The Sperm Ultrastructure of the Nitidulidae Meligethes flavimanus (Stephens, 1830) and of the Phalacridae Olibrus affinis (Sturm. 1807) (Coleoptera Cucujoidea)
The sperm ultrastructure of two members of Cucujoidea was described. The Nitidulidae Meligethes flavimanus sperm are long cells extending along the cyst cell with a series of helicoidal tracts. Each sperm cell has a bi-layered acrosome, a cylindrical nucleus and two mitochondrial derivatives of unequal sizes, the one on the right side larger than the opposite one. The axoneme has the common 9+9+2 microtubule pattern and is flanked by two accessory bodies. The one on the right side has an extension of electron-dense material (a puff-like body) reaching the smaller mitochondrial derivative. The cytoplasm of the sperm cyst hosts numerous bacteria. The sperm of the Phalacridae Olibrus affinis are very long cells that form numerous foldings throughout the cyst. The sperm cell has a long bi-layered acrosome, a cylindrical nucleus and a conventional 9+9+2 axoneme. As in the previous species, two mitochondrial derivatives of different sizes are present, the one on the right side larger than the second one, only 1/3 of the other. Two accessory bodies are on both sides of the axoneme. That on the right side is very large, triangular shaped, while on the opposite side a small electron-dense dot is present. The study performed on the two species of Cucujoidea confirms a close phylogenetic relationship between this superfamily and those of Chrysomeloidea and Curculionoidea, a finding also supported by molecular data.
Introduction
Coleoptera is the most comprehensive insect order with more than 350.000 known species [1, 2, 3, 4, 5]. It was calculated that about 25% of all described animal species is constituted by beetles. The evolution of the group was estimated to occur in the earliest Permian with the divergence among beetle series occurring during the Triassic. Moreover, most superfamilies appeared during Jurassic and almost 64% of families present in the Cretaceous. This origin corresponds to the rise of angiosperms which favoured beetle diversification [6]. According to McKenna, et al. [7] the first beetle pollinators of gymnosperms and early angiosperms, including some early Cucujoidea and Phytophaga, appeared by the late Jurassic, before bees and butterflies. Cucujoidea is a vast superfamily of beetles; the current classification recognizes 37 families extremely diverse and taxonomically difficult [8, 9, 10, 11]. According to molecular data, this superfamily is closely related to Chrysomeloidea and Curculionoidea that are recovered as sister taxa (Phytophaga) [6]. Tenebrionoidea
and Lymexyloidea are recovered together and Cleroidea plus Tenebrionoidea represent the novel hypothesis for cucujform superfamilial relationships [12]. Cleroidea, however, share many characters with certain groups of Cucujoidea such that these two superfamilies are difficult to separate [4, 13]. The superfamily Cucujoidea includes the families of Nitidulidae, also known as sap beetles, and the Phalacridae, also known as shining mold beetles. Nitidulidae is among the more ecologically diverse and species-rich families of Cucujoidea with 4500 species classified in c.351 genera [14]. The Nitidulidae series was suggested to be the earliest-diverged core of Cucujoid lineage [12]. The sperm ultrastructural studies on these two families represent the first series of observations on the superfamily allowing a comparative analysis with the currently available ultrastructural findings on the sperm structure from related groups such as Curculionoidea and Chrysomeloidea [15, 16, 17].
Material and Methods
Specimens Several males and females of Meligethes flavimanus were captured near Siena, on Crataegus sp (Rosaceae), while only three females and two males of Olibrus affinis were found on Asteraceae flowers near Mt. Amiata (Siena) Light microscopy Bundles of sperm of Meligethes flavimanus were isolated from deferent ducts of adult males. The material was placed over a glass in a small drop of 0.1 M phosphate buffer (pH 7.2) to which 3% of sucrose was previously added (PB). The sperm bundles or fragments of the last part of deferent ducts were squashed with a coverslip and observed and photographed using interference contrast with a Leica DMRB microscope equipped with AxioCam MRC5 HR camera (Carl Zeiss).
Transmission Electron Microscopy (TEM)
For TEM observations, adult males of M. flavimanus and Olibrus affinis were dissected in PB to isolate the testes and deferent ducts. The material was fixed overnight in 2.5% glutaraldehyde in PB. After careful rinsing, the material was post-fixed in 1% osmium tetroxide for 2 h. After rinsing, the material was dehydrated with ethanol series (50%- 100%), then transferred to propylene oxide and finally embedded in an Epon-Araldite mixture of resins. We obtained semithin sections of O. affinis samples, with an ultramicrotome Reichert Ultracut, which we stained with 0.1% toluidine blue and observed with a Leica DMRB interference light microscope equipped with an AxioCam MRC5 HR camera (Carl Zeiss). Ultrathin sections were obtained with a Reichert Ultracut, routinely stained with uranyl acetate and lead citrate and observed in a TEM Philips CM10 operating at electron accelerating voltage of 80 kV.
Results
Meligethes flavimanus (Nitidulidae)
The sperm of M. flavimanus is a relatively long cell (about 180 µm long) (Figure 1A) forming a complex of approximately 500 units in each germ cyst (Figure 2A). In longitudinal section sperm cells extend along the cyst volume with a series of helicoidal tracts (Figures 2B & 2C). Quite interesting, the cytoplasm of cyst is filled with many bacteria (Figures 2A & 2B).
In early spermatids, the nucleus, 1.1 µm in diameter, shows two small infoldings at the level of which the layer of microtubules surrounding the structure is missing (Figure 3A). The chromatin, uniformly distributed in the main part of the nucleus, becomes denser at these levels. Beneath the nuclear region an amount of electron-dense material of the centriole adjunct, is evident; within this material a centriole is visible (Figure 3B-3D). On the opposite side of the centriole, a large mitochondrial derivative is present. This structure is characterized by the presence of a large electron-dense inclusion, 0.23 µm wide (Figure 3C,3F). Further the nuclear chromatin becomes distributed in several patches visible in the nucleoplasm (Figurea 3E & 3F). Spermatid cells, in a cross section, have a bi-layered acrosome with perforatorium, 0.4 µm wide (Figure 3E).
The axoneme has the classic 9+9+2 microtubular pattern (Figure 4D). The axoneme is accompanied by two mitochondrial derivatives. The one on the right side is elliptic and larger than the opposite one (0.61 µm X 0.31µm). This mitochondrion shows a globular density in the matrix. Both the structures are surrounded by a layer of microtubules. At this stage of spermiogenesis the accessory bodies are not yet visible (Figure 4A). These structures become evident further and they are visible flanking on both sides the axoneme (Figure 4B,4D). The accessory body on the right side is slightly smaller than the opposite one. This accessory body, however, expands towards the axial region of the cell with a thin electron-dense material. Beneath the accessory bodies, two asymmetrical mitochondrial derivatives are present (Figure 4B,4D). In cross section the one on the right side is larger and circular than the opposite one and it has an electron-dense matrix, while the second mitochondrial derivative is elliptic and shows a moderately electron-dense matrix (Figure 4D). The structures are still surrounded by a layer of microtubules. At the end of spermiogenesis, the sperm cells have a cylindrical shape with an apical bi- layered conical acrosome about 3.0 µm long with an inner perforatorium (Figure 5A). The posterior acrosome region is hosted in an infolding of the apical nuclear region (Figure 5A). The cylindrical nucleus is 0.4-0.5 µm in diameter.

Figure 1: Meligethes flavimanus. A) A long sperm bundles after interference contrast microscope, with many sperm (sp) anchored at the apical region by residual cyst cell (cy), and free flagella (F). B) Several sperm bundle still contained in the cyst cells. The sperm flagella are tightly connected and show evident windings (arrows). In the inset, a detail of the apical region of a sperm cyst to show the twisting of inner sperm. Note the apical nuclear shining region (arrowheads).
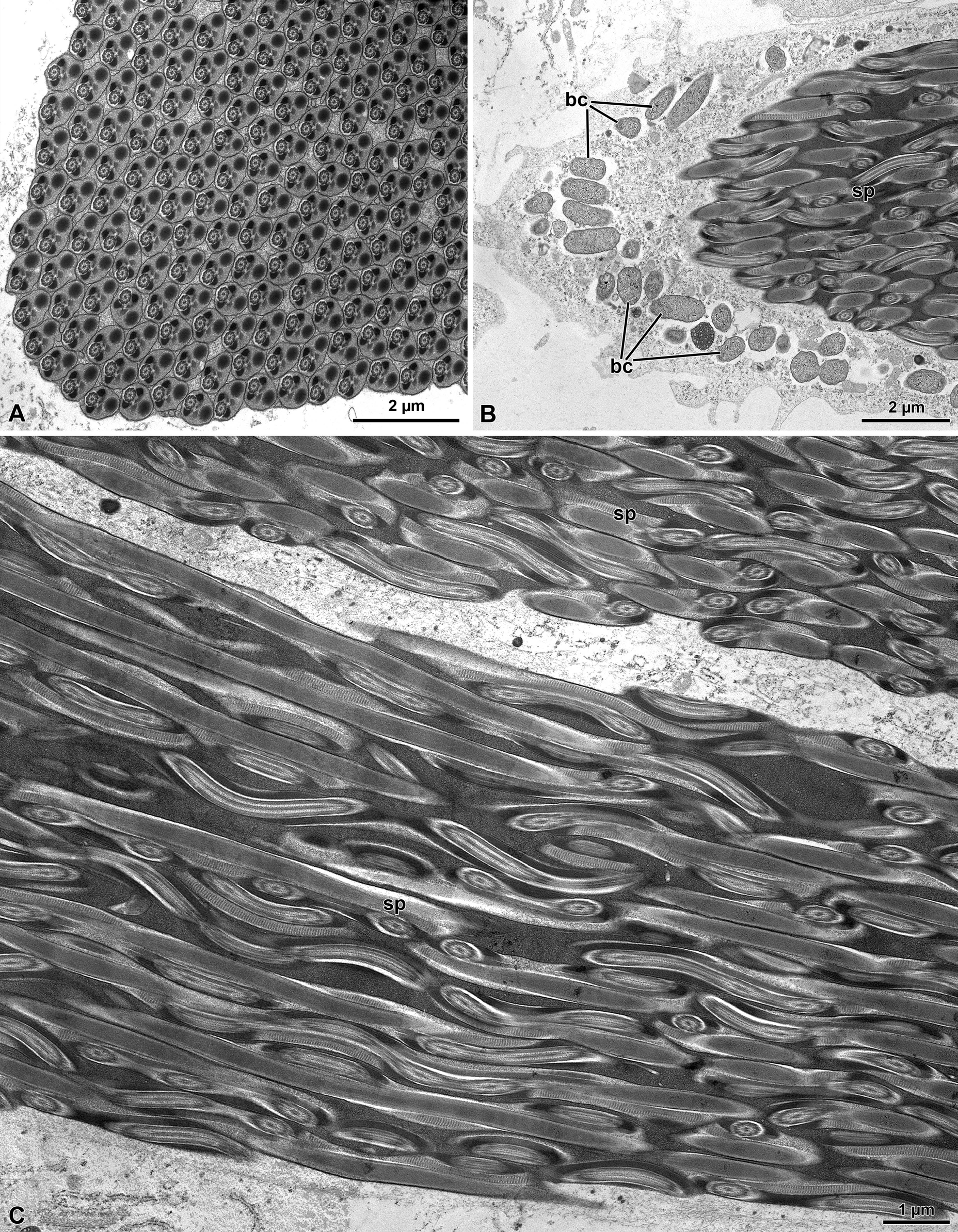
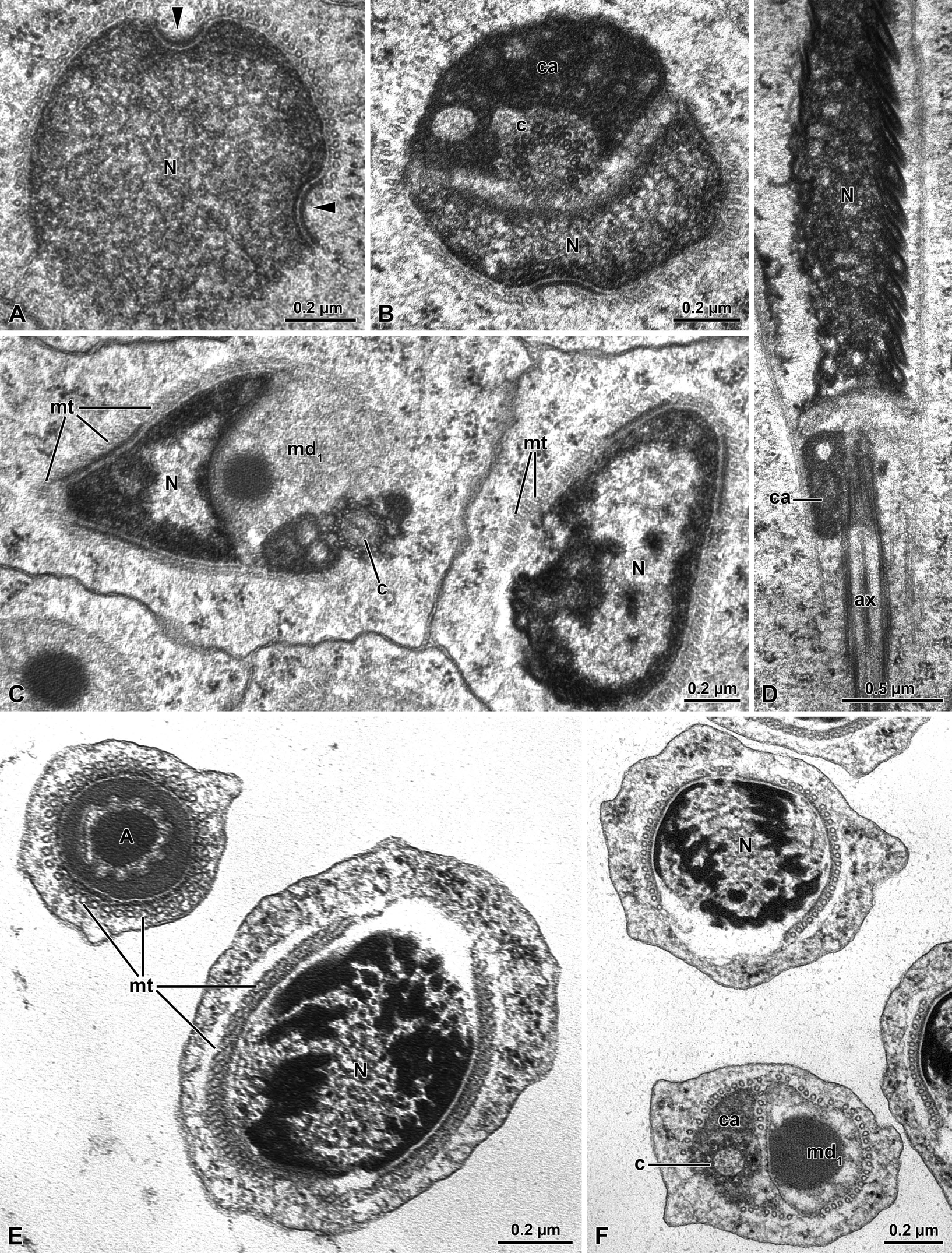
Figure 3: Meligethes flavimanus. A) Cross section through the early nucleus (N) showing the two opposite infoldings of the nuclear envelope (arrowheads). Note the uniform density of chromatin. B) Cross section through the centriolar region with a remnant of nucleus (N), the large centriole adjunct material (ca) surrounding the centriole (c). C) Cross section of the basal nuclear region with the nucleus (N) the centriole (c) and the large mitochondrial derivative (md). Note the layer of microtubules (mt) surrounding the spermatid nucleus. D) Longitudinal section through the spermatid nucleus (N) with the centriole surrounded by the centriole adjunct material (ca) and the axoneme (ax). E) Cross section of still early spermatids with the acrosome (A) and the nucleus (N) surrounded by a layer of microtubules (mt). F) Cross section of early spermatids with the nucleus (N) and the centriolar region with the centriole (c) surrounded by the centriole adjunct material (ca) and the large mitochondrial derivative (md1).
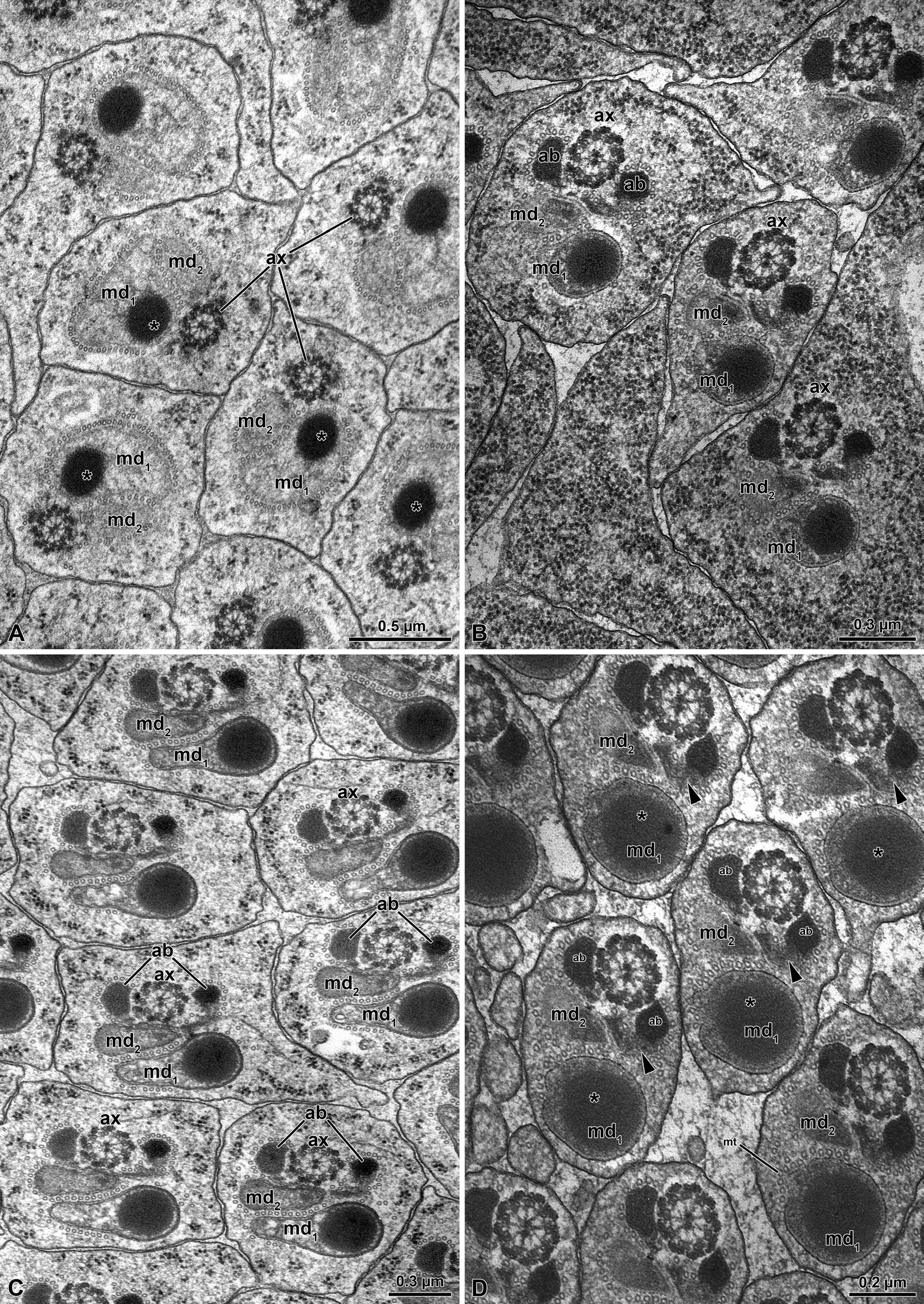
Figure 4: Meligethes flavimanus. A) Cross section of early spermatid flagella with the axoneme (ax), the two unequal mitochondrial derivatives (md1 and md2), the larger one (md1) containing an electron-dense globular material (asterisk). Note that accessory bodies are not yet visible. B-C) Cross section through advanced spermatids with the two unequal mitochondrial derivatives (md1 and md2) and the two accessory bodies (ab) on both sides of the axoneme (ax). D) Cross section of almost mature spermatozoa with the two unequal mitochondrial derivatives (md1 and md2), the larger one (md1) filled with electron-dense material (asterisk) and the right accessory body (ab) with an extension of electron-dense material (arrowhead). A layer of microtubules (mt) still surrounds the structures.
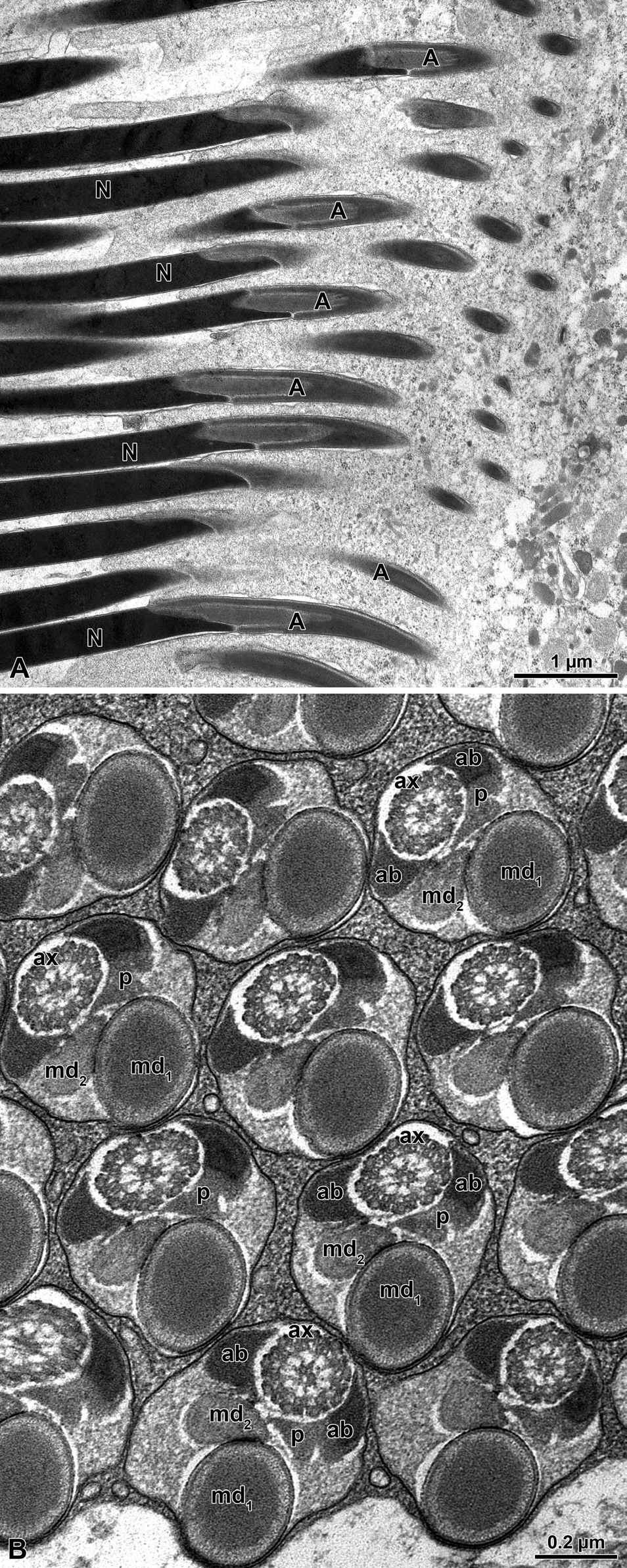
Figure 5: Meligethes flavimanus. A) Longitudinal section through mature sperm. A, acrosome; N, nucleus. B) Crosssection through sperm flagellar axoneme (ax) with the two unequal mitochondrial derivatives (md1 and md2), the two similar accessory bodies (ab); that on the right side is provided with a puff-like body (p). Note also that the position of the large mitochondrial derivative and the axoneme are on the same longitudinal axis of the flagella.
The sperm flagella (Figure 5B) have an elliptical shape in cross section with the large mitochondrial derivative placed in the deeper region of the cross section, opposite to the axoneme. The axoneme is flanked by the two triangular accessory bodies, with that on the right side showing an extension of electron-dense material reaching with its apex the smaller mitochondrial derivative. The tail end is characterized by the lack of accessory bodies and mitochondrial derivatives and by the disorganization of the microtubular pattern.
Olibrus affinis (Phalacridae)
The sperm of O. affinis is are quite long cells (about 280- 300 µm long) assembled in numerous elements in large elongated cyst cells which can reach the huge dimensions of 2.0 µm X 1.8 µm (Figures 6A-6E). The number of sperm per cysts can be variable, but on average they consist of about 500 units (Figure 8A) as the results of 29 cell divisions of early germ cells. Due to the sperm length and the reduced volume of the cyst cells, the sperm run in sinuous way throughout the cyst, forming several loops (Figures 6A-6E).
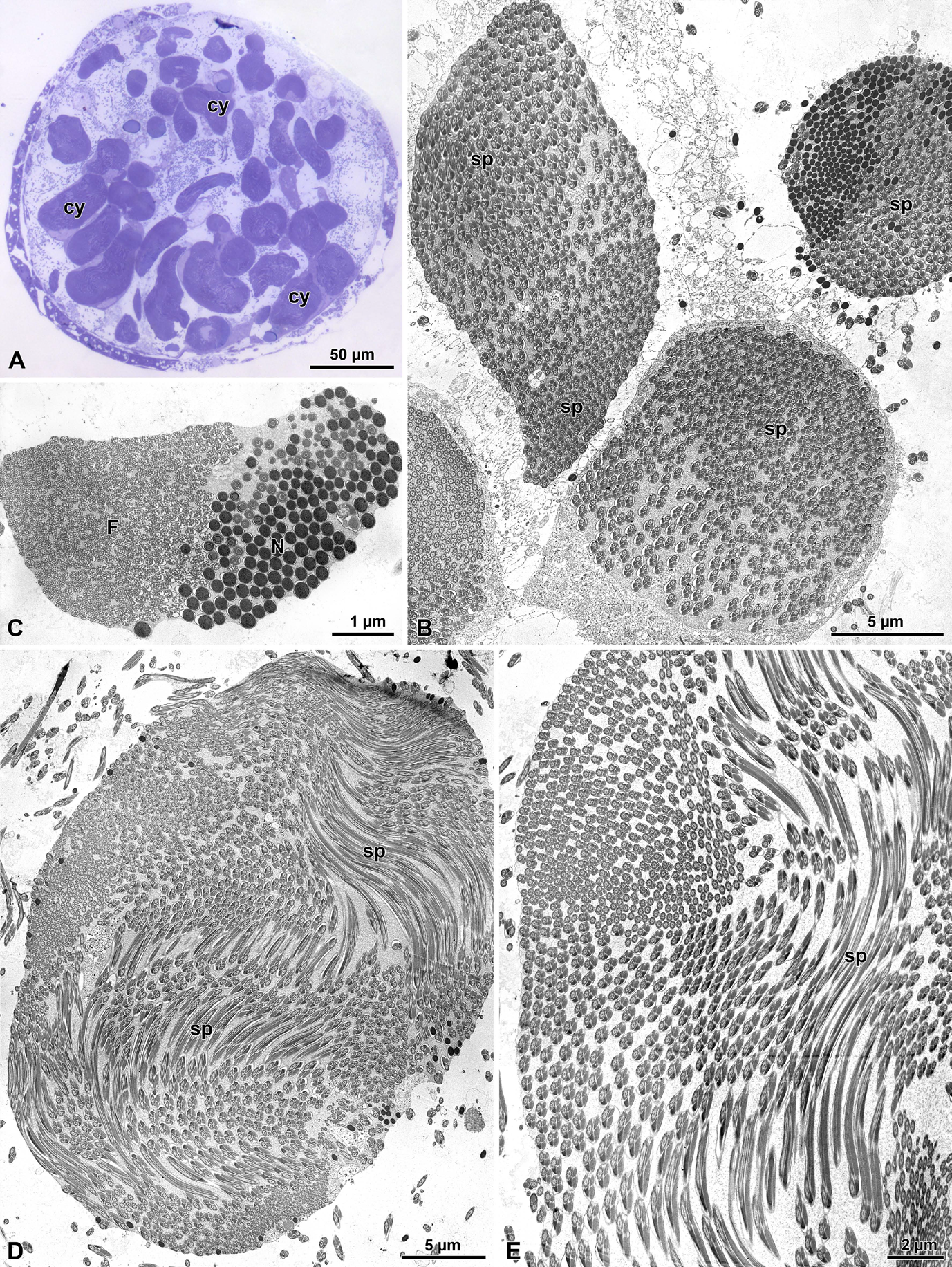
Figure 6: Olibrus affinis. A) Semi-thin section through testis showing the large sperm cysts (cy). B) Cross section at low magnification showing the large number of sperm cells (sp) in each cyst. C) Cross section through a cyst to show the closeness of nuclei (N) and tail ends (F) as consequence of sperm looping. D-E) Cross sections of two large sperm cysts to show the sperm looping (sp) throughout the cyst volume.
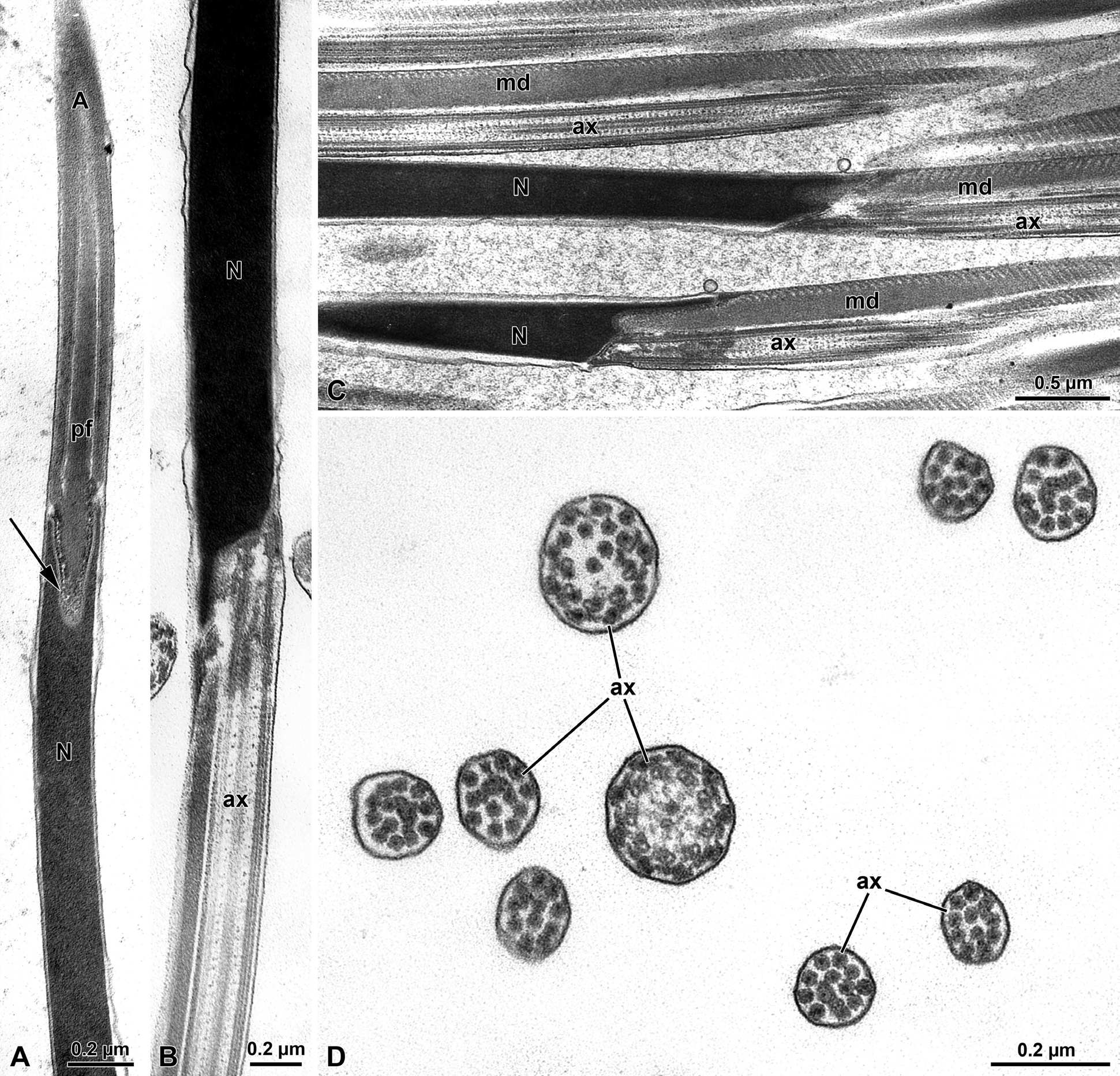
Figure 7: Olibrus affinis. A) Longitudinal section of the apical sperm with the pointed long acrosome (A) provided with the inner perforatorium (pf). Note the insertion of the basal region of the acrosome hosted in a deep nuclear (N) infolding (arrow). B-C) Longitudinal section through the posterior nuclear region (N) showing the infoldings to host the axoneme (ax) and the mitochondrial derivative (md). D) Cross section through the tail end showing the disappearance mitochondrial derivatives and accessory bodies. The flagellar axoneme (ax) is disorganized and singlet microtubules are evident.
The sperm show a long apical bi-layered acrosome, 1.52 µm with an inner perforatorium. The basal region is hosted in a deep nuclear infolding (Figure 7A). The nucleus, about 25 µ long, is a cylindrical structure, 0.18 µm wide (Figures 7A-7C & 8A). The basal nuclear region shows an infolding to adapt a mitochondrial derivative and, on the opposite side, the centriole (Figures 7B,7C) surrounded by a scant centriole adjunct material. The centriole has the conventional structure and is devoid of the central microtubules but has already the 9 outer accessory tubules instead of the triplets of microtubules (Figure 8C). The axoneme has the common structure of 9+9+2 microtubular pattern for most of the flagellar length (Figures 8C,8D). At the tail end, which has reduced its width to 1.6 µm in diameter (Figures 7D & 8B), the axoneme loses its normal configuration, the microtubules becoming disorganized with only singlet microtubules visible. The flagellar axoneme is accompanied by two mitochondrial derivatives of different sizes. The one on the right side is clearly larger and shows a matrix with an evident crystallization, while the second mitochondrial derivative is smaller, about 1/3 of the opposite one (Figures 8C,8D). This different shape of the mitochondrial derivatives was quite evident in cross section of the initial flagellum and for most of its length, but further the two structures became almost of the same size or with the right mitochondrial derivative slightly larger than the opposite one. These structures are no longer visible at the flagellar end (Figure 7D). On both sides of the axoneme, two accessory bodies are visible in cross section. That on the right side is very large, triangular shaped (0.16 µm X 0.13µm), while that on the opposite side is very small and looks like electron-dense dot, only 33 nm wide in cross section (Figures 8C,8D).
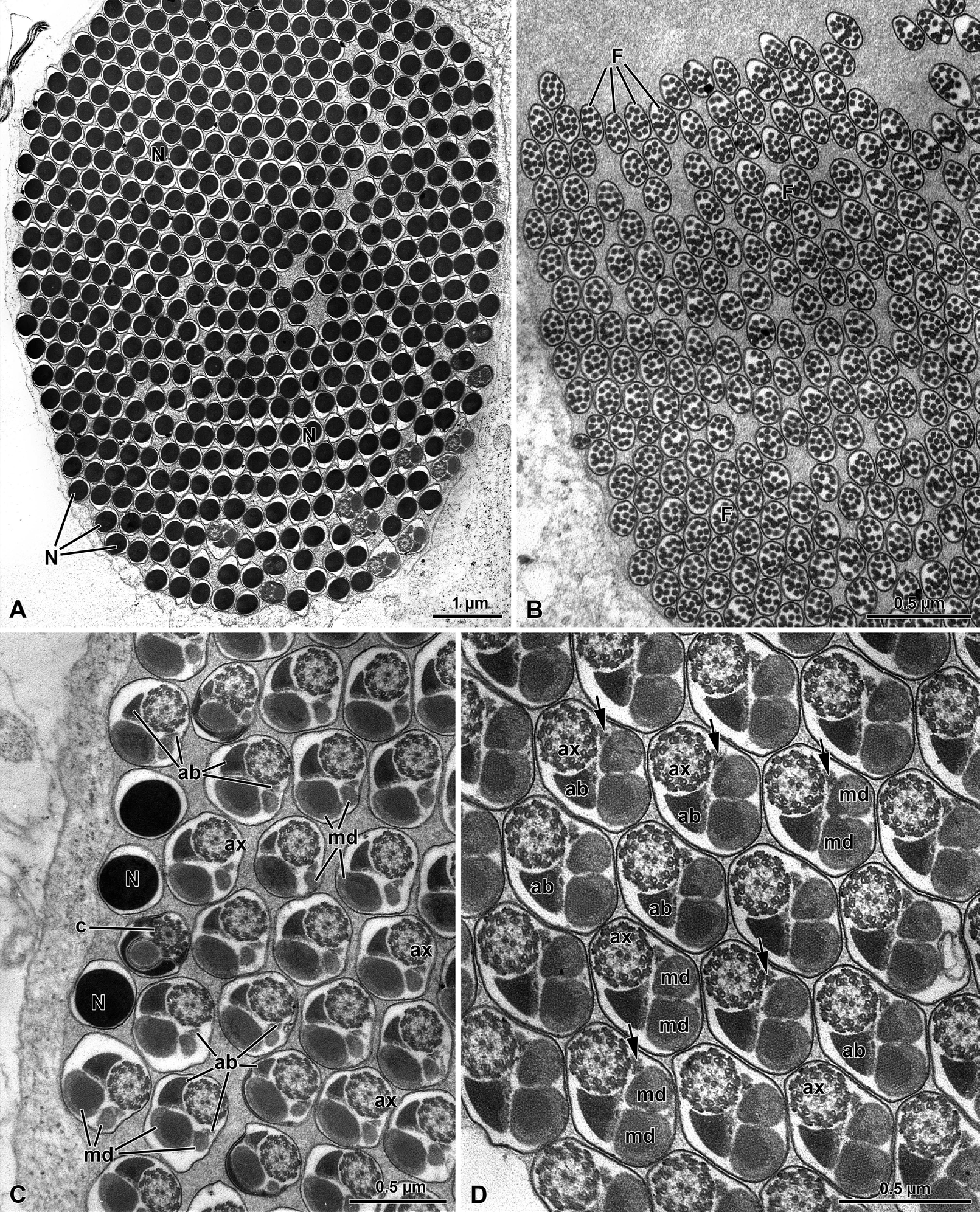
Figure 8: Olibrus affinis. A-B) Cross sections through sperm cyst to show the numerous sperm. Note in Fig. A, the nucleus (N) and in Fig. B, the tail end (F). C) Cross section through sperm flagella. At this level the two mitochondrial derivatives (md) are different in size and shape. as well as the two accessory bodies (ab). ax, axoneme; N, nucleus; c, centriole. D) Cross section of the sperm flagella at a deeper level, with the two mitochondrial derivatives (md) of approximately the same size and the two accessory bodies (ab) very different in size with the right one quite small (arrows). ax, axoneme.
Conclusion
The general sperm ultrastructure of both Cucujoidea families studied in the present work indicates is similar to that of the superfamilies Curculionoidea and Chrysomeloidea. This confirms that all these superfamilies share sperm features typical of the section Cucujiformia. On the contrary, the sperm features appear to be quite different from those of members of already studied [1, 15] and [17, 18]. Thus the complex of characters dealing with sperm morphology confirms that sperm is a valid instrument to provide valuable information for researches on insect phylogeny among species within the superfamily. [19, 20, 21]. The most important character suggesting the occurrence of a relationship between the mentioned groups is the presence, in the sperm flagellum, of two accessory bodies of different sizes. The structure of the accessory bodies is one of the most important features among Coleoptera [19]. Several works on different groups of beetles have established that the number, shape and structural organization of the accessory bodies can be variable, depending on the amount of the material of the centriole adjunct; it is well known that these structures are in continuity with this material [19, 20, 21, 22] and extend along the sperm flagellum on both sides of the axoneme. In the species studied here the two accessory bodies have a different shape. In the Phalacrid O. affinis one accessory body is extremely small, while the opposite one is uncommonly large. Such different in size of the two structures could be due to the scant development of the centriole adjunct material at the centriolar level. Differently, in the Nitidulid M. flavimanus the two accessory bodies are slightly different in size, with the smaller one showing an extension towards the axial region of the sperm flagellum. Such extension is quite evident in the Curculionoidea where it was defined as a “puff-like body”, apparently consisting of part of the material of the accessory body, but displaying a different appearance [16, 17]. A similar structure was also described in sperm of Chrysomeloidea [16, 17]. This similarity between the structure of the accessory bodies in members of Chrysomeloidea and in the species of Nitidulidae must be carefully considered in order to verify whether the same evidence is present in other species of the family. Another important characteristic observed in the species here examined is the presence of two mitochondrial derivatives of different sizes. That on the right side is larger than the opposite one. The presence of such unequal size of the two mitochondrial derivatives is an important marker distinguishing members of the Cucujoidea from those of the superfamilies Cleroidea and Tenebrionoidea which are characterized by two identical mitochondrial derivatives either of regular or small size [22, 23] or giant structures as it occurs in Divales sp. [24]. The asymmetry of the mitochondrial derivatives, with the largest mitochondrion placed, in a flagellum cross section, in the opposite position of the axoneme is also typical of the sperm of Chrysomeloidea and Curculionoidea [16, 17, 25]. The large mitochondrial derivative also has a large crystallization of its matrix, while the smaller mitochondrion has not. Finally, the two species studied here exhibit large cyst cells containing a great number of sperm. This finding was already observed in several beetles and according to Virkki [26] such a feature would be indicative of a archaic phylogenetic position of the species exhibiting this result. A further consideration deals with the large number of sperm cells in each cyst showing numerous sperm tail convolutions. This feature is presumably an adaptation to host numerous and very long sperm in a narrowed cyst volume. This gives rise to multiple folds of the sperm flagella with evident numerous loops determining the positioning of sperm heads close to flagellar ends. We consider noteworthy to mention that the cytoplasm of the cyst cells hosts numerous bacteria. The presence of bacteria in insect tissue is not a rare event. It is known that many insects have intracellular symbiotic bacteria that provide them with essential nutrients [27]. However, many insects rely such symbionts within specialized host cells, the bacteriocytes. In the testes of the Nitidulid species here studied, on the contrary, the bacteria are present within the cytoplasm of cyst cells. Whether their presence can give some benefits to the sperm development within the cyst is unknown. In conclusion, from the study performed on the two species of Cucujoidea we can conclude that the sperm morphology has confirmed the occurrence of a close phylogenetic relationships of this superfamily with those of Chrysomeloidea and Curculionoidea, as also supported by molecular data [6, 7]. The peculiarity dealing with the accessory bodies of extremely different sizes in the member of the family Phalacridae could be due to the different phylogenetic positions of the two families Nemonichidae and Phalacridae which belong to two different clades. Further studies on other members of the family are required to verify this hypothesis.
References
-
Crowson RA (1960) The phylogeny of Coleoptera. Annual Review of Entomology 5: 111-134.
-
Crowson RA (1981) The Biology of Coleoptera Acad. Press Inc, London, UK, pp: 802.
-
Slipinski SA, Leschen RAB, Lawrence JF (2011) Order Coleoptera Linnaeus 1758. In: Zhang Z.-Q. (Edn.), Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, New Zealand Arthropod Collection 3148 (1): 203-208.
-
Lawrence JF, Newton AF (1982) Evolution and classification of beetles. Ann Rev Ecol Syst 13: 261-290.
-
Beutel RG, Leschen RAB (2005) Coleoptera, Beetles volume 1: morphology and systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). In: Kristensen NP, (Ed.), Handbook of Zoology: A Natural History of the Phyla of the Animal Kingdom: Volume IV Arthropoda: Insecta: Part 38. de Gruyter, Berlin, Germany, pp: 11-19.
-
Zhang SQ, Che LH, Li Y, Liang D, Pang H, et al. (2018) Evolutionary history of Coleoptera revealed by extensive sampling of genes and species. Nature Communications.
-
McKenna DD, Shin S, Ahrens D, Balke M, Beza-Beza C, et al. (2019) The evolution and genomic basis of beetle diversity. PNAS 116: 24729-24737.
-
Leschen RAB, Lawrence JF, Ślipiński SA (2005) Classification of basal Cucujoidea (Coleoptera: Polyphaga): cladistics analysis, keys and review of new families. Invertebr Syst 19: 17-73.
-
Leschen RAB, Ślipiński SA (2010) Cucujoidea Latreille, 1802: Introduction, Phylogeny. In: Leschen RAB, Beutel RG, et al. (Eds.), Handbook of Zoology, Vol. IV: Arthropoda: Insecta Part 38. Coleoptera, Beetles, Vol 2: Morphology and Systematics (Polyphaga partim), Walter de Gruyter GmbH & Co., Berlin, Germany, pp: 281-282.
-
Lord NP, Hartley CS, Lawrence JF, McHugh JV, Whiting MF, et al. (2010) Phylogenetic analysis of the minute brown scavenger beetles (Coleoptera: Latridiidae), and recognition of a new beetle family, Akalyptoischiidae fam. n. (Coleoptera: Cucujoidea). Syst Entomol 35(4): 753-763.
-
Cline AR, Smith TR, Miller K, Moulton M, Whiting M, et al (2014) Molecular phylogeny of Nitidulidae: Assessment of subfamilial and tribal classification and formalization of the family Cybocephalidae (Coleoptera: Cucujoidea). Syst Entomol 39(4): 758-772.
-
Robertson JA, Ślipiński A, Moulton M, Shockley FW, Giorgi A, et al. (2015) Phylogeny and classification of Cucujoidea and the recognition of a new superfamily Coccinelloidea (Coleoptera: Cucujiformia). Syst Entomol 40: 745-778.
-
Crowson RA (1955) The Natural Classification of the Families of Coleoptera, Abe Books, Nathaniel Lloyd & Co, London, UK, pp: 187.
-
Jelínek J, Carlton CE, Cline AR, Leschen RAB (2010) Nitidulidae Latreille 1802. In: Leschen RAB, Beutel RG, et al. (Eds.), Handbook of Zoology, Vol. IV: Arthropoda: Insecta Part 38. Coleoptera, Beetles, Vol. 2: Morphology and Systematics (Polyphaga partim), Walter de Gruyter GmbH & Co., Berlin, Germany, pp: 390-407.
-
Baccetti B, Daccordi M (1988) Sperm structure and phylogeny of the Chrysomelidae. In: Jolivet P, Petitpiere E, et al. (Eds.), Biology of Chrysomelidae, Kluwer Academic Publishers, Dordrecht, Netherlands, pp: 357-378.
-
Burrini AG, Magnano L, Magnano AR, Scala C, Baccetti B (1988) Spermatozoa and phylogeny in Curculionidea (Coleoptera). Internat J Morphol Embryol 4: 1-50.
-
Dallai R, Afzelius BA, Lupetti P, Osella G (1998) Sperm structure of some Curculionoidea and their relationship with Chrysomeloidea. Boll Mus Regionale Sci Nat Torino 1: 27-50.
-
Name KPO, Dos Reis GPF, Báo SN (2007) An ultrastructural study of spermiogenesis in two species of Sitophilus (Coleoptera: Curculionidae). Biocell 31(2): 229-236.
-
Jamieson BGM, Dallai R, Afzelius BA (1999) Insects: Their Spermatozoa and Phylogeny. Science Publishers, Enfield, New Hampshire, USA, UK, pp: 555.
-
Dallai R (2014) Overview on spermatogenesis and sperm structure of Hexapoda. Arthropod Struct Dev 43(4): 257-290.
-
Gottardo M, Dallai R, Mercati D, Hörnschemeyer T, Beutel RG (2016) The evolution of insect sperm-an unusual character system in a megadiverse group. J Zool Syst Evol Res 54(4): 237-256.
-
Dias G, Mercati D, Rezende PH, Lino-Neto J, Fanciulli PP, et al. (2022) New findings on the sperm structure of Tenebrionoidea (Insecta, Coleoptera). Insects 13(5): 485.
-
Baccetti B, Burrini AG, Dallai R, Giusti F, Mazzini M, et al. (1973) Structure and function in the spermatozoon of Tenebrio molitor. The spermatozoon of Arthropoda XX. J Mechanochem Cell Motil 2(3): 149-161.
-
Mazzini M (1976) Giant spermatozoa in Divales bipustulatus F. (Coleoptera: Cleridae). Int J Insect Morphol Embryol 5(2): 107-115.
-
Alzahrani AM, Abdelsalam SA, Elmenshawy OM, Abdel- Moneim AM (2013) Ultrastructural characteristics of spermiogenesis in Rhynchophorus ferrugineus (Coleoptera, Curculionidae). Fla Entomol 96(4):1463- 1469.
-
Virkki N (1969) Sperm bundles and phylogenesis. Zeitschift Zellforsch 101:13-27.
-
Masson F, Moné Y, Vigneron A, Vallier A, Parisot N, et al. (2015) Weevil endo-symbiont dynamics is associated with a clamping of immunity. BMC Genomics 16: 819.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework