Bone Traces and Modifications from Feeding by Dermestes maculatus
That D. maculatus feeding can leave marks on bones is established in the literature - that D. maculatus routinely mark bones is not. Our results here, exposing fleshed bones to D. maculatus adults and immatures for almost two months after complete tissue removal, indicate that under natural conditions D. maculatus do not feed on bones. The key issue seems to be the ability of D. maculatus to seek new food sources versus being confined (and eventually starving) on bones. In a natural setting, once a food supply is exhausted, D. maculatus would seek new food sources. If this process is somehow interrupted, such as through placement of an infested body in a sealed container or experimentally over a long (months to a year) period, D. maculatus could feed on bone. Based on this conclusion, whether in forensics analysis or paleontological analysis, we think the working assumption should be that D. maculatus does not alter bone, with the exception (as we outlined) when beetles and bones are in an enclosure that prevents beetle emigration.
Introduction
Insects can use a wide array of biological materials (soft tissues, feces, bones) for feeding, reproduction, and shelter. When they feed on vertebrate tissue, bones may be modified by insect action, and various insect taxa are known to modify bone with their mandibles, including members of the families Dermestidae, Tenebrionidae, Tineidae, and Termitidae. Despite bone modification being a known behavioral trait of many of these insects, little work has been done to record the frequency of occurrence, species-specific modifications of bone, and concise, definitive criteria distinguishing insect modifications from other agents impacting bones.
Markings on bones can have many causes. In particular, weathering causes many types of bone modifications that can be confused with insect modifications [1]. Weathering itself is potentially valuable as evidence indicating the period of time represented in recent or fossil bone assemblages, including those on archeological sites. It also can be an important tool in censusing animal populations in modern ecosystems [2].
Because dermestid beetles are commonly associated with late-stage decomposition of animals, they often feature as entomological evidence in homicides with human decomposition as well as in archeological investigations. Consequently, potential dermestid modification of human bone is of anthropological and forensic importance. Unfortunately, whether or not common dermestid species can alter bone, how they alter bone, or under what circumstances they alter bone are all questions of continuing debate.
The feeding activities of insects, like those of vertebrate scavengers and predators, change the remains and may leave artifacts that can be difficult to assign to a cause. When dry tissues are removed by dermestid beetles, the cause of tissue removal is usually obvious: dermestids are one of the few agents that feed on dried tissue, and sites of dermestid feeding routinely have frass and cast larval skins.
Nevertheless, we find suggestions, and even false confirmations, on both sides of the argument that dermestid beetles can modify bone. One aspect of this debate, contrasts workers in forensic science with workers in paleontology. While forensic scientists seem to be unresolved on this issue, many papers in paleontology treat marks on fossil bones being from dermestids almost as dogma. Hypothesized trace makers most commonly include carrion insects such as dermestid, silphid, and histerid beetles, tineid moths, and a variety of neotropical termite species [1]. Tobien [3] first recognized Neogene mammal bones from Germany that contained distinctive ovoid chambers (2–7 mm in diameter), which he interpreted as dermestid beetle pupal chambers. Similar borings were described in Pleistocene mammal bones from South Africa by Kitching [4], who also associated them with dermestid beetles. Martin and West [5] described slightly smaller (2–4 mm) ovoid bone borings from the late Pliocene of Idaho and the middle-late Pleistocene of Kansas. They followed earlier workers and also attributed these traces to dermestid beetles. However, no direct evidence supporting these various claims was provided in these publications.
In the forensic literature, Schroeder, et al. [6] reported that D. maculatus larvae damaged the humerus and the acetabulum of a human skeleton recovered from indoor conditions [6], furthering the notion that dermestid use beetles bone for pupation chambers and leave marks on bones, despite having only observational data to support their claims. Roberts, et al. [1] conducted a study aimed at establishing modification criteria to bone by dermestids whilst measuring the influences of food availability, food type, and substrata in increasing/decreasing bone modification. Results of this study suggested that a wide variety of modification types were produced by dermestids, including oval-shaped borings into cortical bone and irregular excavations into trabecular (spongy) bone, however, preference was shown for marrow cavities of long bones.
Kirkland, et al. [7], when working with dermestid colonies, observed that dermestids often remove the periosteum from cortical bone, however, most destruction occurred on softer cancellous bone, particularly bird bones, but also on articular facets of mammal bones [8]. Experiments conducted by Hefti, et al. [9] found that once available food sources have been depleted the beetles began to destroy specific areas: particularly the iliac crest of the pelvis and vertebrae. Hefti, et al. [9] goes on to state that “the beetles attack bone when they are deprived of other food.”
Microdamage and Taphonomy of Bone Weathering
Weathering is defined as the process by which the original microscopic organic and inorganic components of a bone are separated from each other and destroyed by physical and chemical agents operating on the bone in situ, either on the surface or within the soil zone [2]. This can strongly affect paleoecologic interpretations concerning the faunal composition, relative abundances of taxa and age- structure of the preserved populations because it indicates that taphonomic biases inherent in an attritional bone assemblage must be taken into consideration [2]. Divided into several stages, bone weathering was referred to in our analysis and is categorized as follows [2]: Stage 0: Bone surface shows no sign of cracking or flaking due to weathering. Usually bone is still greasy, marrow cavities contain tissue, skin and muscle/ligament may cover part or all of the bone surface. Stage 1: Bone shows cracking, normally parallel to the fiber structure (e.g., longitudinal in long bones). Articular surfaces may show mosaic cracking of covering tissue as well as in the bone itself. Fat, skin and other tissue may or may not be present. Stage 2: Outermost concentric thin layers of bone show flaking, usually associated with cracks, in that the bone edges along the cracks tend to separate and flake first. Long thin flakes, with one or more sides still attached to the bone, are common in the initial part of Stage 2. Deeper and more extensive flaking follows, until most of the outermost bone is gone. Crack edges are usually angular in cross-section. Remnants of ligaments, cartilage, and skin may be present. Stage 3: Bone surface is characterized by patches of rough, homogeneously weathered compact bone, resulting in a fibrous texture. In these patches, all the external, concentrically layered bone has been removed. Gradually the patches extend to cover the entire bone surface. Weathering does not penetrate deeper than 1.0-1.5 mm at this stage, and bone fibers are still firmly attached to each other. Crack edges usually are rounded in cross-section. Tissue is rarely present at this stage. Stage 4: The bone surface is coarsely fibrous and rough in texture; large and small splinters occur and may be loose enough to fall away from the bone when it is moved. Weathering penetrates into inner cavities. Cracks are open and have splintered or rounded edges. Stage 5: Bone is falling apart in situ, with large splinters lying around what remains of the whole, which is fragile and easily broken by moving. Original bone shape may be difficult to determine. Cancellous bone usually exposed, when present, and may outlast all traces of the former more compact, outer parts of the bones.
Vertebrate scavenging, in addition to affecting decomposition and insect colonization, may also produce postmortem artifacts that may be initially mistaken for wounds or mutilation. The same can be said for carnivore tooth marks which have been found on bones preserved in formations [10, 11, 12, 13, 14]. Conversely, wounds originally mistaken as rodent damage may actually have other causes [15].
Unlike weathering, ichnites are known as trace fossils or more broadly fossilized footprints, nests, dung, gastroliths, burrow, stomach contents. An ichnotaxon is defined by the International Code of Zoological Nomenclature as “a taxon based on the fossilized work of an organism” that is, the non-human equivalent of an artifact. Ichnites fit into several categories where dermestid beetles are thought to generally be associated. The common indications pertaining to systematic ichnology attribute discrete ovoid borings in bone and hollow, oval chambers with concave flanks bored into inner spongy and outer cortical bone surfaces. Cubiculum ornatus (insect- related ichnogenera) borings are interpreted as insect pupal chambers, based on close resemblance to modern arthropod pupae and ancient examples of bone-hosted pupal chambers [1]. Trace fossils referable to C. ornatus have been described previously by Tobien [3], Kitching [4].
Hypothesized tracemakers most commonly include carrion insects such as dermestid, silphid, and histerid beetles, tineid moths, and a variety of neotropical termite species [2, 5, 16, 17]. Roberts goes on to state that the most commonly cited trace fossil morphologies can be grouped into five general categories: 1) Cubiculum n. igen. (i.e., ovoid chambers); 2) shallow circular to elliptical pits; 3) starshaped pit marks; 4) Osteocallis n. igen. (i.e., surface trails); and 5) tunnels and subcortical cavities; and furthered that a variety of other bone borings which do not readily fit into these categories. More recently a forensic entomologist reported damage by D. maculatus larvae to both the humerus and the acetabulum of a human skeleton recovered from indoor conditions [6, 8]. Parkinson goes on to state that “unlike the vast body of palaeontological literature which suggests that dermestids modify bones in a number of distinctive ways, particularly the creation of pupation chambers or distinctive borings, the literature that relates to the theory makes absolutely no mention to such features.” More recently, it was established while investigating various skeletal preparation techniques that Dermestes beetles were capable of destroying bone, making grooves, holes and chew-marks [18]. However, providing scanning electron microscope images of the modifications identified, the qualitative descriptions provided by Fernández-Jalvo, et al. [18] were shown to be limited in their application for identification and particularly differentiation of dermestid modifications from other reported agents.
While working with dermestid colonies, Bader observed that dermestids often remove the periosteum (or outer surface layer) from cortical bone but noted that most destruction occurred on softer cancellous bone, particularly on bird bones, and also on articular facets of mammal bones [7]. However, to date no comprehensive descriptions of Dermestes modifications have been published that could be used to differentiate such modifications when compared to those created by other potential terrestrial invertebrate agents.
Given the absence of associated body fossils and the paucity of observational and experimental data on the morphology of dermestid borings [1], we also prefer to avoid definitively linking these traces from Cretaceous-age bones to dermestid beetles. However, given the clear and recurrent association with animal remains, we feel confident linking these traces to the activity of necrophagous or osteophagous carrion insect fauna [1].
Other works have briefly described insect borings which may also refer to Cubiculum. Schwanke, et al. [19] noted a similarity between certain cylindrical borings observed in Triassic vertebrate bones from Brazil and purported dermestid pupation chambers. Less clear, but possibly referable to Cubiculum, are large (1 cm-2.5 cm), circular (in cross section) borings documented in dinosaur bones from the Upper Cretaceous of Mongolia [7]. Though percussion marks on bone surfaces as a diagnostic of insect, canid, hominid behavior or otherwise, we aim to confirm what is currently seen specific to the feeding of Dermestes maculateus when left to feed en masse i.e. in colony. In particular, this research may prove useful in developing a geographical database of insect succession on carrion in a variety of habitats and scenarios in North America.
In large measure this question has been addressed and answered to a certain degree with primitive beetles and insects. However, there are still questions about whether we can see consistent evidence of this with extant dermestids species, although it’s not likely appreciated. Many of the aforementioned observations, while reported and accepted, lack experimental data. Across the conflicting nature of existing literature, termites, ants and beetles are by far the most widely accepted agents of bone modification. However, many potential limitations of such studies have been identified, such as a lack of standardized descriptive vocabulary, limited comparative case studies, localized applicability, insignificant sample sizes, unrepeatability, and use of single instead of multiple agents to gauge the frequency and intensity of different agents producing similar modification types [8].
Because Dermestes maculatus is one of the most ubiquitous dermestids in forensic settings, looking at potential bone feeding and marks from D. maculatus is of great practical interest. For this study the following hypotheses are posited for D. maculatus:
- The bone surface modification distribution and types produced are distinguishable as dermestid modifications.
- They will modify the surface of bones.
- They produce a variety of modifications on the surface of bones.
- They will modify bones in fresh /dry states of preservation, condition and of varying densities (thin cortical, thick cortical, and compact bone).
Material and Methods
Microscope and Imaging
Electron microscopy was used to provide necessary details of dermestid mouthpart morphology in both juvenile and adult stages of development to pair with any observed marks on bone surfaces. The targeted mouthpart areas of interest were enhanced including the pictured clypeus, labrum, mandible, segmented maxilla, and labial palps (Figures 1-4) (x-x) in both adult and juvenile specimens (Figures 5-9). Bird remains from a food processing plant with dermestid beetles being the confirmed de-fleshing agent were used in conjunction with extensively fed on bones in colony under controlled circumstances. The following sections detail the procedures used in preparing specimens for photography and study.
Procedures
The signals that derive from electron-sample interactions reveal information about the sample including external texture, chemical composition, and crystalline structure, and orientation of materials making up the sample. The Scanning Confocal Electron Microscope (SCEM) is an electron-optical implementation of the Scanning Confocal Optical Microscope (SCOM) which allows observation and characterization of sub-surface structures of thick, optically non-transparent materials. To determine whether or not marks had been made to bone surfaces, we used a Hitachi 3000 variable pressure scanning confocal electron microscope. The microscope featured high resolution thermionic electron scattering, which relayed the specimen chamber vacuum images in real time. This also allowed visual control around the sample from a range of different angles. The microscope additionally featured a high-density frame memory of 1280 x 960 pixels and an advanced image capture and archiving system for imaging and photography. Four-quadrant solid state backscatter allowed imaging in the compositional, 3D and topographic modes by manipulating samples used from each segment of the detector.
Beetles
Dermestes maculatus were obtained from colonies at the University of Nebraska-Lincoln (Lincoln, Nebraska). Three initial colonies were established in June 2015 with the intent of obtaining genetic homogeneity among test subjects. Colonies were kept in modified 25-gallon aquaria modified with mesh and lining to promote essential air circulation, confine insects to their enclosures, and to prevent the entrance of other unwanted organisms such as flies, ants and mites. Sealant was removed from the inner corners of each tank in order to prevent adults and larvae from ascending and escaping. To promote environmental conditions where beetles and larvae worked most efficiently, colonies were kept in dark rooms having independent temperature control. An ambient temperature ranging from 27-29 degrees Celsius was maintained using suspended 100-watt Exo Terra® Night heating lamps. Initial substrate consisted of a mixture of shredded paper and cottonwood bedding. A healthy population should be sufficiently large to ensure rapid cleaning of bones and tissues. To achieve a healthy and thriving population of adults, food was introduced as needed to further stimulate egg laying. After several generations, a population large enough to recruit adults for consistent experimentation became attainable.
Bones
Samples were taken from beef, pork, rodent, and chicken bone. However, due to the nature of delicate and intricate bone surfaces, microscopy was primarily focused on chicken bone. In combination, bones were both fresh and aged. Aged bones had been fed on by both adults and larvae. In addition to the bones we used for direct sampling, we gained material from a civil case in which there was a question regarding whether or not dermestid beetles had fed on the remains of a bird carcass. We used these remains as material to make a comparison to our controlled samples which were fed on by an approximate density of adult beetles and larvae in a 25-gallon aquarium enclosure. At combination of adult and immature individuals at a ratio of approximately 1:6 per container colony fed on tissues for a period of roughly two and a half months. These conditions in colony closely resemble what might be seen in an infestation in natural settings [20].
Our aim was to mirror natural conditions that would be found on a carcass with a dermestid infestation– and naturally dermestids, like most insects will remain on a food source until the resource has been exhausted. For this reason, the bones used in the study were exposed to adults and juveniles with tissues intact and without any additional preparations. Tissues were initially eaten down to tendons, ligaments, and cartilage before even these tissues were consumed. Once bare bones were observed (usually within one to two days) they were inspected for thorough cleaning. When it was determined all the tissues had been removed, beetles were placed back into the substrate where they remained until the time of preparation for microscopy.
In general, the charging effect is caused by the accumulation of static electric charges on the specimen surface. This can result in many problems which include damage, distortion, or even the eventual destruction of the specimen during observation under the ion beam. Avoid focused ion beam charging effects on specimens they were cleaned and heat dried through convection for a period of up to twenty four hours. Once the initial drying time had been attained a graded series of 70, 90, and 100% ethyl alcohol completed the dehydrating process. After dehydration a fixative osmium immersion was used to chemically dry and maintain the structural details of the samples.
An electrically conductive coating must be applied to electrically insulating samples for study in conventional SEM’s. To prevent charge build-up on electrically insulating materials, insect samples were sputter coated with a layer of gold as a conducting material. Since it was determined that the bones provided by the food manufacturing plant had reached desiccation no sample preparation was applied. All samples were mounted on double sided stubs with double sided conductive tape before being placed into the pressure chambers.
Analysis
In evaluating our images, we used fresh bones in comparison to old bones (both available and through images given in other publications) to gain an estimate of what beetles routinely did to the delicate surfaces through extensive feeding. Samples were visually inspected by more than one observer. All images show the pattern as imaged between 5 and 10 kilovolts at up to 12.0kX magnification with backscattered electron detection.
Results and Discussion
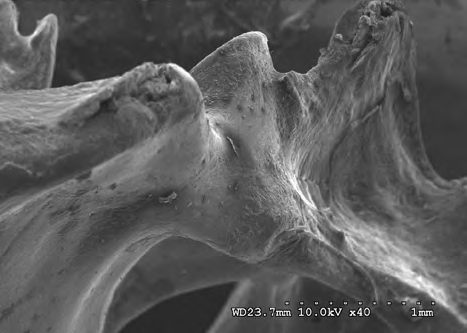
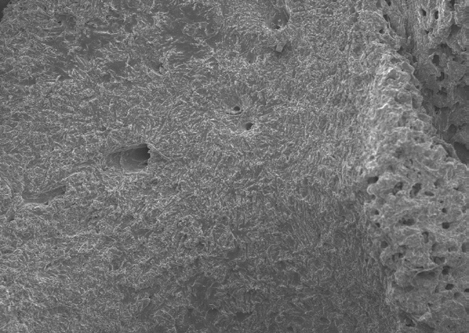
The results shown in Figures 10-13 indicate the linear ridge of the sacrum of the sample used from bird remains. In addition to the linear ridge, the image shows the depression and processes of the left ridge of the sacrum. Damage was determined to be consistent with stage 1 weathering.
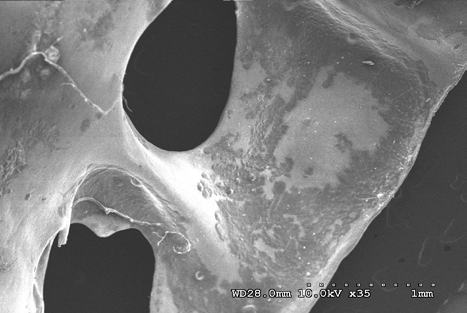
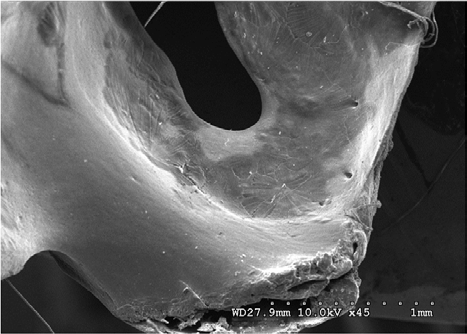
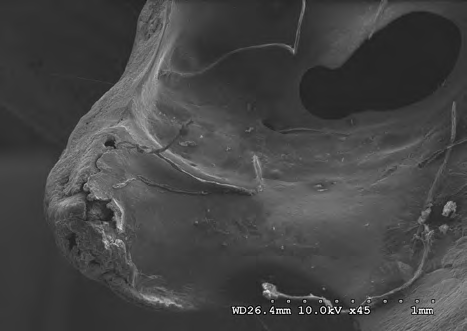
Figures 14-17 provides a dorsal view of stage 2 weathering on anterior sacral depression (bird remains sample) at 10 kV 45X magnification. The image details a pattern of wear showing no marks to the bone surface at increased magnification. Continued imaging along the same sample reveal the same consistencies and pattern of wear ending with fissures along the posterior sacral foramina on a depression at 10 kV 2.0kX magnification It is noted that on the bone surfaces in Figures 18-21 which were prepared with both heat and chemical drying processes there is an extensive pattern of wear consistent with stages 3 and 4 weathering. However, the absence of trace marks, mandibular grooves and furrows resulting from feeding parallels with our assumptions and observations.
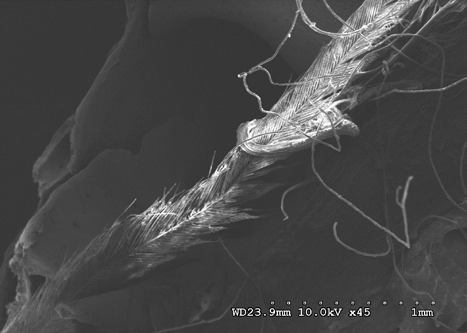
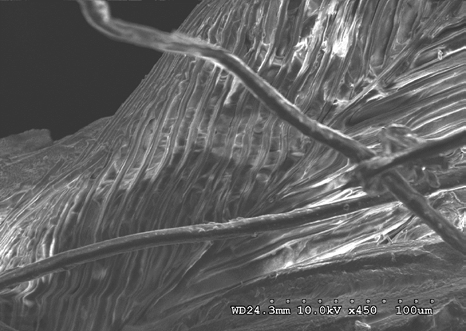
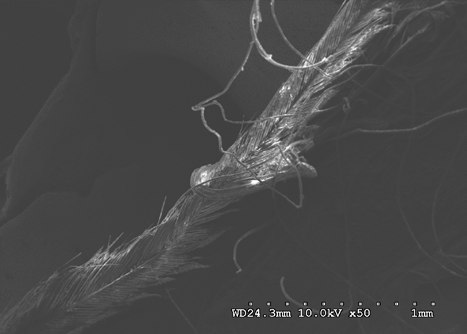
Figures 22-25 exhibit what were determined to be micro feathers and fibers derived from processing at a food plant. It is important to note that even on such delicate tissues and materials we see no damage. This observation illustrates the efficiency and near exactness of bone cleaning intrinsic to dermestid biology. Taken together, these observations indicate that, at magnifications on the order of 120X, the SEM imaging provides a reasonable conclusion that no markings have been made.
The role of demisted beetles in animal decomposition is well documented; they are known to consume soft sub- dermal tissue and skin, hence their name Dermestes derived from the Greek to “consume skin” [20, 21, 22]. Their involvement in animal decomposition and associated consumption of dry and decomposing animal matter typifies members of the Dermestes. Because of this they are routinely used for stripping carcasses of meat for skeletal collections and are widely considered as doing little damage to delicate bones.
The older literature largely denies any role of dermestids (principally, Dermestes maculatus) in altering bone. Howell [23] and Borell [24] state categorically that even skulls of minute sizes are cleaned without the slightest damage to the most delicate of processes. Howell goes on to state that, “…the tympanic bullae of mouse-sized skulls are infrequently eaten by the beetles, possibly in search of blood, processes are not broken off, delicate structures are not destroyed, teeth do not fall out, and sutures do not gape even in the youngest of specimens.” Similarly, Voorhies [25] states that after being exposed to dermestids for a number of weeks, even delicate bat skulls (e.g., Myotis sp.) have tissues removed without any harm to bone which was “cleaned to perfection”.
Among more recent workers, Osuji FNC [26] states that D. maculatus larvae may bore into the flesh of dried fish, but do not bore into either their bones or skulls. Similarly, modification of bones has not been regarded as an indicator of dermestid involvement by some forensic entomologists [22].
In forums among hobbyists and taxidermists who use dermestids it has been said that “[dermestids] will even eat their way through certain plastics in search of food sources given enough time, but bone remains clean, intact and ready to be bleached if desired” [27]. And many museums and institutions use D. maculatus colonies for tissue removal from bones prior to scientific analysis of those bones, implying an absence of any bone modification by D. maculatus.
In contrast, recent literature reports modifications to bone by terrestrial invertebrates, particularly insects including dermestids [8]. For some groups, such as the Tenebrionidae and Termitidae, the evidence of bone modification is thorough and convincing. However, for dermestids many reports of modification are from post hoc associations of dermestids with altered bones, rather than from direct experimental evidence. Even in the instances in which the potential for bone alteration by a dermestid has been made, it is unclear the species or conditions associated with the alteration.
Observations on rodents by Hefti, et al. [9], indicated that once available food sources were depleted, Dermestes maculatus beetles begin to destroy specific areas on bone, specifically the iliac crest of the pelvis, as well as vertebrae. They went on to state that when bones are modified by dermestids, the modifications are obvious. However, the observations Hefti, et al. [9] made did not exclude other possible causes of bone modification, and the reported ability or occurrence D. maculatus to modify bones was not confirmed.
Bones on carrion commonly exhibit distinctive weathering characteristics which can be related to the time since death and to the local conditions of temperature, humidity, and soil chemistry. These characteristics can be crucial in archeology and anthropology, as well as in forensic analysis. Indeed, distinguishing weathering from tool marks and other artifacts on bone is a common issue in forensic physical anthropology.
That D. maculatus feeding can leave marks on bones is established in the literature - that D. maculatus routinely mark bones is not. Our results here, exposing fleshed bones to D. maculatus adults and immatures for almost two months after complete tissue removal, indicate that under natural conditions D. maculatus do not feed on bones. Based on our initial hypothesis we can state that:
- the bone surface modification distribution and types produced are not distinguishable as dermestid modifications,
- D. maculatus does not modify the surface of bones,
- D. maculatus does not produce a variety of modifications on the surface of bones, and
- D. maculatus does not modify bones in fresh /dry states of preservation, condition and of varying densities (thin cortical, thick cortical, and compact bone).
The key issue seems to be the ability of D. maculatus to seek new food sources versus being confined (and eventually starving) on bones. In a natural setting, once a food supply is exhausted, D. maculatus would seek new food sources. If this process is somehow interrupted, such as through placement of an infested body in a sealed container or experimentally over a long (months to a year) period, published work suggests that D. maculatus could feed on bone. Based on this conclusion, whether in forensics analysis or paleontological analysis, we think the working assumption should be that D. maculatus does not alter bone, with the exception (as we outlined) when beetles and bones are in an enclosure that prevents beetle emigration [28, 29, 30, 31, 32].

















References
-
Roberts E, Rogers R, Foreman B (2007) Continental insect borings in dinosaur bone: Examples from the late Cretaceous of Madagascar and Utah. Journal of Paleontology 81(1): 201-208.
-
Behrensmeyer AK (1978) Taphonomic and ecologic information from bone weathering. Paleobiology 4(2): 150-162.
-
Tobien H (1965) Insect feeding marks on Tertiary and Pleistocene mammal bones. Senckenbergiana Lethaia 46: 441-445.
-
Kitching JW (1980) On some fossil arthropoda from the limeworks, makapansgat, Potgieterus. Palaeontologia Africana 23: 63-68.
-
Martin LD, West DL (1995) The recognition and use of dermestid (Insecta, Coleoptera) pupation chambers in palaeoecology. Palaeogeography, Palaeoclimatology, Palaeoecology 113(2-4): 303-310.
-
Schroeder H, Klotzbach H, Oesterhelweg L, Puschel K (2002) Larder beetles (Coleoptera, Dermestidae) as an accelerating factor for decomposition of a human corpse. Forensic Sci International 127(3): 231-236.
-
Kirkland JI, Bader K (2010) Insect trace fossils associated with Protoceratops carcasses in the Djadokhta formation (upper Cretaceous), Mongolia. Palaeogeography, Palaeoclimatology, Palaeoecology 311: 38-47.
-
Parkinson AH (2012) Dermestes maculatus and Periplaneta americana: bone modification criteria and establishing their potential as climatic indicators. MS Dissertation, University of the Witwatersstrand, USA, pp: 1-204.
-
Hefti E, Trechsel U, Ruffenacht H, Fleisch H (1980) Use of dermestid beetles in cleaning bones. Calcified Tissue Institute 31(1): 45-47.
-
Rogers RR, Krause DW, Rogers KC (2003) Cannibalism in the Madagascan dinosaur Majungatholus atopus. Nature 422(6931): 515-518.
-
Dillon LC, Anderson GS (1995) Forensic entomology: the use of insects in death investigations to determine elapsed time since death. Canadian Police Research Centre Ottawa, Ontario.
-
Dillon LC, Anderson GS (1996) Forensic entomology: a database for insect succession on carrion in Northern and Interior BC Technical report TR-04-96. Canadian Police Research Centre, Ottawa, Ontario.
-
Dillon LC, Anderson GS (1997) Forensic entomology- use of insects towards illegally killed wildlife. Technical report, World Wildlife Fund, Toronto, Ontario pp: 12.
-
Patel F (1994) Artefact in forensic medicine: postmortem rodent activity. Journal of Forensic Sciences 39(1): 257- 260.
-
Patel F (1995) Artefact in forensic medicine: pseudo- rodent activity. Journal of forensic sciences 40(4): 706- 707.
-
Rogers RR (1992) Non-marine borings in dinosaur bones from the Upper Cretaceous Two Medicine Formation, northwestern Montana. Journal of Vertebrate Paleontology 12(4): 528-531.
-
Martha T (1994) Bone weathering in the tropical rain forest. Journal of Archaeological Science 21(5): 667-673.
-
Fernandez-Jalvo Y, Monfort MDM (2008) Experimental taphonomy in museums: Preparation protocols for skeletons and fossil vertebrates under scanning electron microscopy. Geobios 41(1): 157-18.
-
Schwanke C, Kellner AWA (1999) Sobre o primeiro registro de Synapsida no Triássico basal do Brasil. 16th Congresso Brasileiro de Paleontologia, Boletim de resumos, 101.
-
Byrd J, Castner J (2001) Forensic entomology. The utility of arthropods in legal investigations. CRC Press, Washington, DC, USA, pp: 620.
-
Cornaby BW (1974) Carrion reduction by animals in contrasting tropical habitats. Biotropica 6(1): 51-63.
-
Smith KGV (1986) A manual of forensic entomology. Cornell University Press, Ithaca, NY, USA.
-
Hall ER, Russell WC (1933) Dermestid beetles as an aid in cleaning bones. J Mammalogy 14(4): 372-374.
-
Borell AE (1938) Cleaning small collections of skulls and skeletons with dermestid beetles. Journal of Mammology 19(1): 102-110.
-
Voorhies CT (1948) A chest for dermestid cleaning of skulls. J Mammalogy 29(2): 188-189.
-
Osuji FNC (1975) The effects of salt treatment of fish on the developmental biology of Dermestes maculatus (Coleoptera, Dermestidae) and Necrobia rufipes (Cleridae). Entomologia Experimentalis et Applicata 18(4): 472-479.
-
Ceciliason A, Andersson G, Lindström A, Sandler H (2018) Quantifying human decomposition in an indoor setting and implications for postmortem interval estimation. Forensic Sci. International 283: 180-189.
-
Hinshaw SII (2003) Use of dermestid beetles for skeleton preparation.
-
Ryan MJ, Chinnery-Allgeier BJ, Eberth DA (2010) New perspectives on horned dinosaurs. Indiana University Press, Bloomington, India.
-
Sommer HG, Anderson S (1974) Cleaning skeletons with dermestid beetles—two refinements in the method. Curator: The Museum J 17(4): 290-298.
-
West DL, Hasiotis ST (2007) Trace fossils in an archaeological context: examples from bison skeletons, Texas, USA. In: Miller W (Ed.), Trace Fossils: Concepts, Problems, Prospects. Elseiver, pp: 545-561.
-
Osuji FNC (1975) The effects of salt treatment of fish on the developmental biology of Dermestes maculatus (Coleoptera, Dermestidae) and Necrobia rufipes (Cleridae). Entomologia Experimentalis et Applicata 18(4): 472-479.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework