid northeast of Brasil (Reptilia, Amphisbaenia). Papéis Avulsos de Zoologia 37: 347-361.
-
Rodrigues MT (2002) Herpetofauna of Quaternary sand dunes of the middle Rio São Francisco, Bahia, Brazil. VIII. Amphisbaena arda sp. nov., a fuliginosa-like checkered patterned Amphisbaena (Squamata, Amphisbaenidae). Phyllomedusa 1: 51-56.
-
Gans C (1962) Notes on amphisbaenids (Amphisbaenia: Reptilia). 5. A redefinition and a bibliography of Amphisbaena alba Linne´. American Museum Novitates 2105: 1-31.
-
Rocha CFD, Van Sluy MS, Vrcibradic D, Hatano FH, Galdino CAC, et al. (2004b) A comunidade dos répteis da Restinga de Jurubatiba. In: Rocha CFD, Esteves FA, et al. (Eds.), Pesquisas de Longa duração na Restinga de Jurubatiba: Ecologia, História Natural e Conservação, Rima, São Carlos, pp: 180-198.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind