Controlling Proteocephalus ambloplites (Cestoda) Infections and Changing Fish Parasitic Fauna Following Post-Rotenone Rehabilitation of Lakes, With Light Microscopy
Efforts to establish desirable fish populations by rotenone treatment of lakes dominated by winter-kill resistant species, e.g., bullhead, and restocking have been attempted by the Wisconsin Department of Natural Resources, as well as by other agencies throughout the United States since the mid-1930s. Such efforts were performed in Little Elkhart Lake, eastern Wisconsin which was treated with rotenone (1.5 ppm) in 1981, to kill its rough fish and restock with desirable fish populations. Pre- and posttreatment samples of fish were examined for parasites in September 1981 and 1988, respectively. This is a revision and update of work originally published in 1993 with the addition of light microscope images of recovered parasites for the first time. Relatively heavy infections in largemouth bass, Micropterus salmoides, (Lacépède) and sunfish in 1981 with Proteocephalus ambloplites (Leidy, 1887) (Cestoda) disappeared after the rotenone treatment. Infections with Neoechinorhynchus cylindratus Van Cleave, 1913 decreased but drastically increased with Leptorhynchoides thecatus (Linton, 1891) Kostylew, 1924 with changes in their fish and invertebrate host populations following rotenone use. Metacercariae of Posthodiplostomum minimum (MacCallum, 1921) (Trematoda) in sunfish and Hysterothylacium brachyurum Ward and Magath, 1917 (Nematoda) in largemouth bass were also present before and after the chemical treatment, respectively. The observed loss of P. ambloplites after 1981 points to a possible method for its control. The unusual presence of the leech Myzobdella lugubris in the anterior intestine of a yellow perch, Perca flavescens (Mitchill) was documented.
Omar M Amin* and Nataliya Y Rubtsova
Keywords: Proteocephalus ambloplites; Parasites; Lakes
Introduction
Parasites are ubiquitous, primarily surviving in a dynamic equilibrium with their host(s) and they are often overlooked in fish health assessments. Changes in the environment, both anthropogenic and environmental, can alter the parasite or host equilibrium and cause disease or mortality in fish. In large lakes, seasonal periodicity in development and maturation of parasites is usually not observed. Our study of Echinorhynchus salmonis Müller in Lake Michigan [1] indicated that recruitment and maturation were associated with colder and warmer temperatures, respectively, and continued throughout the year. The same observation was also made for other parasites in other lakes elsewhere. Two aspects were included in our present study: The first is related to some physical and chemical properties of a small lake in Wisconsin. The second is the diagnosis of the parasites and their intensity pre-and post-chemical treatment.
In November 1981, Little Elkhart Lake, a small lake in Wisconsin, was treated with rotenone to kill its rough fish and restock with desirable fish populations. Before the chemical treatment in 1981, the fish population was dominated by black bullhead, Ictalurus (=Ameiurus) melas Rafinesque, and a small number of northern pike, Esox Lucius Linn.; walleye, Stizostedion (=Sander) vitreum (Mitchill); largemouth bass, Micropterus salmoides (Lacépède); bluegill, Lepomis macrochirus Rafinesque; pumpkinseed, Lepomis gibbosus (Linn.); black crappie, Pomoxis nigromaculatus (Lesueur); yellow perch, Perca flavescens (Mitchill); white sucker, Catostomus commersoni (Lacépède); golden shiner, Notemigonus crysoleucas (Mitchill); and yellow bullhead, lctalurus (=Ameriurus) natalis (Lesueur) [2, 3]. A typical complex of lake invertebrates included abundant copepods cladocerans and rare daphnias [3, 4]. Shore terrestrial vertebrates that may be involved in the life cycle of helminths infesting fish as larvae, included many bird species (mallard, blue-wing, teal, wood duck, etc.) and mammals (mink, weasel, muskrat, raccoon, red squirrel, eastern grey squirrel, etc.) [2]. Little Elkhart Lake has a history of stunted sunfish and intermittent good largemouth bass populations. It is characterized by infrequent winter kill (1952, 1959), chemical treatment and restocking with northern pike, largemouth bass, walleye, yellow perch, and golden shiner in 1961 [5, 6], and the subsequent accidental introduction of black bullhead, pumpkinseed, and largemouth bass by 1971. The black bullhead population increased as another winter kill in 1975 [7] caused weeds and undesirable fish species to become overabundant.
Our objective was to sample the fish from Little Elkhart Lake twice for parasites, once to provide a baseline of the helminth community before the lake was treated with rotenone and another time after treatment to evaluate the impact of chemical treatment.
Materials and Methods
Study Area
Little Elkhart Lake is a 23-ha glacial meso-eutrophic lake that is 3 and 8 meters in average and maximum depth and 1.3 years in average water residence time, in Sheboygan County, eastern Wisconsin (43° 49’ 59.9808’’ N, 88° 1’ 4.3536’’ W). The lake is an extension of the hard groundwater table with moderate nutrient levels, e.g., total phosphorus, nitrogen, and alkalinity averaged 0.04, 1.0, and 124 mg/1, respectively, at various water depths [5, 6]. Dominant species of rooted aquatic vegetation included Myriophyllum spicatum, Najas flexilis, Potamogeton illinoensis, and Potamogeton amplifolious. Algae were present in low densities [2, 7].
Methods
The lake was electroshocked to sample pre-treatment fish on September 22 and 23, 1981. On November 17, 1981, 220 gallons of rotenone were applied at a concentration of 1.5 ppm. Except for bullhead, total kill of all other fish species, including largemouth bass and sunfish, was noted. The crustacean and other invertebrate communities, however, remained practically intact (Nelson 1985a and pers. comm.) [3]. In May and June, 1982, the lake was stocked with 10,000 fingerling largemouth bass from Crystal and Gerber lakes, and 54,310 fingerling hybrid sunfish (bluegill x green sunfish) and bluegill from Beechwood Lake, as well as with 118 big largemouth bass up to 37 cm long. Subsequent electroshocking efforts in 1983 and 1984 demonstrated good survival of largemouth bass, sunfish hybrid, bluegill, as well as black bullhead [4]. On September 1, 1988, a post-treatment sample of fish was similarly taken from the lake. Both pre- and post-treatment fish samples were promptly examined for parasites after transfer to the lab on ice. Parasites were systematically recovered and routinely processed and whole- mounted for microscopical examination.
Processing for Microscopy
Specimens were placed in water overnight or until fully extended then fixed in 70% ethanol. Acanthocephalans were punctured with a fine needle and often subsequently stained in Mayer’s acid carmine, de-stained in 4% hydrochloric acid in 70% ethanol, dehydrated in ascending concentrations of ethanol (24 hr each), and cleared in 100% xylene then in 50% Canada balsam and 50% xylene (24 hr each). Whole worms were then mounted in Canada balsam.
Optical Microscopy
Images were acquired using a Zeiss Axioskop Transmitted Nomarski DIC Phase Contrast Microscope Trinocular (Munich, Germany) and a Canon T3i EOS 600D DSLR Camera (Melville, New York). Measurements are in micrometers unless otherwise noted; the range is followed by the mean values between parentheses when appropriate. Width measurements represent maximum width.
Results and Discussion
Three fish species were collected and examined during each of the 1981 and 1988 surveys: largemouth bass: 40 fish, 16-41 (mean 26) cm long in 1981 and 42 relatively smaller fish, 17-28 (24) cm long in 1988, bluegill: 9, 13-18 (15) cm long in 1981 and 12, 10-20 (15) cm long in 1988, and 4 pumpkin-seed in 1981 (Table 1). Largemouth bass and bluegill, along with all other species, except black bullhead, were totally eliminated from the lake as a result of the 1981 treatment. The 1988 samples were from new fish introductions mostly during 1982. Four pumpkinseed, 12 black crappie, and 13 yellow perch were also examined in 1981, and 7 black bull-head were examined in 1988.
| Fish species | Parasite species | Site of infection | Pre-treatment (1981) | Post-treatment (1988) | ||
|---|---|---|---|---|---|---|
| Fish infected/ examined (%) | No. parasites (mean) | Fish infected/ examined (%) | No. parasites (mean) | |||
| Micropterus salmoides | Proteocephalus ambloplites | Intestine | 34/40 (85%) | 138 (3.45) | 0/42 (0) | 0 (0) |
| Body cavity | 32/40 (80) | 357 (8.93) | 0/42 (0) | 0/42 (0) | ||
| Neoechinorhynchus cylindratus | Intestine | 34/40 (85) | 382 (9.55) | 29/42 (69) | 79 (1.88) | |
| Body cavity | 2/40 (5) | 2 (0.05) | 0/42 (0) | 0 (0) | ||
| Leptorhynchoides thecatus | Intestine | 10/40 (25) | 85 (2.12) | 41/42 (98) | 2,698 (64.24) | |
| Body cavity | 1/40 (2.5) | 15 (0.37) | 13/42 (31) | 39 (0.93) | ||
| Hysterothylacium brachyurum | Intestine | 0/40 (0) | 0 (0) | 7/42 | 5 (0.14) | |
| Body cavity | 0/40 (0) | 0 (0) | 6/42 | 6 (0.14) | ||
| Lepomis macrochirus | Proteocephalus ambloplites | Intestine | 0/9 (0) | 0 (0) | 0/12 (0) | 0 (0) |
| Body cavity | 4/9 (44) | 14 (1.56) | 0/12 (0) | 0 (0) | ||
| Neoechinorhynchus cylindratus | Intestine | 0/9 (0) | 0 (0) | 1/12 (8) | 1 (0.08) | |
| Body cavity | 0/9 (0) | 0 (0) | 0/12 (0) | 0 (0) | ||
| Leptorhynchoides thecatus | Intestine | 0/9 (0) | 0 (0) | 3/12 (25) | 23 (1.92) | |
| Body cavity | 0/9 (0) | 0 (0) | 6/12 (50) | 28 (2.33) | ||
| Posthodiplostomum minimum | Body cavity | 1/9 (9) | 0 (0) | 0/12 (0) | 0 (0) | |
| Lepomis gibbosus | Posthodiplostomum minimum | Body cavity | 3/4 (75) | 32 (8.0) | 0 (0) | 0 (0) |
| Proteocepohalus ambloplites | Body cavity | 2/4 (50) | 8 (2.0) | 0 (0) | 0 (0) | |
| Perca flavescens | Myzobdella lugubris | Intestine | 1/1 | 1 (100) | 0 (0) | 0 (0) |
Table 1: Pre- and post-treatment surveys of fish parasites in Little Elkhart Lake, Wisconsin.
The following presentation addresses 6 species of parasites: 1 cestode, 2 acanthocephalans, 1 nematode, 1 trematode metacercariae, and 1 leech species listed in Table 1 with the prevalence data in the pre- and post-treatment surveys.
Proteocephalus ambloplites (Leidy, 1887) (Proteocephalidae: Cestoda) (Figs. 1-18)
In the 1981 pre-treatment study, the prevalence and intensity of Proteocephalus ambloplites (Leidy) (Cestoda) infections in largemouth bass were moderate to heavy in intestinal and body cavity (gut surface, liver, spleen) locations (Table 1); only 27 mature gravid adults were localized in the intestine. The distribution of P. ambloplites also extended into the body cavity of other fish species, e.g., 14 worms in 4 of 9 bluegill (Table 1), 1 in 1 of 12 black crappie, 6 in 5 of 13 yellow perch, and 4 in 2 of 4 pumpkinseed. In the 1988 post- treatment study, P. ambloplites was absent from all sites in all 3 fish species examined then, including 42 largemouth bass, its major host. The complete disappearance of P. ambloplites from bass and sunfish is perhaps the most dramatic outcome of the 1981 chemical treatment. This clearly corresponded with the mortality of the definitive host, largemouth bass (where adult worms reproduce), and the principal intermediate hosts, e.g., bluegill and other sunfish species, and probably resulted from it, following the application of rotenone. The assumption that the loss of P. ambloplites was related to changes in the fish and not the crustacean (cladoceran, copepod, or amphipod) host populations is based on the following. The increase in L. thecatus infections after the treatment must have been related to the survival of its amphipod intermediate hosts including Hyalella sp., e.g., H. knickerbockeri which is also an intermediate host for P. ambloplites. The mortality of the definitive and paratenic hosts of L. thecatus, particularly largemouth bass and sunfish in 1981, indicates that L. thecatus could have only survived the chemical treatment in the crustacean host. The survival of N. cylindratus was also possible because of the survival of its ostracod intermediate host, possibly Cypria sp. declined to zero after 72 hours [8]. These data support findings about the mortality of fishes and survival of Crustacea after the 1981 treatment of Little Elkhart Lake with rotenone. The complete eradication of the bass tapeworm, P. ambloplites, by eliminating its fish hosts in a closed system like Little Elkhart Lake under the above conditions points to a practical method for the control of this injurious worm. This goal can be accomplished at the same time the quality of the fish fauna is being upgraded.
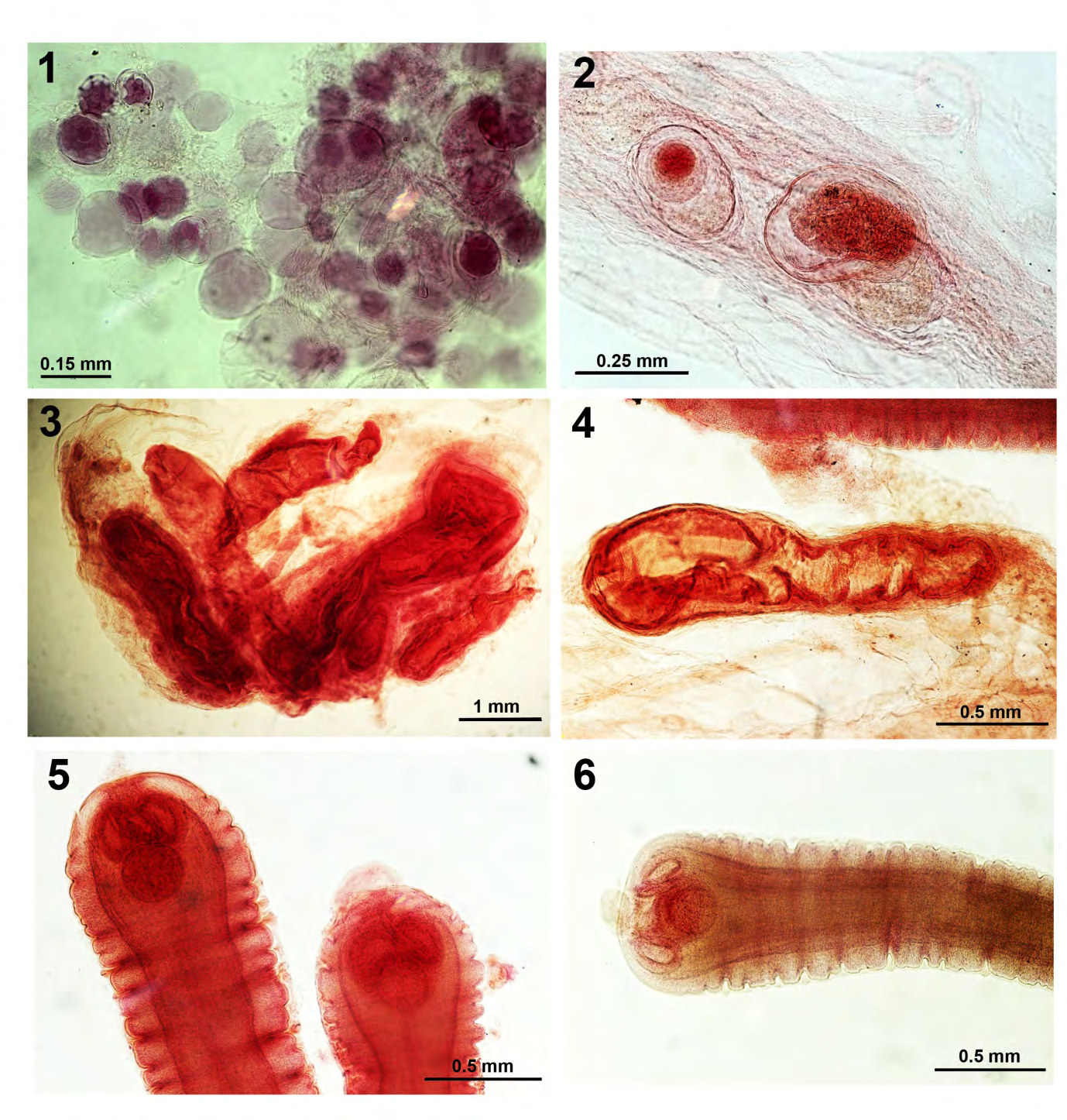
Figures 1-6: Larval stages of Proteocephalus ambloplitis encysted in the body cavity, especially the peritoneal membranes, of Micropterus salmolides from Little Elkhart Lake, Wisconsin. 1. A cluster of cystic forms. 2. Higher magnification of two cysts. 3, 4. Elongated forms are trapped in the peritoneum. 5, 6. More developed small plerocercoids showing some strobilar segmentation, underdeveloped scolex, and a posterior accessory fifth sucker.
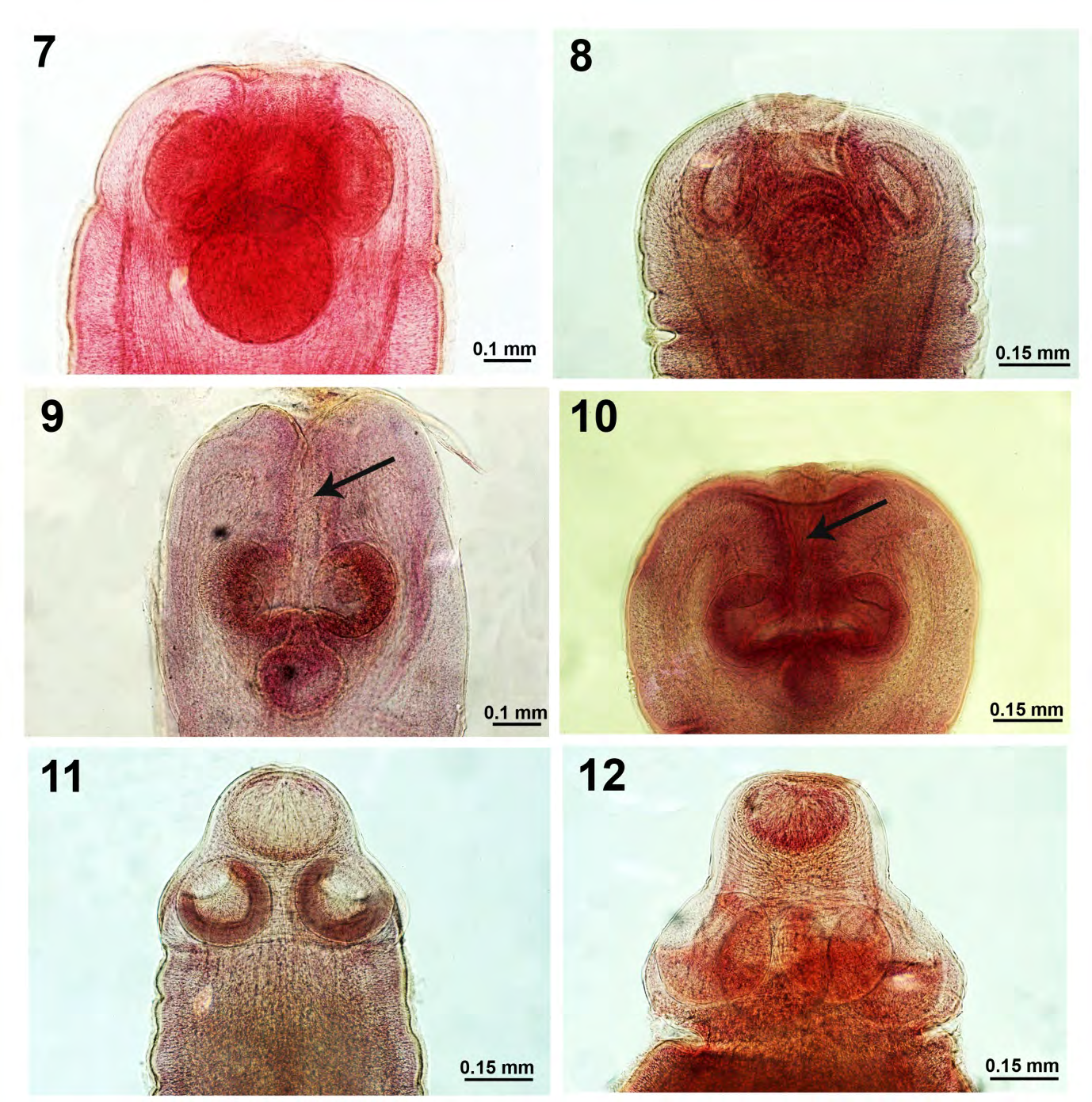
Figures 7-12: Plerocercoids of Proteocephalus ambloplitis in the body cavity of Micropterus salmolides from Little Elkhart Lake, Wisconsin. 7-10. Variations in the organization of the 4 major suckers relative to the posterior accessory sucker in 4 worms. Note the connectivity between the accessory suckers to the 4 anterior suckers and the anterior apical part of the scolex in Figs. 9 & 10. The open anterior furrow (Figs. 9 & 10 arrows) allows for the passage of secretory exudate to be eliminated apically from the posterior accessory sucker. 11, 12. Variations in the scolex of more advanced plerocercoids show the displacement of the accessory suckers anteriorly.
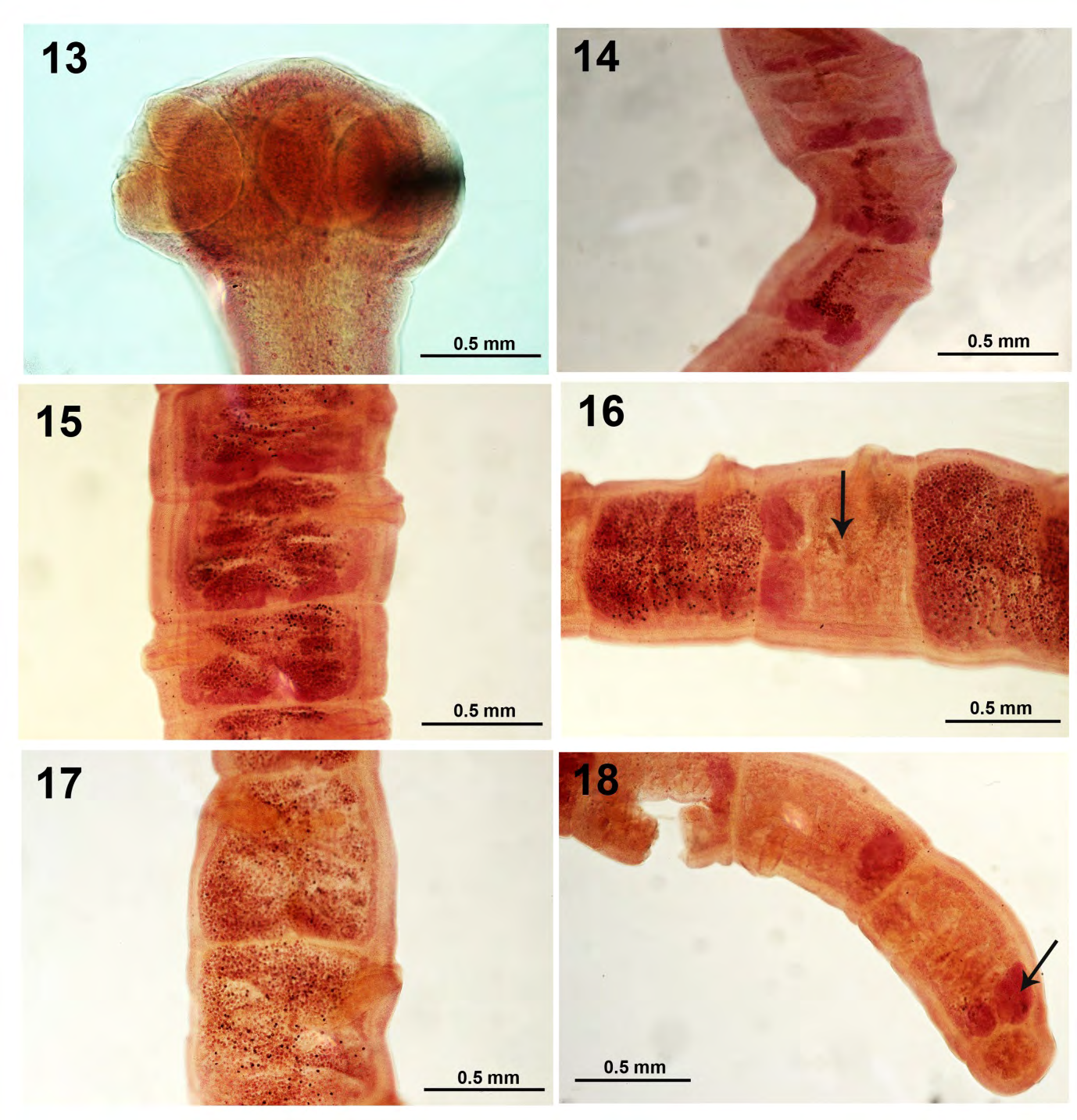
Figures 13-18: Adult Proteocephalus ambloplitis in the intestinal tract of Micropterus salmolides from Little Elkhart Lake, Wisconsin. 13. A fully developed mature adult scolex. 14-18. Progressive development from anterior mature proglottids to posterior gravid proglottids in different worms. Note the irregularly alternating reproductive orifice, the occasional alternation of mature a proglottid within a sequence of gravid proglottids (Fig. 16, arrow), and the persistence of the ovaries till the terminal proglottid (Fig. 18, arrow).
The morphology of the 1981 pretreatment specimens varied depending on the site of infection. The body cavity forms were smaller plerocercoids from <1.00 to 7.00 mm long. The smallest were underdeveloped cyst forms with undifferentiated structures deeply embedded in intestinal mesenteries (Figs. 1-4). Slightly larger forms began to develop a scolex with a 5th accessory ventral sucker (Figs. 5-10) that became displaced apically (Figs. 11,12) which appears to have secretory activity. Short slightly segmented strobilae develop afterwards. The work of Amin and Boraini [9] shed considerable light on the comparative morphology of P. ambloplitis plerocercoids in centrarchids from other Wisconsin lakes with 10-line drawings of plerocercoid metamorphosis. Amin and Elnaggar [10] also demonstrated tissue sections of P. ambloplites plerocercoids in double- walled cysts in the gut wall of various centrarchid fish species from other Wisconsin lakes. Amin [11] studied the ecology and pathology of parenteric P. amploplites in 17 species of fish intermediate hosts from two southeastern Wisconsin eutrophic lakes and Clothier and Heckmann [12] made a histopathological study of P. ambloplites plerocercoids in hepatic tissue of smallmouth bass.
The intestinal forms of P. ambloplites in the pretreatment survey were in two forms: developing plerocercoids with 5 suckers on the scolex representing recent recruitments and developing mature adults reaching 70 mm in length in about equal proportions. The adults were gravid with mature scolex and fully developed 4 suckers (Fig. 13). Developmental stages in the maturation of strobila show mature to fully gravid proglottids in Figs. 14-18. Amin and Cowen [13] discussed the ecology of P. ambloplites in bass in two other nearby southeastern Wisconsin lakes. Amin and Cowen [13] demonstrated the importance of critical temperatures and host size in the parenteric recruitment of P. ambloplitis plerocercoids in the spring.
Specimens. Adults and plerocercoids in the Harold W. Manter Lab., Parasitology collections at Lincoln, Nebraska nos. 217575 & 217576.
Neoechinorhynchus cylindratus (Van Cleave, 1913) Van Cleave, 1919 (Neoechinorhynchidae: Acanthocephala) (Figs. 19-24)
The prevalence and intensity of N. cylindratus (Acanthocephala) was relatively high in largemouth bass before the chemical treatment in 1981 but decreased considerably by 1988 (Table 1). In the 1981 pretreatment survey, the population was reproductively active including mostly gravid females. The mortality of sunfish in 1981 was also considered responsible for the dramatic decline in N. cylindratus population. The major intermediate host of this acanthocephalan species is bluegill [14]; other sunfish species are important paratenic (transport) hosts, e.g., pumpkinseed among others [15]. The fact that one of the paratenic hosts of N. cylindratus is black bullhead, which survived the 1981 chemical treatment, might have helped that acanthocephalan’s marginal survival (Table 1).
Amin studied the host and seasonal distribution of 5 species of Neoechinorhynchus Hamann, 1892 from two nearby lakes in SE Wisconsin between 1974 and 1984. Fourteen species of fish in 7 families were infected with N. cylindratus. We collected 4,158 specimens (18% of 807 examined fish were infected with a mean of 5.15) of N. cylindratus in the land-locked Silver Lake (188 ha) in Kenosha County compared to only 168 specimens (3% of 778 examined fish were infected with a mean of 2.22) in Tichigan Lake (458 ha), a tributary of the Mississippi River in Racine County which is an impoundment on the Fox River. Both Silver Lake and Little Elkhart Lake are land- locked. Amin [16] observed that of the 14 species of fish infected with N. cylindratus in Silver and Tichigan lakes, M. salmoides was the principal definitive host. This situation may be comparable to the case in Little Elkhart Lake. The size, form, and degree of development of body cavity forms and the more mature intestinal forms of N. cylindratus were quite variable. Major morphological features of adult forms from Little Elkhart Lake include variability of lemnisci length (Figs. 19-21), proboscis and hooks (Fig. 22), and female reproductive system showing the swollen sub- ventral gonopore complex, the sub-vaginal muscle, and the undulating uterine bell wall (Figs. 23, 24).
Specimens. In the Harold W. Manter Lab., Parasitology collections at Lincoln, Nebraska no. 217577 and mixed with Leptorhynchoides thecatus no. 217578.
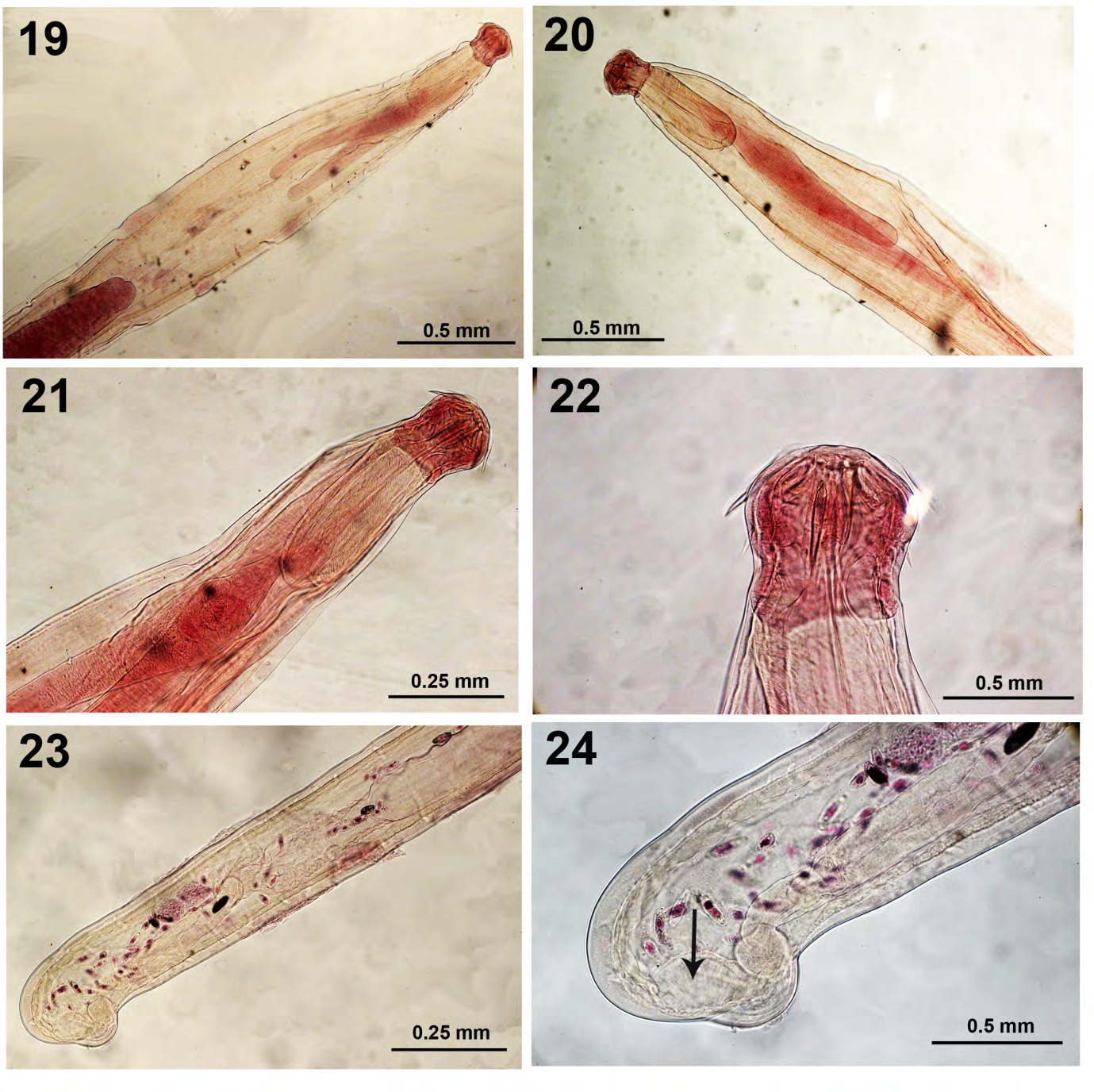
Figures 19-24: Specimens of Neoechinorhynchus cylindratus from the intestine of Micropterus salmoides in Little Elkhart Lake, Wisconsin. 19, 20. The anterior parts of two male specimens showing the relative sizes of the proboscis, receptacle, and lemnisci. Note the variability in the size of lemnisci being equal (Fig. 19) and unequal (Fig. 20). 21. A higher magnification of the anterior part of a third specimen. 22. Enlarged proboscis showing a greater content of chitinous material (dark color) compared to the trunk. 23. The posterior part of a female specimen showing the reproductive system complete with the vagina, thick uterus, selective apparatus, and undulating uterine bell. 24. A higher magnification of the posterior-most part of a female emphasizing the thick uterus, vagina with a single sphincter, gonopore just anterior to the swollen sub-ventral knob and the sub-vaginal muscle (arrow).
Leptorhynchoides thecatus (Linton, 1891) Kostylew, 1924 (Rhadinorhynchidae: Acanthcephala) (Figs. 25- 30)
The trend in L. thecatus infections was opposite that in N. cylindratus; its mean intensity of infection increased dramatically from 2.12 to 64.24 per examined fish in gut locations and from 0.37 to 0.93 in the body cavity of largemouth bass in the pretreatment vs. post-treatment, respectively (Table 1). Bluegill were not infected with either acanthocephalan species in 1981 but were considerably more frequently and heavily infected with L. thecatus than with N. cylindratus in the post-treatment samples of 1988 (Table 1) indicating new infections acquired with stocking bass. In addition, 176 specimens of L. thecatus were also recovered from the gut of 5 of 7 black bullhead (48 worms) and the body cavity of 2 other bullheads (128) in 1988 but not observed in 1981.
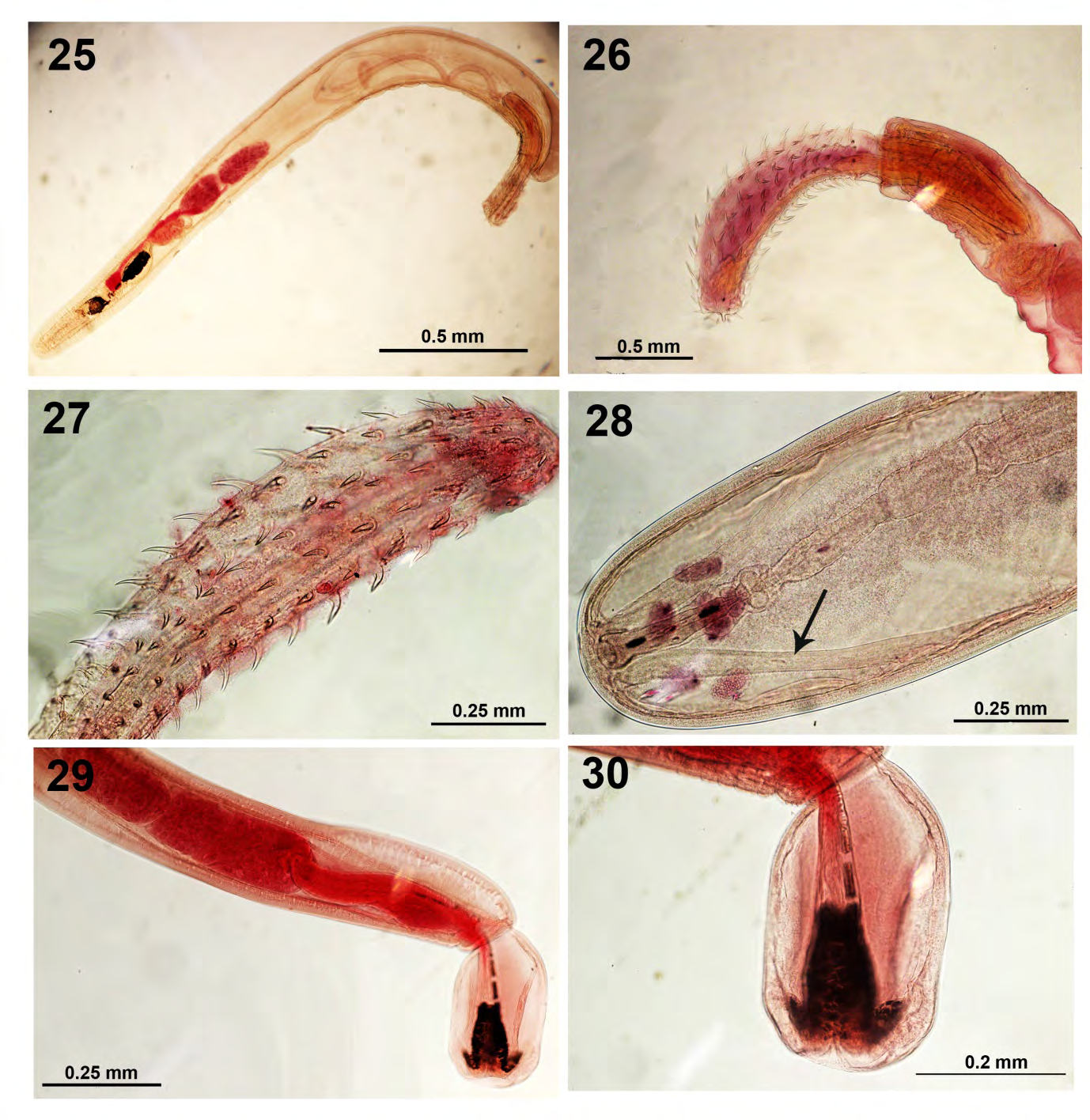
Figures 25-30: Specimens of Leptorhynchoides thecatus from the intestine of Micropterus salmoides in Little Elkhart Lake, Wisconsin. 25. A whole male specimen showing the relative size and proportion of the posteriorly positioned reproductive system and the long cylindroid lemnisci. 26. The size of the proboscis of a male specimen is considerably larger than the size of the receptacle. 27. A high magnification of another proboscis showing more detail of the hooks and their distribution. 28. The female reproductive system and two prominent para-vaginal ligaments (arrow) in one specimen. 29. The male reproductive system in one specimen. 30. A high magnification of the bursa of the specimen in Fig. 29.
In a related study from two nearby SE Wisconsin lakes; Silver and Tichigan lakes, L. thecatus was recovered from the ceca and other intestinal sites of 116 M. salmoides and 1,177 other fishes of 11 species in 6 families between 1976 and 1978. Thirty eight of 72 examined bass (53%) were infected with 138 specimens of L. thecatus (mean 2.28) from the land locked Silver Lake, and 17 of 44 examined bass were infected with 50 specimens of L. thecatus (mean 1.14) from the Fox River connected Tichigan Lake [17]. These results compare well with those from the land-locked Little Elkhart Lake in the 1981 pre-treatment survey with a mean of 2.12/fish in intestinal sites (Table 1). The extreme spike in bass infections in the post treatment may be related to patterns of infections in the restocking sources as monitored in 1988 (Table 1).
Morphological features of specimens of L. thecatus from various populations are represented in figures 25-30 emphasizing the male reproductive system (Figs. 25, 29), the proboscis and hooks (Figs. 26, 27), the bursa (Figs. 29, 30) and the female reproductive system with its prominent ligaments (Fig. 28). The ecology of the species in lake fishes in Wisconsin was studied by Amin [18].
Specimens. In the Harold W. Manter Lab., Parasitology collections at Lincoln, Nebraska no. 217579 and mixed with Neoechinorhynchus cylindratus no. 217578.
Sex Ratio of Acanthocephalans
In 1981, the sex ratio of N. cylindratus in largemouth bass was 1 male: 2.31 females (91% had eggs) and 1:1.44 in L. thecatus (all females had eggs). In 1988, the sex ratio shifted to 1:0.61 and 1:0.63 in N. cylindratus and in L. thecatus, respectively. Gravid females of both acanthocephalan species were considerably less frequent in 1988 than in 1981 indicating significant changes in the reproductive state of the two acanthocephalan species suggesting changes influencing the productivity levels of these two acanthocephalan populations indicating less populations’ fertility.
Hysterocelacium brachyurum (Ward and Magath, 1917) (Raphidascarididae: Nematoda (Figs. 31-34)
This nematode was absent in 1981 but was apparently introduced in the lake with the introduction of largemouth bass in stocking sources in subsequent years. Third-stage larvae of a species of Hysterothylacium were encysted in large numbers in the body cavity of largemouth bass and channel catfish from Lake Mohave, an impoundment of the Colorado River in Arizona [18]. These are two well-known host species of H. brachyurum in the Mississippi River basin. Worms were tightly coiled in a single plane (flat coil) within a tough hyaline cyst wall. Introduction of these nematodes into Arizona was likely coincident with Mississippi basin fishes. The anadromous striped bass, native to the Atlantic Coast and Gulf of Mexico, also could have transported this worm. This was the first published report of Hysterothylacium in Arizona. These nematodes appear to be widespread in nearby states [18] and may be of potential public health and veterinary health importance because some Hysterothylacium species are capable of penetrating the alimentary tract of mammals [19]. Hysterothylacium has apparently spilled over to some native and endangered fishes in Arizona and observed worms were large and markedly coiled but not enclosed in any membranes. Impact of infection on small, susceptible native fishes that may be threatened or endangered cannot be overlooked in stocking and management decisions [18]. The anterior end (Figs. 31, 32) and the posterior end (Figs. 33, 34) of specimens from Little Elkhart Lake are shown to feature the apical papilla (Fig. 32) and the conical tail (Fig. 34) (arrows) characteristic of that species.
Specimens. In the Harold W. Manter Lab., Parasitology collections at Lincoln, Nebraska no. 217580.
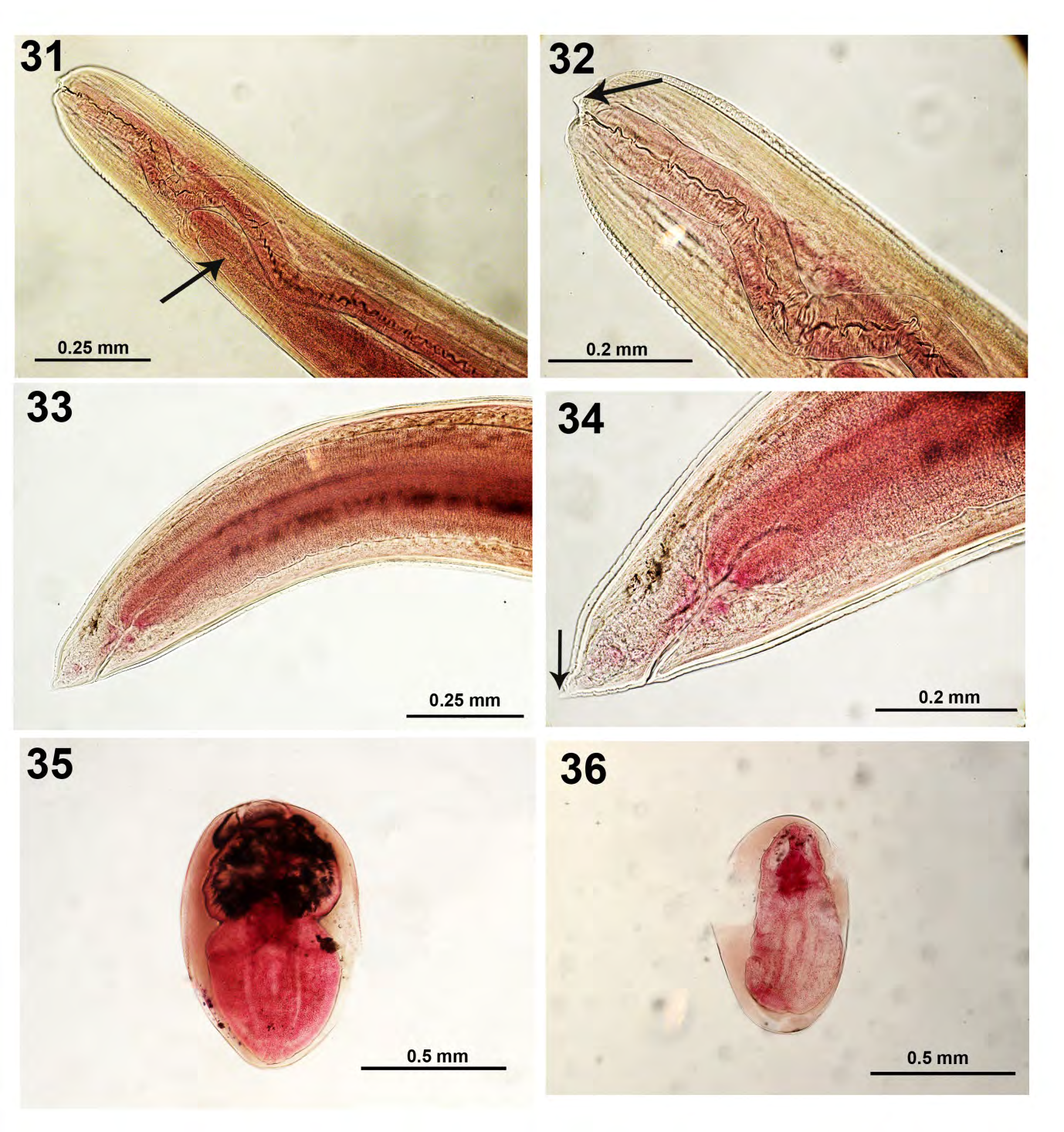
Figure 31-36: Four specimens of the nematode Hysterocelacium brachyurum from the intestine (Figs. 31-34) and 2 metacercariae of Posthodiplostomum minimum (Figs. 35, 36) in body cavity locations of Micropterus salmoides in Little Elkhart Lake, Wisconsin. 31, 32. Anterior body location of one specimen at two different magnifications showing the esophagus and intestinal caecum (Fig. 31, arrow) and an apical lateral papilla (Fig. 32, arrow) of H. brachyurum. 33, 34. Posterior part of the same specimen at two different magnifications showing the intestine and anus (Fig. 33) and the conical tail (Fig. 34, arrow). 35, 36. Two metacercariae of P. minimum from the body cavity.
Posthodiplostomum minimum (MacCallum, 1921) metacarcariae (Diplostomidae: Trematoda) (Figs. 35,36)
These metacercariae (Figs. 35, 36) were scarcely found in body cavity locations of L. machrocirus (1/9) but more commonly (3/4) in Lepomis gibbosus only during the pre- treatment survey of 1981 (Table 1). The loss of the light P. minimum infections after the 1981 treatment might have been affected by the mortality of its centrarchid fish intermediate hosts. The morphology of these metacercariae was comparable to that of metacercariae from this and other fish species in other Wisconsin waters that were, however, sampled in tissue sections [10]. Palmieri [20] studied host- parasite relationships and intraspecific variations in P. minimum. Specimens. In the Harold W. Manter Lab., Parasitology collections at Lincoln, Nebraska no. 217581.
Myzobdella lugubris Leidy, 1851 (Piscicolidae: Hirudinea) (Figs. 37-39)
Only one specimen of this leech species (Figs. 37- 39) was found in the anterior intestine of a yellow perch, Perca flavescens (Mitchill), in the 1981 survey. A total of 453 leeches of the same species were recovered from fish from two nearby lakes in Kenosha and Racine counties, Wisconsin between 1976 and 1978 [21]. These fish species did not include yellow perch but included 9 centrarchids and 4 species in 3 other families. The site of infection is unusual, but M. lugubris which is commonly found on fins, is known to also infest channel catfish most heavily around the mouth [21] and occasionally in the mouth of other fish [22]. The reproductive system of that one specimen had the 5 usual pairs of testes (Fig. 38) but Amin [21] reported many variations of fewer numbers reaching 1 single testis per specimen. The site of missing testes was not consistent. Anatomical details of specimens attached to the pectoral fin of L. macrochirus collected from Silver Lake, SE Wisconsin were shown (Figs. 55-57) of Amin and Elnaggar [10]. Specimen. In the Harold W. Manter Lab., Parasitology collections at Lincoln, Nebraska no. 217582.
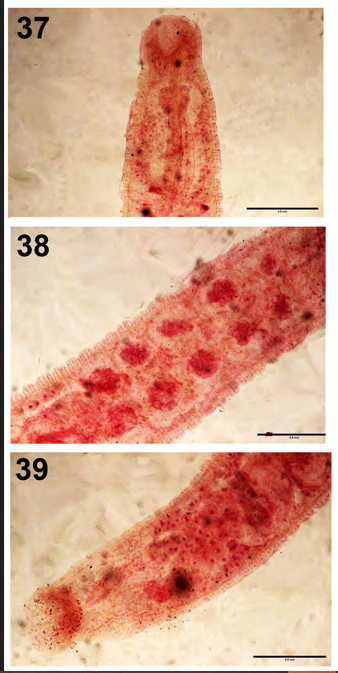
Figure 37-39: The piscicolid leach Myzobdella lugubris from the anterior intestine of a yellow perch, Perca flavescens, in Little Elkhart Lake, Wisconsin. 37. Anterior part showing oral sucker, pharynx, and esophagus. 38. Mid-section with the typical 5 pairs of testes. 39. Posterior part showing intestinal diverticula, rectum, and posterior sucker.
Chemical Rehabilitation
Establishing desirable fish populations by rotenone treatment of lakes dominated by winter-kill resistant species, e.g., bullhead, and restocking have been attempted by the Wisconsin Department of Natural Resources, as well as by other agencies throughout the United States since the mid-1930s [8]. The impact of such chemical treatments on invertebrate and fish populations and consequently on their parasite fauna is, however, not known. The case of the recent chemical rehabilitation of Little Elkhart Lake in eastern Wisconsin provided an opportunity to examine the effect of chemically induced host population changes on the composition and prevalence of parasite species (Table 1). Findings not only reflected answers to scientific curiosities, but also indicated a possible new approach to the control of some injurious fish parasites such as P. ambloplites.
Rotenone and Ecological Shifts
Parasite community succession stabilization was estimated to take at least 5 years from the initiation of major ecological shifts, e.g., impoundment [14]. In our study, 8 years elapsed between the pre- and the post-treatment studies. Brown and Ball [23], Sharma [24], and Wright [25] indicated that populations of microcrustaceans, e.g., Daphnia and Cyclops, were variably affected but not eliminated after treatment with rotenone. Major ecological shifts were shown not to deplete crustacean populations, e.g., copepods [14]. The half-life of rotenone in warm (24°C) and cold water (0°C) ponds in Wisconsin during September and March was 13.9 and 83.9 hr respectively [8]. Rotenone at a concentration of 1 ppm remained toxic to bluegills in wire cages for 7-18 days in ponds during an Alabama winter but caused no residual deleterious effects upon subsequent bluegill production [25]. These data support findings about the mortality of fishes and survival of Crustacea after the 1981 treatment of Little Elkhart Lake with rotenone.
Acknowledgments
This work could not have been completed without the contributions of two people. Colleen A. Dickey contributed to the post-treatment study while studying at the University of Wisconsin-Parkside. She had a BS in Biology in 1989 and a Master of Education from National-Louis University and used to teach at the Glenview District in Illinois. The other person is Alan R Spallato who contributed to the pre-treatment study while studying at the University of Wisconsin-Parkside where he received his BS in Biology in 1983. He worked at the Madison Hazleton Laboratories before he became an Environmental Chemist at the State Laboratory of Hygiene, Madison, Wisconsin, in 1989. The help and cooperation of James McNelly, Larry Claggett, and John Nelson, Wisconsin Department of Natural Resources was indispensable for the carrying out and the completion of the field work and for providing relevant resources.
References
-
Amin OM, Burrows JM (1977) Host and seasonal associations of _Echinorhynchus_ _salmonis_ (Acanthocephala: Echinorhynchidae) in Lake Michigan fishes. J Fish Res Board Can 34(3): 325-331.
-
Claggett L (1981) Little Elkhart Lake Fishery Rehabilitation. Environ Impact Assess Screen Worksheet Wis Dept Nat Resour.
-
Nelson JE (1985a) Progress report - F264 -Little Elkhart Lake - 1983 data. Wis Dept Nat Resour Intradept Memo. pp: 10.
-
Nelson JE (1985b) Progress report-F264-Little Elkhart Lake - 1984 data. Wis Dept Nat Resour Intradept Memo. pp: 14.
-
Schultz PT (1964) Little Elkhart Lake - 1963 survey. Wis Dept Nat Resour Intradept Memo.
-
Schultz PT (1965) Little Elkhart Lake, Sheboygan County - 1964 observations. Wis Dept Nat Resour Intradept Memo.
-
Amin OM, Dickey CA, Spallato AR (1993) The impact of chemical rehabilitation on the parasitic fauna of fish in a Wisconsin lake. Trans Wisc Acad Sc Arts Lett 81: 1-6.
-
Gilderhus PA, Allen JL, Dawson VK (1986) Persistence of rotenone in ponds at different temperatures. N Amer J Fisher Manage 6: 129-130.
-
Amin OM, Boraini MA (1992) Cestoda from lake fishes in Wisconsin: The morphological identity of _Proteocephalus_ _ambloplitis_. Trans Am Microsc Soc 111: 193-198.
-
Amin OM, Elnaggar A (2022) Diagnosis of helminth parasites in tissue sections of freshwater fish from Wisconsin, USA. Sci Parasitol 23(1-2): 1-19.
-
Amin OM (1990) Cestoda from lake fishes in Wisconsin. The ecology and pathology of _Proteocephalus ambloplitis_ plerocercoids in their fish intermediate hosts. J Helminthol Soc Wash 57(2): 113-119.
-
Clothier BD, Heckmann RA (2002) Plerocercoid infections of _Proteocephalus ambloplitis_ (Leidy, 1887) in hepatic tissue of smallmouth bass, _Micropterus dolomieu_ Lacépède, a histopathology study. Proc Parasitol 33: 1-25.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework