Redescriptions of Illiosentis ctenorhynchus Cable & Linderoth, 1963 (Illiosentidae) and Micracanthorhynchina motomurai (Harada, 1935) (Rhadinorhynchidae: Gorgorhynchinae) (Acanthocephala: Palaeacanthocephala)
We provide redescriptions of two species of acanthocephalans in one class Palaeacanthocephala and 2 families collected from hosts in different geographical locations in Jamaica and Korea. Each species account is annotated and morphologically presented using light microscopy showing new characteristic diagnostic features. In Illiosentidae, we provide a new description of Illiosentis ctenorhynchus Cable & Linderoth, 1963 from the type host, the yellow goatfish Mulloidichthis martinicus (Cuvier) in Jamaica that completes and corrects the inadequate and incomplete original description. In Rhadinorhynchidae (Rhadinorhynchinae), we redescribe Micracanthorhynchina motomurai (Harada, 1935) from the pale chub Zacco platypus (Temminck & Schlegel) and the dark chub Zacco temmincki (Temminck & Schlegel) in Korea and complement much missing information and images. We also provide museum numbers for lectotype male, allolectope female, and paralectotypes that we have deposited as Harada did not designate type material for M. motomurai.
Omar M Amin* and Nataliya Y Rubtsova
Introduction
We have collected thousands of parasitic organisms from vertebrates over the years as part of an overall research program from North America and elsewhere in the world. Other collections were gifted to us for diagnosis or for joint research efforts. Significant collections were published. A few others were just saved and put aside after having been processed as whole mounts as reference material for future examination. We have started this series of investigations by exploring a group of digeneans, cestodes, and nematodes that have not been previously studied or published [1]. The polymorphid acanthocephalans were the subject matter of the second number of this series [2]. In the third volume, we have described and illustrated acanthocephalans of two classes, Archiacanthocephala and Eoacanthocephala, and noted their host and geographical distribution [3]. In the present volume, we redescribe two species in the Paleacanthocephala that have been poorly described with much missing information that we complement.
Materials and Methods
The specimens reported in this presentation were collected using routine methods for the examination of relevant hosts.
Processing for Microscopy
Specimens were already processed and whole mounted by the donors. Specimens of Illiosentis ctenorhynchus Cable, et al. [4] were placed in tap water until turgid and refrigerated. They were stained with Harris hematoxylin, dehydrated in ascending concentrations of ethanol to absolute alcohol, cleared with methyl salicylate and whole-mounted in terpineol and xylodamar. Specimens of Micracanthorhynchina motomurai were stained in aceto-carmine.
Optical Microscopy
Images were acquired using a Zeiss Axioskop Transmitted Nomarski DIC Phase Contrast Microscope Trinocular (Munich, Germany) and a Canon T3i EOS 600D DSLR Camera (Melville, New York). Images were transferred to a USB and then to a computer for processing and management. Measurements are in micrometers unless otherwise noted; the range is followed by the mean values between parentheses when appropriate. Width measurements represent maximum width.
Results and Discussion
Illiosentis ctenorhynchus Cable and Lindderoth, 1963
• (=Dollfusentis ctenorhynchus (Cable and Linderoth, 1963) Golvan, 1969 (Illiosentidae) (Figs. 1-8) Cable and Linderoth [4] described I. ctenorhynchus from 2 males and one immature female collected from yellow goatfish, Mulloidichthys martinicus (Cuvier) in Jamaica. Golvan placed it in the genus Dollfusentis without sufficient justification. We follow the classification of Amin, et al. [5] which restores the validity of the genus Illiosentis Van Cleave and Lincicome, 1939 with molecular validation.
We describe I. ctenorhynchus from one male and one female and complete deficiencies in the original description. Most measurements are comparable to those that were described in the first description but structures being described for the first time are bolded constituting the significant parts justifying the redescription (Tables 1 & 2).
| Parasite | Host | Distribution |
|---|---|---|
| Family Illiosentidae | ||
| Illiosentis ctenorhynchus Cable & Linderoth, 1963 | Mulloidichthys martinicus (Cuvier), yellow goatfish | Jamaica |
| Family Rhadinorhynchidae | ||
| Micracanthorhynchina motomurai (Harada, 1935) | Zacco temmincki (Temminck & Schlegel) & Z. platypus (T. & S.) | Chonju & Chon Buk, Korea |
Table 1: Two species of Acanthocephala (Palaeacanthocephala) redescribed from wildlife in this paper and their hosts and geograph
General. With essential characteristics of the genus Illiosentis. Worms small, slender, cylindrical, slightly wider anteriorly (Figs. 1, 2). Sexual dimorphism not prominent in length of trunk, dorsal vs. ventral proboscis hooks and trunk spines. Proboscis long, club-shaped, with 12 longitudinal rows of 18 hooks each (Figs. 3-6). Apical hooks 30-35 long, subapical hooks 43-45 long, getting gradually smaller posteriorly with posterior- most 5-7 spine-like hooks crowded and rootless of comparable size, about 15 long. Separated ventral hooks in crescent 40-45 long. Anterior dorsal trunk spines in 3 or 4 rows measuring 25 long in male and 42 long in female. Corresponding anterior ventral trunk spines in 7 rows measuring 25-38 in male and 25-50 in female (Figs. 5, 6). Sensory papillae between hooks 7-9 from posterior not conspicuous. Neck prominent, 125 dorsally X 104 at base. Proboscis receptacle double-walled, about one and half times as long as proboscis with cephalic ganglion somewhat near its anterior end (Figs. 3, 4). Lemnisci digitiform, unequal, markedly longer than receptacle (Fig. 3).
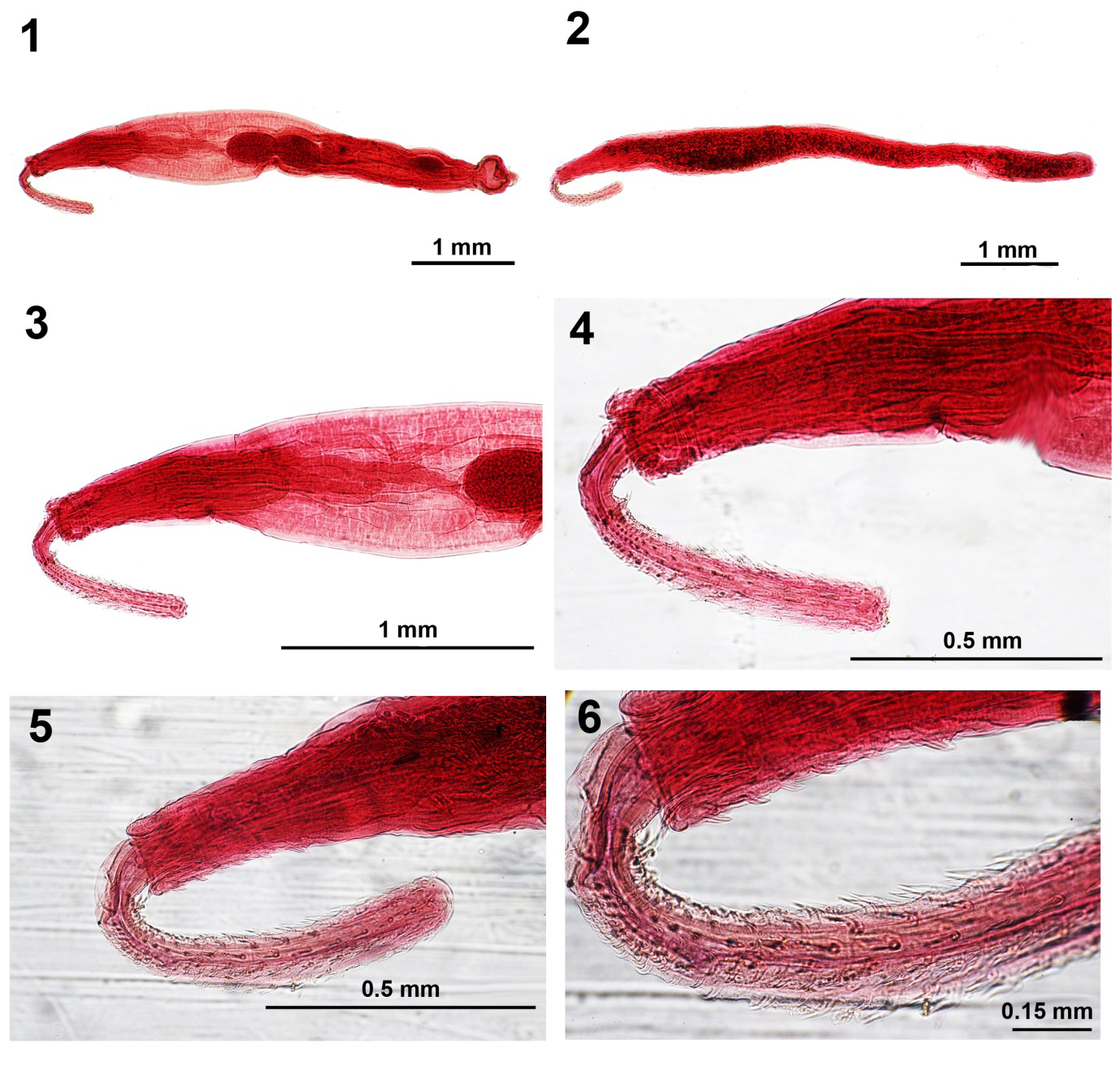
Figures 1-6: Light microscope images of specimens of Illiosentis ctenorhynchus from Mulloidichthys martinicus in Jamaica.
- A whole-mounted male specimen showing the complete reproductive system and the relative sizes of the proboscis, receptacle, and lemnisci.
- The allolectotype female.
- The anterior half of the male specimen showing the proboscis, receptacle, lemnisci, and part of the anterior testis.
- The proboscis of the male specimens.
- The proboscis of the female specimen and some of the trunk spines.
- A higher magnification of Fig. 5 showing more detail of ventral trunk spines and the types of proboscis hooks especially the posterior miniature spine-like hooks.
Male. 3.67 mm long by 0.62 mm wide anteriorly. Proboscis 600 X 100 anteriorly. Neck 125 dorsally X 104 at base. Receptacle 1.07 mm X 0.17 mm. Lemnisci unequal, 1.300 X 0.052 mm and 1.52 X 0.062, considerably longer than receptacle. Testes ovoid, equal, contiguous, post equatorial each measuring 425 X 250 mm (Figs.
3, 7). No space separating reproductive structures. About 6 cement glands with large nuclei, ovoid, 166 X 104. Saefftigen’s pouch medial, bulb-like 520 X 228, contiguous with cement glands. Sperm vesicle 468 X 147, more posterior than and dorsal to Saefftigen’s pouch posteriorly. Bursa (Fig. 7) 385 X 333.
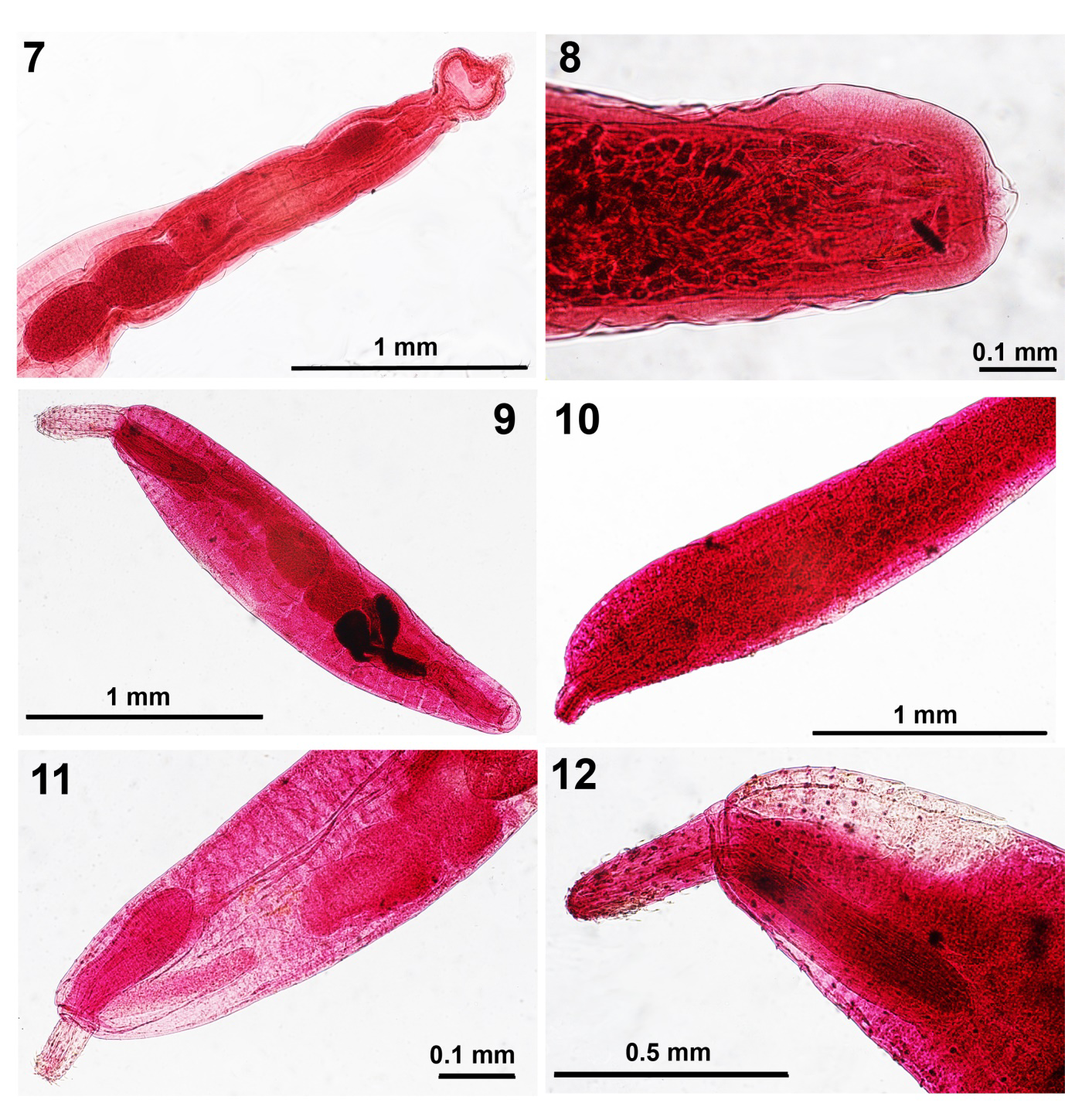
Figures 7 & 8: Light microscope images of specimens of Illiosentis ctenorhynchus from Mulloidichthys martinicus in Jamaica showing the posterior male reproductive system and the posterior end of the female trunk depicting the terminal gonopore. Figs. 9-12. Images of specimens of Micracanthorhynchina motomurai from Zacco temmincki & Zacco platypus in Korea. 9. A whole mounted lectotype male showing fusiform trunk, proboscis, receptacle and reproductive system. 10. The allolectotype female specimen. 11. The anterior half of a male paralectotype specimen showing the corresponding length of the receptacle and lemnisci and the anterior portion of the reproductive system. 12. The anterior portion of a male specimen showing the distribution of dorsal and ventral trunk spines.
Gravid female. Trunk 4.62 mm long by 0.40 mm wide anteriorly. Proboscis 600 X 112 anteriorly. Neck 177 dorsally X 114 at base. Receptacle 884 X 177. Lemnisci and reproductive system obscured by eggs. Gonopore terminal in rounded posterior end (Fig. 8). Eggs ellipsoid with marked polar prolongation of fertilization membrane, 65-80 X 15-23. • Specimens: Cable and Linderoth deposited the holotype male in the Helminthol. Coll. of the US Natl. Museum no. 60344. We have deposited one male and one female allolectotype on one slide (etched with 86-12-20-14) at the Nebraska State Museum’s Harold W. Manter Laboratory (HWML) Parasitology Collection no. HWML- 217503.
• Remarks: Our redescription adds a considerable amount of information that was missing in the original description. These include information about worm size and shape, localization, size and numbers of anterior trunk spines, proboscis shape, armature, hooks and roots sizes, neck and receptacle sizes, shape of lemnisci, testes shape and sizes, number and anatomy of cement glands, shape and measurements of Seafftigen’s pouch, seminal vesicle, and bursa. Information on a gravid female was also provided for the first time since the original description included only one immature female. Our contributions to the redescription of the gravid female designated herein as lectotype female include size of trunk, proboscis, neck, receptacle, the size and anatomy of the egg, and the determination that the female gonopore is terminal.
Micracanthorhynchina motomurai (Harada, 1935) Ward, 1951
• (= Micracanthorhynchus motomurai Harada, 1935; Micracanthocephalus motomurai (Harada, 1935) Harada, 1938 (Rhadinorhynchidae: Gorgorhynchinae) (Figs. 9-16) Van Cleave and Lincicome [6] established their family Gorgorhynchidae (now Subfamily Gorgorhynchinae) for rhadinorhynchid species with 4 cement glands as opposed to species in Rhadinorhynchidae in the current subfamily Rhadinorhynchinae with 8 cement glands. Exceptions are known. Micracanthorhynchina motomurai was first described by Harada [7] as Micracanthorhynchus motomurai but the genus name was preoccupied and the species name changed to Micracanthorhynchina motomurai by Ward [8] irrespective of changing it earlier to Micracanthocephalus motomurai by Harada [9].
We have been gifted 12 specimens of M. motomurai (8 males & 4 females) by the Chon Buk University in Chon Buk, Korea. The original description included a few line drawings of a whole male, two hooks, and an inaccurate female reproductive system that were repeated by Petrochenko [10] and Golvan [11], among other observers. Drawings of a whole female, a row of hooks with roots, and eggs were missing. We provide a qualitative and metric redescription of the species in which we included measurements of lemnisci, testes, and cement glands, among others, that were missing in the original description (Table 2).
| Authority | Harada (1935, 1938), Glovan (1969) | Redescription (this paper) |
|---|---|---|
| Hosts | Zacco temmincki (Temminck & Schlegel) & Z. platypus (T. & S.) & Chanodichthys macropus Günther | Zacco temmincki (Temminck & Schlegel) & Zacco platypus (Temminck & Schlegel) |
| Locality | Korea & Taiwan | Chonju & Chon Buk, Korea (Chon Buk University) |
| Sample size | ---- | 8 males, 4 females |
| Cuticle | With many fragmented nuclei | With many fragmented nuclei |
| Lacunar system | 2 lateral longitudinal canals & transverse branches looking as pseudo-segmented | 2 lateral longitudinal canals & transverse branches looking as pseudo-segmented |
| Trunk (males) L X W (mm) | Fusiform: 1.6-3.5 X 0.35-0.50 | Fusiform: 2.05-3.87 X 0.45-0.90 |
| Trunk (females) L X W (mm) | Fusiform: 3.2-4.5 X 0.50-0.60 | Fusiform: 2.05-3.00 X 0.55-0.80 |
| Ant. trunk spines | 8-11 dorsal & 18-22 ventral | Males: 7-9 dorsal & 15-19 ventral* Females: 8-13 dorsal & 17-21 ventral |
| Spine length (μm) | 20 | 15-21. Larger ventrally & in females |
| Proboscis L X W (μm) | Club-shaped: 300-500 X 150 | Club-shaped: 312-416 X 125-166 |
| Hook rows X hooks per row | 12 X 8-9 | 12 X 8-9 |
| Hooks 1-8/9 length (μm) | 56, 56, 50, 44, 41, 30, 24, 18 | 38-55, 35-55, 37-45, 35-52, 30-45, 27-40, 18-37, 18-30, 12-22 |
| Hook root | Asymmetrical with anterior process | Asymmetrical with anterior process |
| Receptacle L X W (μm) | Double-walled: 550-900 X 200 | Double walled: 395-676 X 125-239 |
| Cephalic ganglion | Near middle of receptacle | Near middle of receptacle |
| Lemnisci (μm) | About as long as receptacle | Equal, baculiform, longer than receptacle: 458-832 X 73-187 |
| Anterior testis L X W(μm) | Round, equatorial | 333-675 X 166-425 |
| Posterior testis L X W (μm) | Round, equatorial | 372-575 X 250-500 |
| Cement glands | Pyriform, 4 | Pyriform, 4: 156-675 X 104-325 |
| Uterus & uterine bell L (mm) | 1.4 | ---- |
| Gonopore | Ventro-subterminal in both sexes | Ventro-subterminal in both sexes |
| Eggs L X W (μm) | 40 X 16 | 25-38 X 9-15, with polar prolongation |
Table 2: Redescription and comparative morphometrics of Micracanthorhynchina motomurai from Zacco spp. in Korea.
Comparisons were made with the specimens described by Harada [7, 9] in the same Table 2. In his original description, Harada [7] did not designate type material and none was found to examine. We therefore, designate a lectotype male, an allolectoptype female, and paralectotypes with our redescription.
• Redescription of Micracanthorhynchina motomurai (Harada, 1935) Based on 8 males and 4 females. Qualitative and quantitative descriptions of males and females are outlined in Table 2 covering the complete characteristics of M. motomurai (Harada, 1935) in a tabulated format. Our figures 9-16 illustrate the fusiform shape of males and females (Figs. 9, 10), the club-shaped proboscis (Figs. 12, 14), the double- walled receptacle and the lemnisci of corresponding size (Figs. 11, 13), dorsal and ventral trunk spines (Fig. 12), male reproductive system with sub-ventral gonopore (Fig. 9, 15), and the sub-ventral gonopore of females (Fig. 16).
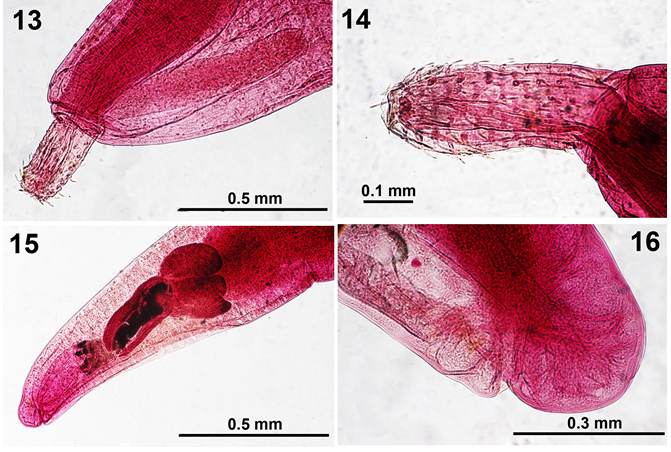
Figures 13-16: Light microscope images of female specimens of Micracanthorhynchina motomurai from Zacco temmincki & Zacco platypus in Korea. 13. A perspective of the shape and size of a lemniscus compared to that of the receptacle and a partially retracted proboscis. 14. A high magnification of a female proboscis. 15. The posterior part of a male reproductive system showing the sub-ventral gonopore 16. The posterior portion of a female reproductive system showing the sub-ventral gonopore.
- Specimens: Deposited at the Nebraska State Museum’s Harold W. Manter Laboratory (HWML) Parasitology Collection no. 217504 (lectotype), no. 217505 (allolectotype) and no. 217506 (paralectotypes; 3 slides).
- Remarks: Our specimens demonstrated that the trunk spines are more numerous and relatively larger in females than in males and larger ventrally than dorsally. The lemnisci were equal and measured on the average longer than the proboscis receptacle. The posterior testis was shown to be shorter and wider than the anterior testis. The cement glands were measured for the first time. The eggs showed polar prolongation of the fertilization membrane. Table 2 documenting the above observations, among others, that are in agreement with Harada’s [7] incomplete account, constitutes a redescription of M. motomurai.
Acknowledgements
This project was partially supported by an ongoing institutional grant from the Parasitology Center, Inc., Scottsdale, Arizona, USA. We are immensely grateful to the collaborators who contributed the study materials that made the execution of this work possible. Collaborators simply shared research material as part of joint research understandings. We also gratefully recognize the help of Dr. Gabor Racz., Collection Manager, Harold W. Manter Laboratory of Parasitology, University of Nebraska-Lincoln for kindly accessing and cataloging our specimens.
References
-
Amin OM, Rubtsova NY (2023a) New Annotated Records of Helminth Parasites, Mostly from North America, with Light Microscopy. I. Trematoda (Digenea), Cestoda, Nematoda. Int J Zoo Animal Biol 6(5): 000507.
-
Amin OM, Rubtsova NY (2023b) New Annotated Records of Helminth Parasites with Light Microscopy II. Acanthocephala (Polymorphida). Int J Zoo Animal Biol 6(6): 000530.
-
Amin OM, Rubtsova NY (2023c) New Annotated Records of Helminth Parasites with Light Microscopy. III. Acanthocephala (Archiacanthocephala and Eoacanthocephala). Int J Zoo Animal Biol 6(5): 000507.
-
Cable RM, Lideroth J (1963) Taxonomy of some Acanthocephala from marine fishes concerning species from Curaçao, N. A. and Jamaica, W. I. J Parasitol 49 (5): 706-716.
-
Amin OM, Chaudhary A, Singh HS (2023) Redescription of _Illiosentis cetratus_ Van Cleave, 1945 (Acanthocephala: Illiosentidae) from _Menticirrhus undulatus_ (Girard) in California, with notes on _Illiosentis furcatus_ from Peru. Folia Parasitol 70: 018.
-
Van Cleave HJ, Lincicome DR (1940) A reconsideration of the acanthocephalan family Rhadinorhynchidae. J Parasitol 26 (1): 75-81.
-
Harada I (1935) Zur Acanthocephalen fauna von Japan. Memoirs of the Faculty of Science and Agriculture, Taihoku Imperial University of Formosa. Japan 14 (2): 7-23.
-
Ward HL (1951) The species of Acanthocephala described since 1933. I. J Tenn Acad Sci 26 (4): 282-381.
-
Harada I (1938) Acanthocephalen aus Formosa (1.). Dr. A. OKA Jub., Annotationes Zoologicae Japonenses 17(3/4): 419-427.
-
Petrochenko VI (1956) Acanthocephala of Domestic and Wild Animals, I, Moscow, Acad Sci USSR pp: 465.
-
Golvan Y (1969) Systématique des Acanthocephales (Acanthocéphala Rudolphi, 1801). Mémoires du Muséum Natl d’histoire Nat 47: 1-373.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework