Studies of Serrasentis sagittifer (Linton, 1889) Van Cleave, 1923 from cobia Rachycentron canadum (Linn.) in the Arabian Gulf; with Molecular Analysis and Optical Microscopy
The finding of a population of Serrasentis sagittifer (Linton, 1889) Van Cleave, 1923 in Cobia Rachycentron canadum (Linn.) in the Arabian Gulf off Iranian waters prompted the study of its molecular parameters known previously only from a population of adults from the same host species, and of immatures from northern Australian waters and from other hosts in the Red Sea. We have previously described S. sagittifer from Cobia and from paratenic hosts in the same Gulf waters off Kuwait and Qatar and detailed its worldwide morphological variability and geographical distribution. We have processed and sampled a few specimens from the Arabian Gulf off Iran for morphological verification of their diagnosis and offered a few light microscopical images that accentuate their identity in Iranian waters for the first time. We document additional diagnostic features of S. sagittifer using optical microscopy. We determine that the distribution of S. sagittifer runs parallel that of its primary adult host, R. canadum, worldwide. We have used the 18S and 28S gene regions of ribosomal RNA, including the mitochondrial cox1 gene, for molecular analysis of an Arabian Gulf population for the first time. Maximum likelihood and Bayesian inference methods were used for phylogenetic reconstructions, the sequences of S. sagittifer from Iran were grouped within the genus Serrasentis and placed as a sister group to S. nadakali George and Nadakal, 1978. Due to the lack of molecular data for other species of this group of Rhadinorhynchidae, the present phylogenetic inferences are required to continue exploring the genetic diversity of this group of species to infer their phylogeny. The cox 1 haplotype network inferred with 14 sequences of S. sagittifer revealed 13 haplotypes separated from each other by a few substitutions. Our ocular microscopy accentuated morphological details of S. Sagittifer and the haplotype network genealogy produced in this study based on mt cox1 sequences from S. sagitiffer show a phylogeographic structure for the first time; therefore, the haplotypes grouped into their own geographic clusters.
Omar M Amin1*, Anshu Chaudhary2, Milad Badri3, Aida Vafae Eslahi3 and Hirdaya S Singh2,4
¹Institute of Parasitic Diseases, Scottsdale, Arizona, USA
Introduction
The genus Serrasentis Van Cleave, 1923, has a confused taxonomic history as it was originally established as the genus Echinogaster by Monticelli [1]. Afterwards Porta [2] compact the genus Echinogaster to a subgenus of Echinosoma Porta, 1907 [2] and also added Echinorhynchus (=Serrasentis) sagittifer Linton, 1889, to his new genus [3]. Subsequently, Van Cleave established the genus Serrasentis as a nom. nov. for the reason that the name Echinogaster was also previously occupied. Furthermore, Van Cleave [4] included in the valid genus Serrasentis two known species, i.e., Serrasentis lamelliger (Diesing, 1854) Van Cleave, 1924 [4], and Serrasentis socialis (Leidy, 1851) Van Cleave, 1924 [4]. Afterwards, the species names were searched by Van Cleave and Lincicome [5] with the name Serrasentis sagittifer having priority over S. socialis, and approving that the samples he had inspected were in fact of S. sagittifer. In the family Isthmosacanthidae, the four accepted genera are: Golvanorhynchus Noronha, Fabio and Pinto, 1987 [6], Gorgorhynchoides Cable and Linderoth, 1963 [7], Isthmosacanthus Smales, 2012 [8], and Serrasentis [8, 9].
Adults of Serrasentis sagittifer are most common in cobia Rachycentron canadum (Linn.) worldwide. The geographical distribution of S. sagittifer was comparable to that of the primary definitive host, R. canadum. It was described morphologically in the Atlantic North and South America, the Atlantic west coast of West Africa, the Indian Ocean to East Africa, the Red Sea and the Arabian Gulf, the Indo-Pacific Ocean in Australia, Indonesian islands, South China Sea and Vietnam, and in unusual localities in California, Oregon, and Washington [10]. Our treatment [10] included the history of research on S. sagittifer in all these locations. We described the adult population from R. canadum and of the immatures from 8 species of paratenic hosts (Bothus sp. Rafinesque (Bothidae) Lutjanus coccineus (Cuvier) (Lutjanidae), L. fulviflamma (Forsskål), Nemipterus japonicus (Bloch) (Nemipteridae), N. peronii (Valenciennnes), Otolithus argenteus (Cuvier) (Otolithidae), Upoeneus sulphureus Cuvier (Mullidae), and Pseudorhombus arsius (Hamilton) (Paralichthyidae) in the Arabian Gulf of Kuwait and Qatar [10]. Our description was supplemented with (1) 17 SEM images, (2) a detailed table of comparative morphometrics of juveniles and adults from Arabian Gulf off Kuwait and Qatar [10], Northern Australia [11], Atlantic North America [12, 13, 14], and Veracruz, Mexico [15], (3) three X-ray panels of elemental scans in hook tips, center and edge of mid-hooks, and in edge of hook base were analyzed for sulfur, calcium, and phosphorus contents; the chemical footprint of the species. (4) An analysis of the micropore pattern and its association of the internal crypts associated with the differential absorption of nutrients.
The only other land-mark contribution to the understanding of S. sagittifer is the redescription by Barton, et al. [11] who on page 120 described their Australian specimens as having “Genital pore subterminal in both sexes (figs. 5, 10)” while their own figures 2 & 10 clearly show terminal gonopore in males as reported in all other descriptions and in our own specimens. Their line drawing (fig. 11) shows an oblong egg while eggs in our specimens were considerably more elongated. More importantly, they state that posterior hooks have “discoid roots” but their figures 8 and 9 depict hooks 13-16 to be rootles (Barton, et al. [11], page 123). Actually, in our specimens, it is these hooks 13-16 that have short anteriorly directed roots and it is the next anterior 4 hooks that have discoid roots. The line drawings of proboscis hooks in Golvan (1956 [12], Plate 4b, c, and 1969, fig. 86), Golvan and Houin (1964 [14], fig. 33), and Fonesca et al. (2019 [16], fig. 1c), did not show hook roots. Salgado Maldonado’s [15] and Golvan [17] provided a good species descriptions and Salgado Maldonado [15] included measurements of dorsal vs. ventral hooks but, like most other observers, they made no reference to or line drawings of hook roots. Golvan’s [17] image of an ovoid egg (fig. 84c), like that in Barton, et al. [11], was not accurate. Other well-studied populations include the one from Brazil by Travassos [18] who initially quoted Golvan’s [12] description and provided excellent line drawings. Travassos [18] made no reference to hook roots either and confused the diverticula with protonephridia that are not found in Paleacanthocephala. Travassos (1966 [18], page 7 and uninformative fig. 1) also erred in stating that the “filamentous cement glands are arranged in two more or less sheathed systems of at least 6 glands that are partially fused during their course towards the posterior end;” the number of cement glands is 4.
We include with our molecular analysis, light microscopy images of diagnostic features of our Iranian specimens of S. sagittifer to augment the SEM images published in Amin, et al. [10]. Barton, et al. [11] also provided baseline 18S rDNA, 28S rDNA, and Cox 1 sequence data for a diverse collection of specimens (2 adults and 8 cystacanths) from 4 different species of host fish (R. canadum, Lutjanus sebae, Lutjanus malabaricus, and Lethrinus laticaudis) in 3 different geographical locations (Northern Territory, Western Australia, Queensland). Abdel-Ghaffar, et al. [19] also provided molecular analysis of S. sagittifer from gilthead sea bream Sparus aurata Linn. in the Red Sea using 18S rDNA sequence.
In order to correctly identify the present acanthocephalan specimens to species level beyond ocular microscopy, we used the 18S and 28S subunit ribosomal DNA (28S) and mitochondrial cytochrome c oxidase subunit 1 (cox_1) for sequencing and haplotype analysis. Phylogenetic analyses were also executed based on the 18S and _cox1 sequence data using maximum likelihood (ML) and Bayesian inference (BI) approaches to elucidate the phylogenetic relationships between the congeners of S. sagittifer and other Serrasentis species. Analysis of the cytochrome oxidase subunit 1 (cox_1) of _S. sagittifer demonstrated the presence of thirteen haplotypes. Interestingly, haplotype segregation in that study did not correspond to host species; in contrast, it corresponded to geographical distribution.
Materials and Methods
Collections
The gonads of two of five examined cobia, Rachycentron canadum were infected with 6 specimens of S. sagittifer in the Persian Gulf off the Iran coast on March 31, 2023. Four specimens were processed for microscopical examination, imaging, and validation of identity. The remaining two specimens were used for molecular studies.
Methods for Microscopical Studies
Four worms (2 males, 2 females) were punctured with a fine needle and subsequently stained in Mayer’s acid carmine, destained in 4% hydrochloric acid in 70% ethanol, dehydrated in ascending concentrations of ethanol (24 hr each), and cleared in 100% xylene then in 50% Canada balsam and 50% xylene (24 hr each). Whole worms were then mounted in Canada balsam. Measurements for diagnostic verification were made in micrometers, unless otherwise noted. Specimens normally deposited in the University of Nebraska’s State Museum’s Harold W. Manter Laboratory (HWML) collection, Lincoln, Nebraska, USA are kept in OMA's collection.
Optical Microscopy
Images created for this presentation were acquired using a Zeiss Axioskop Transmitted Nomarski DIC Phase Contrast Microscope Trinocular (Munich, Germany) and a Canon T3i EOS 600D DSLR Camera (Melville, New York). Measurements are in micrometers unless otherwise noted; the range is followed by the mean values between parentheses when appropriate.
Molecular Procedures
Genomic DNA was extracted from two specimens using a Qiagen DNeasy tissue kit (Qiagen Inc., Valencia, California, USA) according to the manufacturer’s instructions. The partial 18S region was amplified by polymerase chain reaction (PCR) using the primer 18SU467F (forward, 5’-ATCCAAGGAAGGCAGCAGGC-3’); 18SL1310R (reverse, 5’-CTCCACCAACTAAGAACGGC-3’) [20]. The partial 28S region was amplified by PCR using 2 overlapping PCR fragments and the primers for 28S gene were forward, 5’-CTAAGGAGTGTGTAACAACTCACC-3’, reverse, 5’-AATGACGAGGCATTTGGCTACCTT-3’) and forward, 5’-GATCCGTAACTTCGGGAAAA-3’, reverse, 5’-CTTCGCAATGATAGGAAGAGCC-3’ [21]. The partial cox1 region was amplified by PCR using the primer LCO1490 (5’-GGTCAACAAATCATAAAGATATTGG-3’) and HC02198 (5’-TAAACTTCAGGGTGACCAAAAAATCA-3’) [22]. The cycling conditions were as described previously [23]. PCR products were checked on 1.5% agarose gels and purified with Purelink™ Quick Gel Extraction and PCR Purification Combo Kit (Invitrogen, Löhne, Germany). Sequencing from each amplification product was carried out for both directions. Sequences of 28S gene available on the GenBank were less in comparison to 18S and mt cox1 gene; therefore, we excluded the 28S gene from the analysis. 18S and mt cox1 gene were aligned individually using ClustalW and adjusted manually. The DNA sequences obtained in this study were compared by using the BLASTn (nucleotide BLAST) (http://www.ncbi. nlm.nih.gov) with available species data. The nucleotide sequences were deposited in GenBank (https://www.ncbi. nlm.nih.gov/genbank/) (accession numbers PP916512, PP916495, PP916498, PP916505, PQ099320, PQ099319).
Phylogenetic Analyses
Phylogenetic analyses were performed based on the 18S and cox1 sequence data using maximum likelihood (ML) inference with software MEGA v.11 and Bayesian inference (BI) with Topali 2.5 [24, 25], respectively. Echinorhynchus truttae Schrank, 1788 (Polyacanthocephala: Echinorhynchida) was treated as the out-group. The detailed information of Serrasentis species comprised in the present phylogenetic analyses is provided in Table 1. We used jModelTest version 2.1.7 [26] to select a best-fitting substitution model for the 18S and mt cox1 sequences according to the Akaike information criterion. The GTR + G + I model was recognized as ideal nucleotide substitution model. For ML tree reliability was tested using 1000 bootstrap replications. For BI tree, running two independent Markov chain (MCMC) runs of four chains for 10 million generations and sampling tree topologies every 1000 generations. Genetic divergences were calculated for the alignment as uncorrected p-distances using MEGA v.11. S. sagittifer species published sequences of the cox1 gene sequences were used in the phylogenetic analysis for making a median-joining network [27] that inferred for the species relationship amongst haplotypes, estimated by building the haplotype with Network v.10.2.0.0 software version [28].
Results
We currently describe the molecular profile of specimens of S. sagittifer from R. canadum in the Arabian Gulf for the first time and provide diagnostic optical microscopy images for identity verification with the use of our own reference material and the key in Barton, et al. [11].
Amin, et al. [10] provided a listing of reports of juveniles and adults of S. sagittifer from all paratenic and definitive hosts reported from all over the world to date and described their juveniles and adults from 8 species of paratenic hosts and from Rachycentron canadum, respectively, from the same localities in the Arabian Gulf compared to other detailed descriptions of adults from northern Australia, Atlantic North America, and Mexico.
Morphological Validation of Iranian Specimens
A sample of 4 whole-mounted specimens of the Iranian material was processed for optical microscopy and their identity was verified.
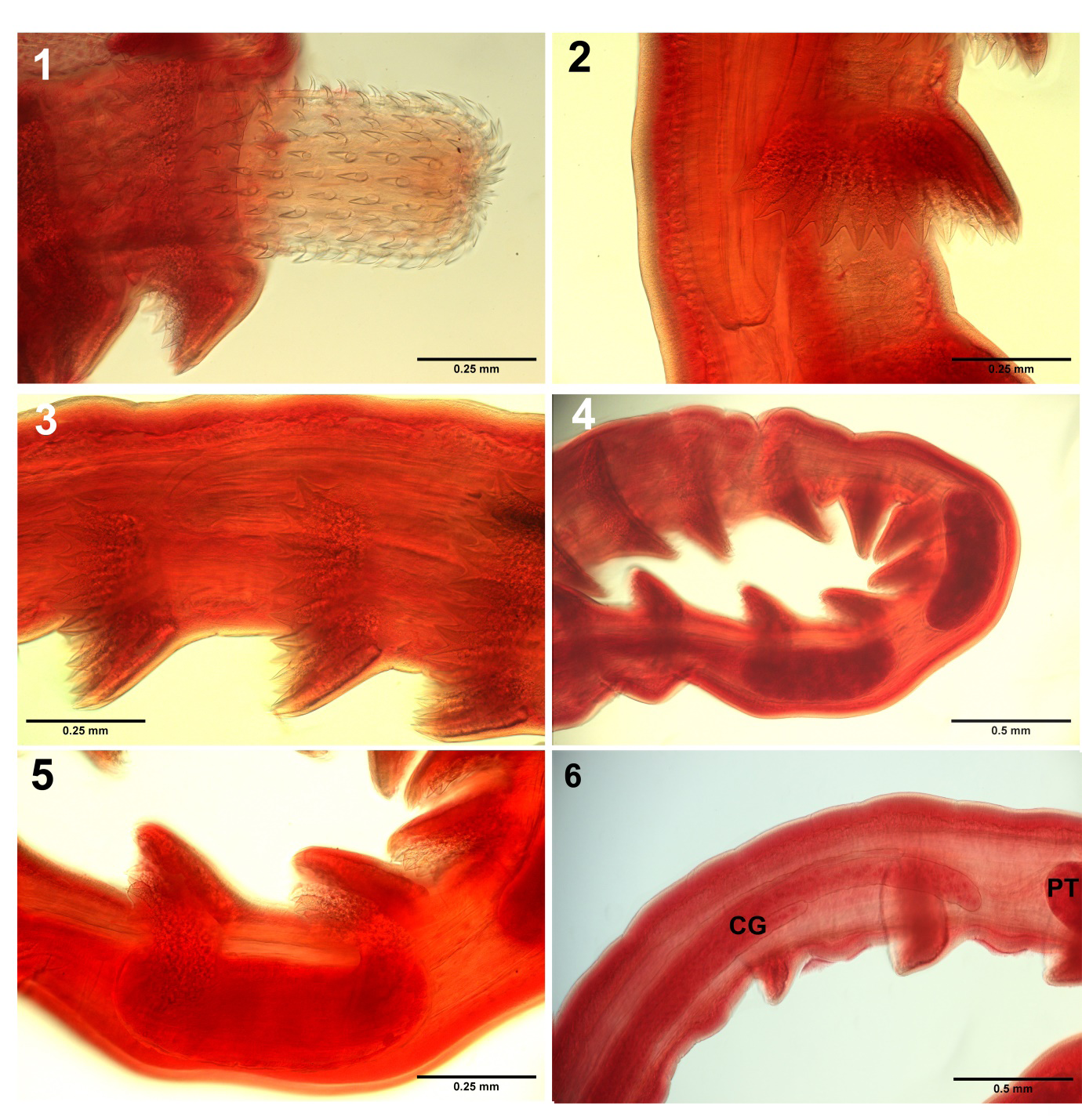
Figures 1-6: Optical microscopy images of specimens of Serrasentis sagittifer from cobia Rachycentron canadum (Linn.) in the Persian Gulf off Iranian waters.
- The proboscis, in part) showing the dorsal slender hooks and the ventral robust ones. Note the simple posteriorly directed roots longer than blades.
- Profile of one anterior spinous comb and the posterior part of the proboscis receptacle.
- Detail of 3 spinous combs showing their granular bases.
- A segment of a male trunk showing the occasional proximity of testes.
- Higher magnification of the posterior part of figs
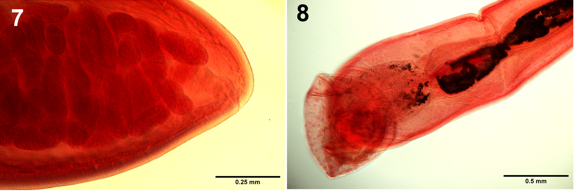
Figures 7,8: Optical microscopy images of reproductive structures of specimens of Serrasentis sagittifer from cobia Rachycentron canadum (Linn.) in the Persian Gulf off Iranian waters. 7. The posterior end of female trunk shows the sub-terminal position of the vagina and gonopore. 8. The posterior parts of a male reproductive system show the terminal gonopore and the bursa.
A few light microscopy images focusing on key diagnostic characters are included for documentation and morphological updates (Figs. 1-8), that do not repeat the SEM images included in Amin, et al. [10]. Some inaccurate aspects of the redescription by Barton, et al. [11] are herein corrected. Our optical microscopy images depict the following features. The slender dorsal proboscis hooks and the robust ventral ones are clearly depicted (Fig. 1). Fig. 2 shows detail of the spines and rays of one transverse latero- ventral spine comb with 10 spines on one side adjacent to the area of the proboscis receptacle depicting thick walls, especially dorsally. Distal end of receptacle thin with double nuclei (first reported here). A series of 3 spinous trunk combs also at the receptacle area demonstrate their granular base (Fig. 3). Fig. 4 shows the occasional proximity of testes to each other in a trunk area studded with a number of spinous combs. A higher magnification of the posterior portion of the specimen is shown in fig. 4 (Fig. 5) depicts detail of the nearby spinous combs and their granular base especially near the posterior testis. The trunk just posterior to the posterior testis (PT) (Fig. 6) shows the non-contiguous cement glands of unequal lengths staggering anteriorly (CG) (first reported here). The posterior end of the female trunk (Fig. 7) shows the sub-terminal position of the vagina and the gonopore. The posterior end of the male reproductive system shows the bursa which may be covered with a large number of single-nucleated cells (Fig. 8) that may be of sensory nature (first noted here). The male gonopore is clearly terminal, not subterminal as indicated by Barton, et al. [11].
Molecular Analysis
Analyses of the two datasets of 18S rDNA and mt cox1 sequences produced almost similar topologies, therefore, described under Results. For each dataset, the ML and BI analyses produced phylogenetic trees by comparably branching topology and congruent support values. These phylogenetic trees portrayed strongly supported clades (including I–III in Fig. 9) within the Rhadinohynchidae.
Two 18S sequences of S. sagittifer obtained herein shows no nucleotide divergence between them. In the genus Serrasentis, 18S sequences are available for S. sagittifer and S. nadakali species. S. sagittifer sequences are available from Australia (MF426933-41, MN705841) and Egypt (KF648554) including the present study specimens (Fig. 9). The phylogenetic reconstruction (Fig. 9) represented clades I including S. nadakali from the host Rachycentron canadum in India. Clade II contained S. sagittifer isolates (MF426933-41, MN705841) harboring different hosts, viz., R. canadum, Lethrinus laticaudis, Lutjanus sebae and L. malabaricus and representative sequences by Australia region clustering within with strong support (Table 1; Fig. 9). The third strongly supported clade (clade III) encompassed exclusively parasites of the Middle East from close zoogeographical areas, i.e. Egypt and Iran. This clade included S. sagittifer isolates from Egypt (from Sparus aurata) and Iran (R. canadum) clustering together with strong support, were genetically close to each other that reflects the geographical region impacts on the parasite phylogeny instead of host herein (Fig. 9). Pairwise comparison of the 18S sequences of isolates of S. sagittifer obtained herein with that of other congeners from Australia showed a range of nucleotide divergence between 1.36 to 1.37%. The 18S sequences of S. sagittifer obtained herein shows a genetic divergence with Egyptian isolate was of 1.24%.
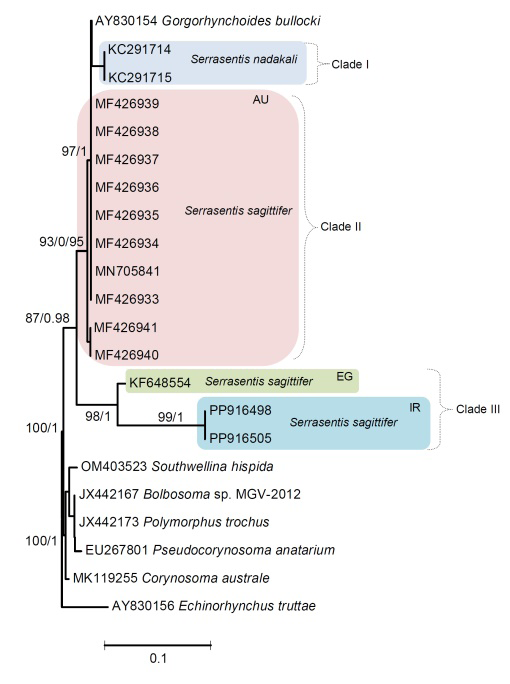
Figure 9: Maximum likelihood (ML) and Bayesian inference (BI) based on the 18S sequence data of acanthocephalan species showing the phylogenetic relationships. Bootstrap support (BS) values ⩾75 in ML tree and Bayesian posterior probabilities (BPP) ⩾ 0.75 in BI tree are shown. Branch length scale bar at lower indicates nucleotide substitutions per site. Biogeographical regions: Australia (AU); Egypt (EG) and Iran (IR).
| Species | Host | Host origin | GenBank accession nos. | References |
|---|---|---|---|---|
| 18S gene | ||||
| Serrasentis sagittifer | Lethrinus laticaudis | Australia | MF426933, 938, 939 | Barton, et al. [11] |
| Lutjanus sebae | Australia | MF426934, 940, 941 | Barton, et al. [11] | |
| Rachycentron canadum | Australia | MF426935, 936 | Barton, et al. [11] | |
| Lutjanus malabaricus | Australia | MF426936 | Barton, et al. [11] | |
| Rachycentron canadum | Australia | MN705841 | Huston, et al. [25] | |
| Sparus aurata | Egypt | KF648554 | Abdel-Gaber, 2013* | |
| Rachycentron canadum | Iran | PP916498, PP916505 | This study | |
| Serrasentis nadakali | Rachycentron canadum | India | KC291714, 715 | Paul, et al. 2013* |
| Mt cox1 gene | ||||
| Serrasentis sagittifer | Lethrinus laticaudis | Australia | MF134302, 303, 304 | Barton, et al. [11] |
| Rachycentron canadum | Australia | MF134301, 297, | Barton, et al. [11] | |
| Lutjanus sebae | Australia | MF134296, 298, 300 | Barton, et al. [11] | |
| Lutjanus malabaricus | Australia | MF134299 | Barton, et al. [11] | |
| Rachycentron canadum | Australia | MN692700, 701, 702 | Huston, et al. [20] | |
| Rachycentron canadum | Iran | PP916512, PP916495 | This study | |
| Serrasentis nadakali | Rachycentron canadum | India | KC291712, 713 | Paul, et al. 2013* |
| Serrasentis gibsoni | Cyclopsetta chittendeni | Mexico | OR826958, 965, 973, 974 | Martínez-Aquino et al. [26] |
| Syacium papillosum | Mexico | OR826975 | Martínez-Aquino et al. [26] |
Table 1: Serrasentis species information used for phylogenetic analysis based on the 18S RRNA and mt cox1 gene sequences. An aste
For the mt cox1 gene, the ML and BI analyses produced trees with congruent topologies. Hence, only the ML tree is presented, using values alongside nodes representing bootstrap support from ML analyses and posterior probabilities from BI, respectively (Fig.10). The two cox1 sequences of S. sagittifer obtained herein, display no nucleotide divergence detected. In the genus Serrasentis, in regard to mt cox1 sequences, three species have their cox1 sequences data available in GenBank, viz., S. sagittifer, S. nadakali and S. gibsoni. The investigated Serrasentis species formed three major well-supported clades. The first clade (I) encompassed all the Australian species of S. sagittifer (MF134296-304, MN692700-702), hosts R. canadum, Lethrinus laticaudis, Lutjanus sebae and L. malabaricus, respectively (table 1; Fig. 10). The second clade (II) encompassed S. sagittifer isolates from the Arabian Gulf (Iran) of the present study parasitizing R. canadum with well supported monophyletic lineage (Fig.10). Similar to the 18S gene, no mt cox1 sequence is available for any Middle Eastern isolate of S. sagittifer available and the phylogenetic analyses strongly support the far phylogenetic proximity of our Arabian Gulf isolate of S. sagittifer from Australian region that also corresponded to the 18S phylogeny results (Fig. 10). The other two well-supported groups, clade III include species of S. nadakali from India and S. gibsoni from Mexico, respectively (Fig. 10). The genetic distance values between Iranian and Australian isolates of S. sagittifer ranged between 1.33 and 1.39%.
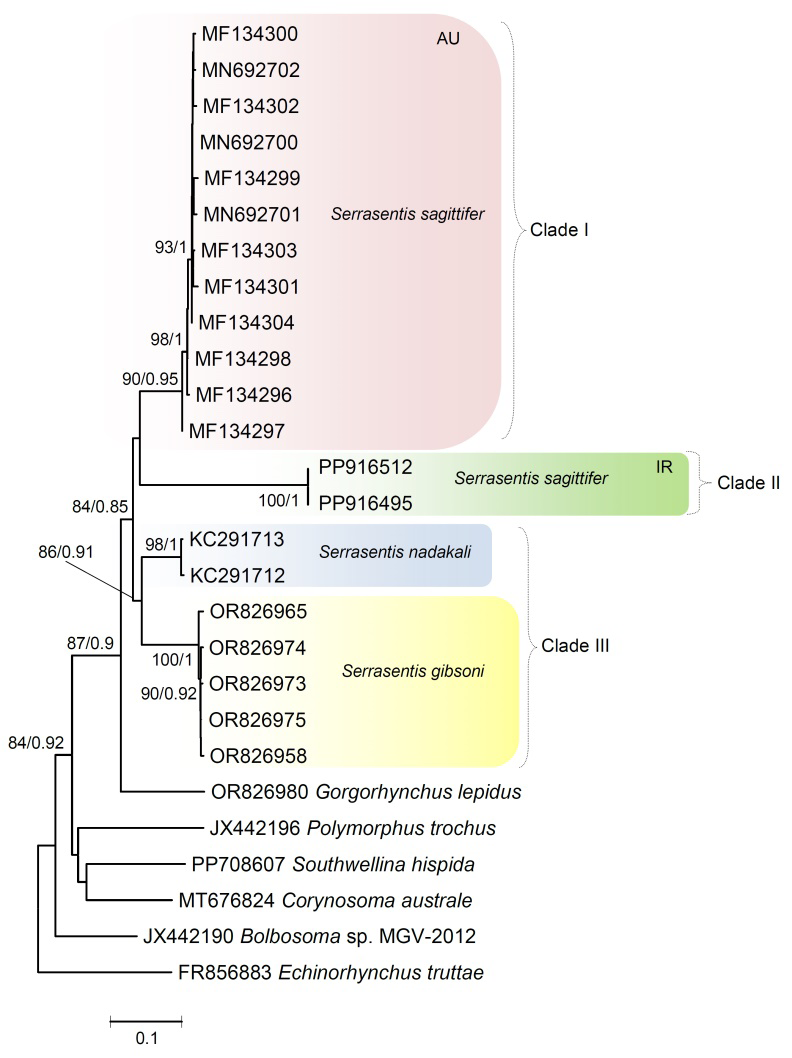
Figure 10: Phylogenetic tree resulting from Maximum likelihood and Bayesian inference analysis of mt cox1 sequence data showing placement of isolates of Serrasentis sagitiffer from the present study including others available on the GenBank database. The accession numbers follow species names. The values above branches represent maximum likelihood bootstrap support values ≥ 75% and Bayesian posterior probability ≥ 0.75, respectively. Branch length scale bar at lower indicates nucleotide substitutions per site. Biogeographical regions: Australia (AU) and Iran (IR).
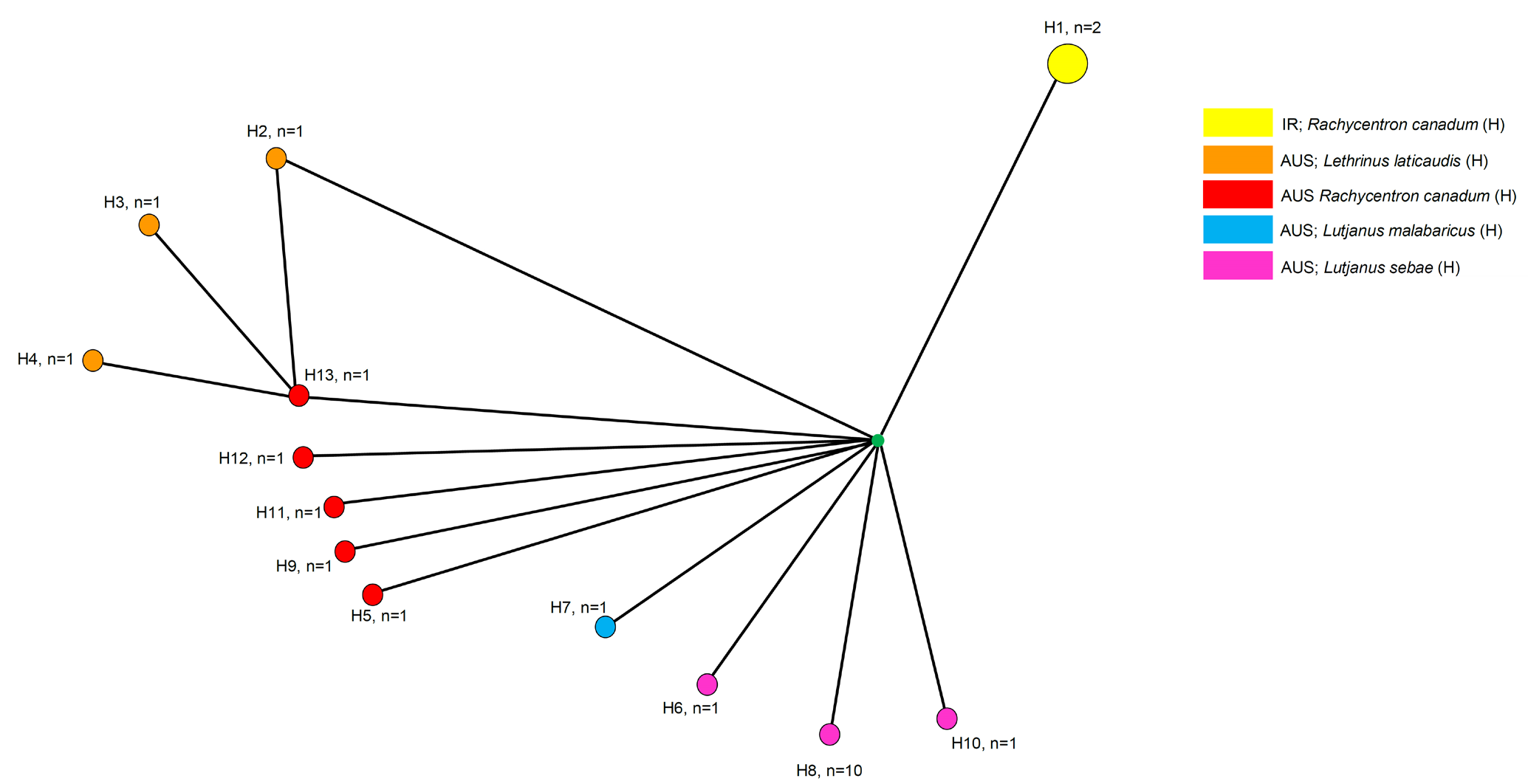
In total, 13 haplotypes among 14 sequences from of S. sagittifer were found and the haplotype diversity (H) was 0.9890 (Fig.11). The haplotype network built for the isolates of S. sagittifer species shows a phylogeographic structure. In fact, of the 13 mtDNA haplotypes detected, H1 shared by the two Arabian Gulf (Iranian) isolates infected the host R. canadum (Fig. 11). The other isolates infecting the same host, R. canadum from Australia, comprises the haplotype H5, H9, H11, H12, H13, respectively (Fig. 11). The other Australian host, L. laticaudis for isolates of S. sagittifer corresponding to H2, H3, H4. An H7 specimen from Australia was found infected with L. malabaricus while H6, H8, H10 shows the host Lutjanus sebae from Australian waters (Fig. 11). The mt cox1 sequence analyses have shown the presence of a number of haplotypes that commonly allow for grouping according to host and/or geography drainages.
Discussion
We have provided additional new morphological features of S. sagittifer not previously published (Figs. 1-8). We have also previously discussed the morphological variability and the world-wide distribution of this acanthocephalan in various hosts, especially cobia, R. canadum which has an overlapping range of distribution with S. sagittifer [10]. Cobia, the only species in the family Rachycentridae, is a migratory pelagic fish that may occur throughout the water column in the tropical and subtropical seas of the world, except in the central and eastern Pacific [31, 32]. Adult cobias are coastal and continental shelf fish, occasionally entering estuaries [33, 34] and living in salinities of 5.0 to 44.5 (euryhaline) [35]. Intestinal damage from S. sagittifer worms was severe in cobia examined by George and Nadakal [36] who obtained infected fish from the Arabian Sea near the Trivandrum coast.
While there was no clear penetration” of the intestine, host physiology and “considerable histopathological alterations in the fish intestine, including hyperplasia of connective tissue cells, metaplasia of epithelial cells [36], and hypertrophy of muscle cells, necrosis, degeneration of the mucus epithelium, and considerable destruction of intestinal villi were observed” (George and Nadakal [36], page 61). Cobia usually feeds near the bottom; however, they may also take prey near the surface [37]. In addition to their presence in the Arabian Gulf (this paper), we have also documented the distribution of this host-parasite system in the West Atlantic coasts of North and South America, east Atlantic off the west coast of West Africa, west Indian Ocean, East Africa, Red Sea, and the Indonesian Islands [10].
Phylogenetic relationships identified for S. sagitiffer may reflect a geographical pattern for the isolates. This specificity pattern is supported by the 18S, mt cox1 phylogeny including haplotype study between the host-parasite/geographical associations for S. sagitiffer. Based on our phylogenetic analyses, S. nadakali and Gorgorhynchoides bullocki is sister to S. sagittifer in the 18S tree, with well supported nodal support and in concordance to the study of Martínez-Aquino, et al. [26]. Several authors suggest paraphyletic assemblage for Rhadinorhynchidea on the basis of the number of cement glands [11, 21, 25, 38, 39, 40, 41]. Later, Pichelin, et al. [42] remarked that for the identification of acanthocephalan families, notably for the Rhadinorhynchidae, the number of cement glands is a distinctive character, although, few authors are do not agree [43, 44].
In this study, it is attention-grabbing to notice that the species of Serrasentis and Gorgorhynchus (both with four cement glands) form a separate clade in comparison to Gorgorhynchoides spp. i.e., G. bullocki (with six cement glands). Here, we think that may be the cement glands number shown as a synapomorphy at the level of genera and families. Specimens of S. sagittifer seem to be highly host specific for the cobia, R. canadum, irrespective of the geographical region. The widespread distribution of S. sagittifer across a range of different geographical areas/habitats may contribute to reducing their genetic divergence among populations. The present study confirms that S. sagittifer is a widespread parasite as also reported by Barton, et al. [45] and Martínez- Aquino, et al. [26]. The haplotype network genealogy produced in this study based on mt cox1 sequences from S. sagitiffer show a phylogeographic structure; therefore, the haplotypes grouped into their own geographic clusters. Despite dealing with the same species in large geographic distances among the populations, we demonstrate that the populations were genetically discriminated from each other.
Conclusions
Our light microscopy images focusing on key diagnostic characters document morphological characters of specimens of S. sagittifer and update features not previously documented or erroneously reported. For example, we correct inaccurate aspects of the redescription by Barton, et al. [11] and detail additional aspects of the acanthocephalan morphology. Phylogenetic relationships identified for S. sagitiffer reflect a geographical pattern for the isolates for the first time. This specificity pattern is supported by the 18S, mt cox1 phylogeny including haplotype study between the host- parasite/geographical associations for S. sagitiffer. Future morphological studies with molecular phylogenetic trees including more species of the family Rhadinorhynchidae and Isthmosacanthidae are required so that we would be able to approve the phylogenetic relationships identified herein in Isthmosacanthidae, and elucidate the progression of their morphological characters like the number of cement glands.
Acknowledgements
This project was supported by an Institutional Grant from the Parasitology Center, Inc. (PCI), Scottsdale, Arizona. We are very grateful to our two Iranian colleagues, Dr. Zahra Gharibi who collected and dissected fish samples and was a link between Qazvin and Hormozgan Universities, and to Dr. Majid Pirestani whose administrative connections with Fisheries and overall support are gratefully appreciated. The authors thank the Department of Zoology, Chaudhary Charan Singh University, Meerut, India for providing laboratory facilities.
Authors Contributions
Amin identified the acanthocephalan S. sagitiffer, did the research, wrote the manuscript except for the molecular part, and created the optical microscope images. Chaudhary performed the molecular analysis including the phylogenetic trees and tables. Milad only helped with fish dissection, administrative tasks, and shipping and Eslahi only obtained and dissected fish and managed the parasite collection and communication with the senior author. Singh provided the molecular lab facility.
Compliance with Ethical Standards
- Conflict of Interest: The authors declare conflicts of interest none.
- Ethical Approval: The authors declare that they have observed all applicable ethical standards.
References
-
Monticelli FS (1905) Su di un Echinorhynco della collezione del Museo Zooligico di NapoIi (_Echinorhynchus_ _rhytidodes_ Monticelli). Annuario del Museo Zoologico della Reale Universita di Napoli 2 n.s. Nuova Serie No 251: 1-11.
-
Porta A (1907) Contributo allo studio degli acantocefali dei pesci. Biologica Torino 1: 377-423.
-
Van Cleave HJ (1918) Acanthocephala of the subfamily Rhadinorhynchinae from American fish. J Parasitol 5: 17-24.
-
Van Cleave HJ (1924) A critical study of the Acanthocephala described and identified by Joseph Leidy. Proc Acad Nat Sci Philadelphia 76: 279-332.
-
Van Cleave HJ, Lincicome DR (1940) A reconsideration of the acanthocephalan family Rhadinorhynchidae. J Parasitol 26: 75-81.
-
Noronha D, de Fabio SP, Pinto RM (1978) _Golvanorhynchus_ _golvani_ n. g., n. sp. (Palaeacanthocephala, Allorhadinorhynchinae). Atas Soc Biol Rio de Janeiro 19: 63-64.
-
Cable RM, Linderoth J (1963) Taxonomy of some Acanthocephala from marine fishes with reference to species from Curaçao, N. A., and Jamaica, W. I. J Parasitol 49: 706-716.
-
Smales LR (2012) A new acanthocephalan family, the Isthmosacanthidae (Acanthocephala: Echinorhynchida), with the description of _Isthmosacanthus fitzroyensis_ n. g., n. sp. from threadfin fishes (Polynemidae) of northern Australia. Syst Parasitol 82: 105-111.
-
Huston DC, Smales LR (2021) _Gorgorhynchoides_ _pseudocarangis_ n. sp. (Acanthocephala: Isthmosacanthidae) from _Pseudocaranx_ _dentex_ (Carangidae) in southeast Queensland, Australia, with comments on the Isthmosacanthidae. Syst Parasitol 98: 399-412.
-
Amin OM, Heckmann RA (2021) New perspectives on the distribution, morphology and hook chemistry of _Serrasentis sagittifer_ (Linton, 1889) Linton, 1932 using SEM and Energy Dispersive X-ray Analysis. Sci Parasitol 22: 88-111.
-
Barton DP, Smales LR, Morgan JAT (2018) A redescription of _Serrasentis_ _sagittifer_ (Rhadinorhynchidae: Serrasentinae) from _Racycentron_ _canadum_ with comments on its biology and its relationship to other species of _Serrasentis_. J Parasitol 104(2): 117-132.
-
Golvan YJ (1956) A new species and variety of _Acanthocephalus_ parasitic of marine fish from the coasts of Senegal and redescription of _Serrasentis socialis_ (Leidy 1851) Van Cleave 1924. Ann Parasitol Hum Compar 31: 225-239.
-
Golvan YJ (1961) Le Phylum des Acanthocephala. Troisième note. La Classe des Palaeacanthocephala (Meyer, 1931). Ann Parasitol Hum Comp 35: 76-91.
-
Golvan YI, Houin R (1964) Revision of Palaeacanthocephala. Second note. The Van Cleave and Lincicome family of Gorgorhynchidae, 1940. Ann Parasitol Hum Comp 39: 535-605.
-
Salgado-Maldonado G (1978) Acantocephala of fishes V. Redescription of four species of Palaeacantocephala parasitic of fishes of Mexico. An Inst Biol Univ Nacion Aut Mex Ser Zool 49: 49-69.
-
Fonseca MCG, Knoff M, Felizardo NN, Lopes Torres EJ, Nogueira Di Azevedo MI, et al. (2019) Acanthocephalan parasites of the flounder species _Paralichthys isosceles_, _Paralichthys patagonicus_ and _Xystreurys rasile_ from Brazil. Braz J Vet Parasitol 28: 346-359.
-
Golvan YJ (1969) Systematics of Acanthocephala (Acanthocephala Ruldolphi, 1801). The order of Palaeacanthocephala Meyer, 1931. The superfamily of Echinorhynchoidea (Cobbold, 1876) Golvan et Houin 1963. Mem Mus Natl Hist Ser A Zoologie pp: 373.
-
Travassos L (1966) _Serrasentis sagittifer_ (Linton, 1889) (Acantocephala). Mem Inst Osw Cruz 64: 1-10.
-
Abdel-Ghaffar F, Morsy K, Abdel-Gaber R, Mehlhorn H, Al Quraishy S, et al. (2014) Prevalence, morphology, and molecular analysis of _Serrasentis_ _sagittifer_ (Acanthocephala: Palaeacanthocephala: Rhadinorhynchidae), a parasite of the gilthead sea bream _Sparus aurata_ (Sparidae). Parasitol Res 113: 2445-2454.
-
Suzuki N, Hoshino K, Murakami K, Takeyama H, Chow S (2008) Molecular diet analysis of phyllosoma larvae of the Japanese spiny lobster _Panulirus japonicus_ (Decapoda: Crustacea). Mar Biotechnol (New York), 10: 49-55.
-
García-Varela M, Nadler SA (2005) Phylogenetic relationships of Palaeacanthocephala (Acanthocephala) inferred from SSU and LSU rDNA gene sequences. J Parasitol 91: 1401-1409.
-
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol 3: 294-299.
-
Amin OM, Chaudhary A, Singh HS (2023) Redescription of _Illiosentis cetratus_ Van Cleave, 1945 (Acanthocephala: Illiosentidae) from _Menticirrhus undulatus_ (Girard) in California, with notes on _Illiosentis furcatus_ from Peru. Folia Parasitol: 70.
-
Milne I, Lindner D, Bayer M, Husmeier D, Mc Guire G, et al. (2008) TOPALi v2: a rich graphical interface for evolutionary analyses of multiple alignments on HPC clusters and multi-core desktops. Bioinformatics 25: 126-127.
-
Tamura K, Stecher G, Kumar S (2021) MEGA11: Molecular Evolutionary Genetics 518 Analysis Version 11. Mol Biol Evol 38: 3022-3027.
-
Huston DC, Cribb TH, Smales LR (2020) Molecular characterisation of acanthocephalans from Australian marine teleosts: proposal of a new family, synonymy of another and transfer of taxa between orders. Syst Parasitol 97: 1-23.
-
Martínez-Aquino A, García-Teh JG, Ceccarelli FS, Aguirre- Macedo ML, Vidal-Martínez VM (2023) Integrative taxonomy of _Serrasentis gibsoni_ n. sp. (Acanthocephala: Isthmosacanthidae) from flatfishes in the Gulf of Mexico. J Helminthol: 97.
-
Darriba D, Taboada GL, Doallo R, Posada D (2012) jModelTest 2: more models, new heuristics and parallel computing. Nature Methods 9: 772.
-
Bandelt HJ, Forster P, Rohl A (1999) Median-joining networks for inferring intraspecific phylogenies. Mol Biol Evol 16: 37-48.
-
Bandelt HJ, Dress AWM (1992) A canonical decomposition theory for metrics on a finite set. Adv Math 92: 47-105.
-
Shaffer RV, Nakamura EL (1989) Synopsis of biological data on the cobia _Rachecentron canadum_ (Pisces: Rhachycentridae). NOAA Technical Report NMFS 82, FAO Fisheries Syn 153: 21.
-
Ditty JG, Shaw RF (1992) Larval development, distribution, and ecology of cobia _Rachycentron canadum_ (Family: Rachycentridae) in the northern Gulf of Mexico. Fish Bull 90: 668-677.
-
Collette BB (1978) Rachycentridae. In: Fischer W (Ed.), FAO species identification sheets for fishery purposes, western central Atlantic (Fishing area 31), FAO, Rome, Italy 4.
-
Robins CR, Ray GC (1986) Field guide to Atlantic coast fishes of North America. Houghton Mifflin Co., Boston, MA, pp: 354.
-
Resley MJ, Webb K, Holt GJ (2006) Growth and survival of juvenile cobia _Rachycentron canadum_, at different salinities in a recirculating aquaculture system. Aquaculture 253: 398-407.
-
George PV, Nadakal AM (1981) Observations on the intestinal pathology of the marine fish, _Rachycentron_ _canadus_ (Giinther) infected with the acanthocephalid worm, _Serrasentis nadakali_ (George and Nadakal, 1978). Hydrobiol 78: 59-62.
-
Knapp FT (1951) Food habits of the sergeantfish, _Rachycentron canadum_ Copeia 1: 101-102.
-
Verweyen L, Klimpel S, Palm HW (2011) Molecular phylogeny of the Acanthocephala (class Palaeacanthocephala) with a paraphyletic assemblage of the orders Polymorphida and Echinorhynchida. PLoS One 6: e28285.
-
Braicovich PE, Lanfranchi AL, Farber MD, Marvaldi A, Luque JL, et al. (2014) Genetic and morphological evidence reveals the existence of a new family, genus and species of Echinorhynchida (Acanthocephala). Folia Parasitologica 61: 377-384.
-
Lisitsyna OI, Kudlai O, Cribb TH, Smit NJ (2019a) Three new species of acanthocephalans (Palaeacanthocephala) from marine fishes collected off the East Coast of South Africa. Folia Parasitol 66: 12.
-
Sharifdini M, Amin OM, Heckmann RA (2020) The molecular profile of _Paratrajectura longcementglandatus_ Amin, Heckmann et Ali, 2018 (Acanthocephala: Transvenidae) from percid fishes in the marine waters of Iran and Iraq. Helminthologia 57: 1-11.
-
Pichelin, S, Cribb, TH (2001) The status of the Diplosentidae (Acanthocephala: Palaeacanthocephala) and a new family of acanthocephalans from Australian wrasses (Pisces: Labridae). Folia Parasitol 48: 289-303.
-
Amin OM, Heckmann RA, Ha NV (2011) Description of two new species of _Rhadinorhynchus_ (Acanthocephala, Rhadinorhynchidae) from marine fish in Halong Bay, Vietnam, with a key to species. Acta Parasitol 56: 67-77.
-
Amin O (2013) Classification of the Acanthocephala. Folia Parasitol 60: 273-305.
-
Barton DP, Smales LR (2015) Acanthocephalan cystacanths from flatfish (Order Pleuronectiformes) in Tropical Australian waters. J Parasitol 101: 429-435.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework