Diversity of Ants (Hymenoptera: Formicidae) Captured by Myrmeleon (Neuroptera: Myrmeleontidae) Larvae in a Ecosystem of the Brazilian Cerrado Biome
This study aimed to investigate the diversity of ants captured in Myrmeleon pits in a Cerrado environment (Brazilian savanna) and assess the relationship between pit size and capture success. Field expeditions were performed in the Inhamum Municipal Environmental Protection Area, Caxias, Maranhão State, Brazil. Pits of Myrmeleon larvae were observed, and captured ants were collected and identified. Our results showed that Myrmeleon larvae inhabiting this Cerrado site rely on ants as their main natural prey. Seven ant species were identified. There was a positive relationship between pit diameter and capture success. This is the first investigation of the diversity of ants naturally predated by antlions.
Introduction
Larvae of the genus Myrmeleon Linnaeus (Neuroptera: Myrmeleontidae), commonly known as antlions, are round shaped and have long, sickle-like jaws that assist in the absorption of body fluid from their prey [1, 2]. The predation strategy of antlion larvae consists in the construction of funnel-like pits to capture prey [3, 4, 5]. The larva’s abdomen is used as a hoe to excavate sandy soil and the mouth to cast sand out from the pit [6].
Antlion larvae are generalist predators of small arthropods. However, because ants are highly abundant in most natural environments, they are antlions’ main prey Gotelli NJ [7], Gotelli NJ [8], hence the common name. When a prey falls into the trap, the antlion larva hurls sand particles to make it slide to the bottom of the pit. Then, the larva bites the prey to inject digestive saliva and uses its jaw to weaken and bury the captured insect, leading to death. After all body fluids are consumed, antlions discard the carcasses outside the pit [9, 10, 11, 12]. The predation success of antlion larvae can be affected by biotic factors, such as prey availability and escape tactics, and abiotic factors, such as temperature, type of substrate, pit location, and soil moisture [3, 5, 13].
Ants are one of the most numerous and diverse groups of organisms in tropical biomes. Therefore, members of the Formicidae family are extremely important for ecosystem function Hölldobler B, et al. [14], Fernández F [15] and have diverse types of interactions with other organisms. For instance, ants are a major source of food for several animal species (myrmecophagy) [16, 17]. Antlions are one of the major ant-eating insects in open ecosystems such as the Cerrado (Brazilian savanna). The presence of antlions has important impacts on the behavior, population control, and territorial distribution of epigeic ants Gotelli NJ [8], Johnson RA [18], Freire LG [19], including invasive species [20].
The biological and behavioral aspects of Myrmeleon under natural conditions are poorly described in the literature [8, 21, 22]. Information on the diversity of taxa naturally predated by antlion larvae is particularly lacking. Therefore, this study aimed to investigate the diversity of Formicidae captured in pits of Myrmeleon larvae in a Cerrado environment and analyze the relationship between pit size and capture success.
Material and Methods
Sampling was performed at the Inhamum Municipal Environmental Protection Area (04°53′S 43°24′W, 66 m elevation), municipality of Caxias, Maranhão State, Brazil (Figure 1). This conservation unit covers an area of about 4,500 ha. The vegetation is a mosaic of typical Cerrado plant communities, ranging from savanna phytophysiognomy (trees and shrubs dispersed in a grassy stratum) to dense forest (continuous tree canopy) [23]. The region is under tropical savanna climate (Aw) Köppen W, et al. [24] with an average annual temperature of 27°C, relative humidity of 70–73%, and precipitation of 1600–2000 mm. There are two well-defined seasons, a rainy season from December to June and a dry season from July to November [25, 26].
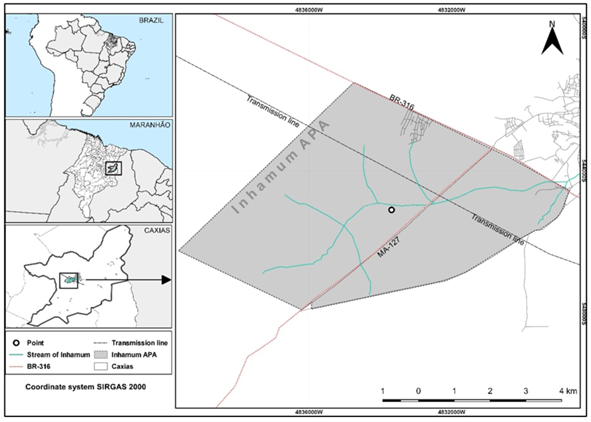
Two expeditions were performed in December 2019 (rainy season). Clusters of Myrmeleon pits were located at the border of a hiking trail. First, pit diameter was measured using a caliper. Then, pits were observed during 4 h (from
8:00 a.m. to 12:00 p.m.) and prey naturally captured by antlion larvae were collected. Captured prey included all individuals that fell into the pit and were immobilized by Myrmeleon larvae; individuals that escaped the attack were not considered. A total of 12 pits were observed during the first expedition, and 17 during the second.
Collected specimens were removed from the bottom of the pit by using entomological forceps and placed in Falcon tubes containing 70% alcohol. Three specimens of Myrmeleon larvae were also collected on each day of observation for genus confirmation. All biological material was transported to the Laboratory of Myrmecology of the State University of Maranhão (LAMIR/UEMA), Caxias, Brazil.
Ants were initially identified at the genus level using dichotomous keys [27, 28, 29]. Confirmation of species and morphospecies was achieved through direct comparison with reference materials from the myrmecological collection of LAMIR/UEMA, where the collected specimens were deposited. Other insects (beetles, bugs, and cockroaches) were identified at the family level using the dichotomous keys of Rafael JA, et al. [30].
Pits were classified according to the presence or absence of predated individuals, and differences in pit size between groups were assessed by Student’s t-test [31]. Pearson’s correlation tests Morettin PA, et al. [31] were used to investigate the relationship between pit size and capture success. All statistical analyses were performed using R software [32].
Results
Seventeen ant species predated by Myrmeleon were collected and distributed in four subfamilies, six genera, and seven species (Table 1). In addition to ants, four individuals from other taxonomic groups were collected: a bug nymph (Hemiptera: Reduviidae), a beetle (Coleoptera: Tenebrionidae), and two cockroach nymphs (Blattodea: Blaberidae).
| Subfamily/ant species | Number of Individuals |
|---|---|
| MYRMICINAE | |
| Cephalotes pusillus(Klug,1824) | 1 |
| Pheidole sp. fallax group | 4 |
| FORMICINAE | |
| Brachymyrmex heeriForel,1874 | 2 |
| Camponotus crassusMayr,1862 | 7 |
| Camponotus novogranadensisMayr,1870 | 1 |
| ECTATOMMINAE | |
| Ectatomma muticumMayr,1870 | 1 |
| PSEUDOMYRMECINAE | |
| Pseudomyrmex oculatus(Smith,1855) | 1 |
| Non-formicid insects | Number of individuals |
| BLABERIDAE | 2 |
| TENEBRIONIDAE | 1 |
| REDUVIIDAE | 1 |
Table 1: Diversity of taxa captured in natural pits of Myrmeleon larvae in a Cerrado environment in Caxias, Maranhão, Brazil.
Twenty-nine Myrmeleon larvae pits were observed in sandy soil only, revealing an aggregated distribution pattern. Pit diameter ranged from 5 to 50 mm, with a mean of 21.93 mm. In total, 21 prey were successfully captured, that is, 0.04 individuals/min in Myrmeleon pit aggregates. However, only 44.82% of pits were successful in capturing at least one prey; that is, 16 larvae traps did not catch prey during the observation period.
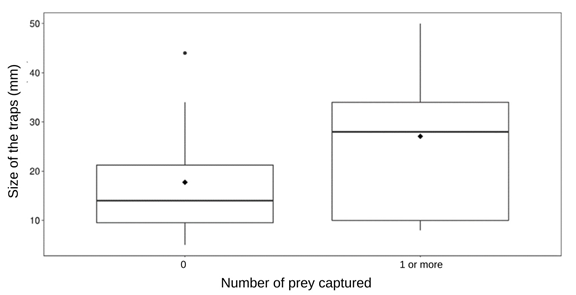
Table 2 & Figure 2 show comparisons between successful (1 prey or more) and unsuccessful (0 prey) traps. Unsuccessful traps (n = 16) had a mean size (diameter) of 18 mm and successful traps (n = 13) were sized on average 27 mm. Therefore, unsuccessful traps were generally smaller than successful traps, although differences in diameter were not significant (p = 0.069).
| n | Mean | Standard deviation | Minimum | First quartile | Median | Third quartile | Maximum | |
|---|---|---|---|---|---|---|---|---|
| 0 | 16 | 17.8 | 10.8 | 5 | 9.5 | 14 | 21.2 | 44 |
| 1 or more | 13 | 27.1 | 14.6 | 8 | 10 | 28 | 34 | 50 |
Table 2: Descriptive statistics of Myrmeleon pit size (mm) by presence or absence of trapped prey (p = 0.069).
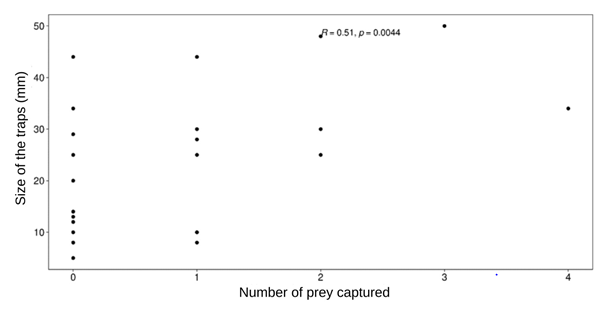
When considering the total number of pits (n = 29), we observed a positive correlation between pit diameter and capture success (r = 0.51, p = 0.0044, Figure 3). This result shows that the larger the Myrmeleon larvae pit diameter, the higher the chance of success in capturing prey.
Discussion
Our results showed that, in the Brazilian Cerrado, ants of different species are the major prey of Myrmeleon larvae. We observed a positive correlation between pit diameter and predation success. However, no significant differences in pit size were observed when comparing successful traps and unsuccessful traps.
Camponotus crassus was the most frequent ant species predated by Myrmeleon larvae. The high frequency is due to the wide occurrence of C. crassus in the Cerrado, including in Cerrado areas in Maranhão State, Brazil [33, 34]. The ant species constantly exploits plant resources (such as extrafloral nectaries), which demands a large number of workers [35]. During foraging, the ants are at risk of becoming victims of antlion traps. As reported by Glenn S, et al. [20], Camponotus ants, members of the subfamily Dolichoderinae, are easier to prey because of their thin cuticle and lack of abdominal protection compared with subfamilies of Formicidae that have thick cuticles. These ants also lack a sting Santos JC, et al. [36] possibly limiting their defense repertoire against predator attacks.
Pheidole sp. fallax group, the second most frequently predated taxon, are small individuals quite common in the studied preservation area (Andrade & Martins, unpublished). Gotelli NJ [8] found that species of the genus Pheidole are affected by the presence of Myrmeleon larvae pits. The abundance of the ant species is reduced in areas with wide distribution of larvae pits. The P. fallax group represents cosmopolitan ants with populous nests, which explains the high number of individuals sampled in the present study [37, 38, 39]. Thus, we highlight that Camponotus and Pheidole are important genera in the diet of Myrmeleon larvae in the Brazilian Cerrado.
Beetles and bugs were previously reported as non- formicid prey of Myrmeleon larvae [5]. Here, we report for the first time that cockroaches are a natural source of food for Myrmeleon. Given that predators generally respond more quickly to encounters with frequently consumed prey Uetz GW [40], Morse DH [41] it is expected that antlion larvae respond faster to the presence of ants than to prey that rarely fall into their traps. Future studies can test this hypothesis by quantifying the capture success of ants and other types of prey by antlion larvae.
The observed aggregated distribution pattern of Myrmeleon larvae pits is the result of habitat selection. Larvae seek ideal conditions for the construction of pits. When aggregations occur, larger larvae tend to be more effective in capturing prey than smaller larvae, as body size is correlated with pit diameter and larger traps can intercept a larger number of prey [7, 10, 12, 22, 42]. Our results showed that this correlation, albeit moderate, occurred in Myrmeleon pit clusters in the study area. However, the relationship between pit size and predation success was not observed when comparing successful and unsuccessful traps. This result might be related to the low natural predation rates of antlions.
We found that 0.90 prey were captured per pit per hour. Similar results were obtained by Heinrich and Heinrich B, et al. [5] in investigating the natural predation of Myrmeleon immaculatus DeGeer in a coastal area of the United States of America. The low frequency of antlion predation under natural conditions was associated with the presence of larvae at early stages of development. During this phase, larvae are not very efficient in capturing prey, which can lead to the starvation and, ultimately, death of many individuals [2].
Another important factor observed in the field was the large number of escapes. In general, ants avoided Myrmeleon traps, and even when they fell into the pit, many managed to escape the attacks of larvae. The capture rates of antlion larvae are quite low under natural conditions [2, 5]. Ants’ ability to evade traps, known as avoidance behavior, is the main cause of low capture rates [8, 43]. Gotelli NJ [8] reported that ants may emit chemical signals to warn nestmates of Myrmeleon pit agglomerations, as evidenced by the reduced foraging activity in areas with traps.
Antlion larvae can perceive the presence of prey through the vibration of soil particles [44, 45]. Thus, even before a prey falls into the pit, the larva is aware of its presence. According to Guillette LM, et al. [46], animals rely on past experiences to evaluate their chance of predation success and adjust their behavior in order to obtain a higher energy return. Thus, when predators realize that certain prey cannot be captured, they choose not to attack to save energy [47].
This is a pioneering study on the diversity of formicids naturally predated by Myrmeleon larvae. Our results show aspects of the biology of Myrmeleontidae in the Brazilian Cerrado, contributing to the knowledge of the natural behavior of antlions, which is poorly documented.
Acknowledgments
This study was supported by the following Brazilian institutions: Foundation to Support Research and Scientific and Technological Development of Maranhão (FAPEMA) and Postgraduate Program in Biodiversity, Environment and Health at the State University of Maranhão (PPGBAS/UEMA). The authors would like to thank Antonio F. S. de Sousa for his collaboration during field collections.
Author Contributions
Thito Thomston Andrade: Investigation, Formal analysis, Writing - Original draft. Luiza Carla Barbosa Martins: Conceptualization, Methodology, Investigation, Writing - Review & Editing. Tatiane do Nascimento Lima: Formal analysis, Writing - Review & Editing. Vinícius Albano Araújo: Formal analysis, Writing - Review & Editing.
References
-
Humeau A, Rougé J, Casas J (2015) Optimal range of prey size for antlions. Ecological Entomology 40(6): 776-781.
-
Nonato LM, Lima TN (2011) Predation behavior of the Myrmeleon brasiliensis (Neuroptera: Myrmeleontidae) larval instars. Revista Colombiana de Entomologia 37(2): 354-356.
-
Griffiths D (1980) The feeding biology of ant-lion larvae: prey capture, handling and utilization. Journal of Animal Ecology 49(1): 99-125.
-
Griffiths D (1986) Pit construction by antlion larvae: a cost-benefit analysis. Journal Animal Ecology 55(1): 39- 57.
-
Heinrich B, Heinrich MJ (1984) The pit-trapping foraging strategy of the ant lion, Myrmeleon immaculatus DeGeer (Neuroptera: Myrmeleontidae). Behavioral Ecology and Sociobiology 14(2): 151-160.
-
Fertin A, Casas J (2006) Efficiency of antlion trap construction. Journal of Experimental Biology 209(18): 3510-3515.
-
Gotelli NJ (1993) Ant lion zones: causes of high‐density predator aggregations. Ecology 74(1): 226-237.
-
Gotelli NJ (1996) Ant community structure: effects of predatory ant lions. Ecology 77(2): 630-638.
-
Morrison LW (2004) Spatiotemporal variation in antlion (Neuroptera: Myrmeleontidae) density and impacts on ant (Hymenoptera: Formicidae) and generalized arthropod foraging. Annals of the Entomological Society of America 97(5): 913-922.
-
Scharf I, Ovadia O (2006) Factors influencing site abandonment and site selection in a sit-and-wait predator: a review of pit-building antlion larvae. Journal of Insect Behavior 19(2): 197-218.
-
Scharf I, Barkae ED, Ovadia O (2010) Response of pit- building antlions to repeated unsuccessful encounters with prey. Animal Behaviour 79(1): 153-158.
-
Dias SC, Santos BA, Werneck F, Lira PK, Carrasco- Carbadillo V, et al. (2006) Efficiency of prey subjugation by one species of Myrmeleon larvae (Neuropetra: Myrmeleontidae) in the central Amazonia. Brazilian Journal of Biology 66(2): 441-442.
-
Arnett AE, Gotelli NJ (2001) Pit-building decisions of larval ant lions: effects of larval age, temperature, food, and population source. Journal of Insect Behavior 14(1): 89-97.
-
Hölldobler B, Wilson EO (1990) The Ants. Cambridge, Harvard University Press, pp: 732.
-
Fernandez F (2003) Introduction to ants of the Neotropical region. Instituto Humboldt, Bogotá, Colombia, pp: 424.
-
Delabie JHC (1999) Aspects of myrmecophagy in the Neotropical Region. Naturalia 24: 225-231.
-
Delabie JHC, Jahyny B (2007) The animal myrmecosphere: relations of dependence between forms and other animals. O Biológico 69(2): 7-12.
-
Johnson RA (2001) Biogeography and community structure of North American seed-harvester ants. Annual Review of Entomology 46(1): 1-29.
-
Freire LG, Lima TDN (2019) Effect of rain on trap building by Myrmeleon brasiliensis in a riparian woodland from the Cerrado biome in Brazil. Entomologia Experimentalis et Applicata 167(6): 561-565.
-
Glenn S, Holway D (2008) Consumption of introduced prey by native predators: Argentine ants and pit-building ant lions. Biological Invasions 10(3): 273-280.
-
Gatti MG, Farji‐Brener, AG (2002) Low density of ant lion larva (Myrmeleon crudelis) in ant‐acacia clearings: high predation risk or inadequate substrate. Biotropica 34(3): 458-462.
-
Lima TDN, Faria RR (2007) Microhabitat selection by ant- lion larvae Myrmeleon brasiliensis (Návas) (Neuroptera: Myrmeleontidae), in a Forest Reserve, Aquidauana, MS / Microhabitat selection by ant-lion Larvae Myrmeleon brasiliensis (Návas) (Neuroptera: Myrmeleontidae), in a Forest Reserve, Aquidauana, State of Mato Grosso do Sul. Neotropical Entomology 36(5): 812-814.
-
Ribeiro JF, Walter BMT (2008) The main phytophysiologies of the Cerrado biome. In: Sano SM, Almeida SP (Eds.), Closed: environment and flora. Planaltina, Brazil, pp: 153-212.
-
Köppen W, Geiger R (1928) Climate of the Erde. Gotha: Verlag Justus Perthes.
-
Conceição GM, Ruggieri AC, Silva EO, Gomes EC, Roche HMV (2011) Plant species and dispersal syndromes of the municipal environmental protection area of Inhamum, Caxias, Maranhão, Brazil. Ambiente & Água- An Interdisciplinary Journal of Applied Science 6(2): 129-142.
-
Albuquerque AB (2012) River bridge and the municipal environmental protection area of Inhamum. In: Caxias MA, Barros MC (Eds.), Biodiversity of the Inhamum municipal environmental protection area, São Luís: UEMA, Brazil, pp: 107-116.
-
Bolton B (1994) Identification guide to the ant genera of the world. Massachusetts. Harvard University Press, pp: 222.
-
Bolton B (2003) Synopsis and classification of Formicidae. Memoirs of the American Entomological Institute pp: 370.
-
Baccaro FB, Feitosa RM, Fernández F, Fernandes IO, Izzo TJ, et al. (2015) Guide to ant genera in Brazil. INPA Publisher, Manaus, Brazil.
-
Rafael JA, Melo GAR, Carvalho CJB, Casari SA, Constantino R (2012) Insects from Brazil: diversity and taxonomy. Ribeirão Preto. Holos Editora pp: 810.
-
Morettin PA, Bussab WO (2013) Basic Statistics. In: 8th (Edn.), Saraiva Publishing.
-
Team CR (2022) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna.
-
Brandao CRF, Silva RR, Feitosa RM (2011) Cerrado ground-dwelling ants (Hymenoptera: Formicidae) as indicators of edge effects. Zoologia (Curitiba) 28(3): 379-387.
-
Prado LPD, Feitosa RM, Triana SP, Gutiérrez JAM, Rousseau GX, et al. (2019) An overview of the ant fauna (Hymenoptera: Formicidae) of the state of Maranhão, Brazil. Papéis Avulsos de Zoologia pp: 59: 38.
-
Oliveira OS, Brandão CRF (1991) The ant community associated with extrafloral nectaries in Brazilian cerrados. In: Cutler DF, Huxley CR (Eds.), Ant-Plant Interactions. Oxford, Oxford University Press, USA pp: 198-212.
-
Santos JC, Del-Claro K (2009) Ecology and behaviour of the weaver ant Camponotus (Myrmobrachys) senex. Journal of Natural History 43(23-24): 1423-1435.
-
Marinho CG, Zanetti R, Delabie JH, Schlindwein MN, Ramos LDS (2002) Diversity of ants (Hymenoptera: Formicidae) from leaf litter in eucalyptus forests (Myrtaceae) and cerrado areas in Minas Gerais. Neotropical Entomology 31(2): 187-195.
-
Ramos LDS, Filho RZ, Delabie JH, Lacau S, Santos MDFS (2003) Litter ant (Hymenoptera: Formicidae) communities in “stricto sensu” cerrado areas in Minas Gerais. Lundiana 4(2): 95-102.
-
Tibcherani M, Nacagava VAF, Aranda R, Mello RL (2018) Review of ants (Hymenoptera: Formicidae) as bioindicators in the Brazilian Savanna. Sociobiology 65(2): 112-129.
-
Uetz GW (1992) Foraging strategies of spiders. Trends in Ecology & Evolution 7(5): 155-158.
-
Morse DH (2000) The effect of experience on the hunting success of newly emerged spiderlings. Animal Behaviour 60(6): 827-835.
-
Day MD, Zalucki MP (2000) Effect of density on spatial distribution, pit formation and pit diameter of Myrmeleon acer Walker, (Neuroptera: Myrmeleontidae): patterns and processes. Austral Ecology 25(1): 58-64.
-
Lucas JR (1989) The structure and function of antlion pits: slope asymmetry and predator-prey interactions. Animal Behaviour 38(2): 318-330.
-
Devetak D, Ncinger-Vracko B, Devetak M, Marhl M, Spernjak A (2007) Sand as a medium for transmission of vibratory signals of prey in antlions Euroleon nostras (Neuroptera: Myrmeleontidae). Physiological Entomology 32: 268-274.
-
Mencinger-Vracko B, Devetak D (2008) Orientation of the pit-building antlion larva Euroleon (Neuroptera, Myrmeleontidae) to the direction of substrate vibrations caused by prey. Zoology 111(1): 2-8.
-
Guillette LM, Hollis KL, Markarian A (2009) Learning in a sedentary insect predator: Antlions (Neuroptera: Myrmeleontidae) anticipate a long wait. Behavior Process 80(3): 224-232.
-
Borror DJ, Triplehorn DJCA, Johnson NF (2005) Borror and DeLong’s Introduction to the Study of Insects. In: 7th (Edn.), Thompson Brooks, Belmont, CA, USA.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework