Review of the Taxonomic Status of the Chelid Turtle Parahydraspis paranensis Wieland 1923 from Late Miocene at Parana, Entre Rios Province, Argentina: with Comments on the Paleoenvironmental Settings from the lower Levels of Ituzaingó Formation
The panchelid turtles (stem chelid + crown chelid), one of the pleurodiran clades, display a disjunct distribution in Australasia and South America. Panchelids appeared in the fossil record during South America's Early Cretaceous (Aptian-Albian) and Late Cretaceous (Cenomanian) in Australasia. Among the crown Chelidae, the Chelinae include genera such as Acanthochelys, Mesoclemmys, Platemys, Phrynops, Ranacephala, and Rhinemys characterized by a restricted, poor or null fossil record in South America. Among this chelid clade, Parahydraspis paranenesis is a carapace-based species described by Wieland in 1923 from upper Miocene Ituzaingó Formation outcrops along the Parana river cliffs, Parana city, Entre Rios Province, Argentina. Wieland referred to this new species as a different genus from the extant Hydraspis (an old synonymous genus of the genus Phrynops), depending upon a misinterpretation of the plastral bridge. Although a recent phylogenetic analysis nested this Miocene species with stem chelid species, in this contribution based on anatomical evidence on the holotype and referred specimens, we consider Parahydraspis paranensis as a fully valid species of the Phrynops genus. The continental terrestrial and aquatic fauna of the "Conglomerado osifero" lower levels of the Ituzaingó Formation where the holotype and specimens referred to as “Parahydraspis” paranensis were recovered, indicate a warmer climate and suggest the presence of forested areas along river’s shores where these levels were deposited.
Introduction
The Testudinata (stem turtles + Testudines) appeared in the fossil record during the Late Triassic (Norian) in Pangea [1, 2, 3, 4, 5, 6, 7, 8, 9, 10]. Testudines rapidly expanded in geographic range and ecological diversity during the Mesozoic [11, 12]. The speed with which this diversity built up suggests that turtles may have undergone rapid, early adaptative radiation [13].
However, this early diversification was not sustained, and living turtle diversity is low. Consequently, crown turtles are a relatively species-poor clade of amniotes, with currently 357 recognized living species and 486 taxa relative to other, much younger crown amniote clades such as birds (>10,000 species), mammals (>5,000 species), and squamate reptiles (>10,000 species) [14, 15, 16, 17, 18].
According to [19] diversity, time, and diversification analyses indicate that the two major lineages within the clade Cryptodira and Pleurodira, diverged in the Late Triassic, followed by the Early Jurassic divergence between the softshell Trionychia (Carettochelyidae and Trionychidae) and the remaining cryptodires. Most recognized families diverged in the Late Cretaceous or Early Paleogene [20, 21, 22, 23, 24].
A specious comparison between these two different clades Cryptodira (hidden-necked turtles) and Pleurodira (side-necked turtles), with extant representatives indicates that the former (261 spp., 379 taxa) is a more specious clade than the latter [14]. Among extant pleurodiran turtles (96 spp, and 107 taxa), crown Chelidae displays the greater species richness (represented by 61 spp., and 67 taxa), in comparison with Pelomedusoidea (35 spp., and 40 taxa) [14].
Extinct and extant panchelid (stem + crown) species are aquatic and semiaquatic turtles and display a disjunct distribution across Australasia and South America [14, 25, 26]. It has 61 extant spp., 67 taxa, and about 26 valid extinct species are known [14, 26, 27, 28]. The Cretaceous and Paleogene stem chelids are well recorded in Patagonia but are scarcer in Australia [26, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44]. In contrast, the Neogene record of valid species is more specious in Australia than in South America [26, 29, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55].
According to [14] among the crown Chelidae, the Chelinae include genera such as Acanthochelys, Chelus, Mesoclemmys, Phrynops, Platemys, Ranacephala and Rhinemys [56, 57, 58, 59].
Among the Chelinae clade, Acanthochelys and Phrynops are characterized by a restricted and poor fossil record in southern South America. Acanthochelys includes four extant species (A. macrocephala, A. pallidipectoris, A. spixii and A. radiolata, but the only extinct species is A. cosquinensis from the Pliocene of Cordoba province (Argentina) [48, 60, 61, 62, 63]. In addition, the genus Phrynops sensu stricto (Phrynops geoffroanus complex [64] includes three or four extant species (P. geoffroanus [65], P. hilarii [62] P. tuberosus [66], and P. williamsis [64], after [14], depending on whether the authors [67, 68] considering P. tuberosus as synonymous or as a subspecies of P. geoffroanus respectively or recognized it as a whole valid species at first [66, 14, 59, 60, 69, 70, 71].
Among the Chelinae clade Parhydraspis paranensis is a carapace-based species described by Wieland in 1923 from upper Miocene Ituzaingó Formation outcrops along the Parana river cliffs, Parana city, Entre Rios Province, Argentina. At this time, Wieland referred to this new species as a different genus from the extant Hydraspis (an old synonym of genus Phrynops), depending on its particular interpretation of the plastral bridge, which is absent in the holotype (Figures 1A-1C).
![Figure 1: Parahydraspis paranensis holotype (YPM 1565). Photographs and drawings (A) dorsal and (B), visceral views of the carapace, (C) Wieland’s interpretation of plastral bridge redrawing from Wieland [46].](/fulltextimages/13603/fig_1.png)
Although a recent phylogenetic analysis [28] nested this Miocene species with stem chelid species, most of the current turtle synthesis papers suggest the assignation of Parahydraspis pararanensis to Phrynops sensu stricto or its close relationships with extant species of Phrynops geoffroanus complex [48], other authors have proposed its specific validation [26, 72]. In this contribution based on anatomical evidence of the holotype and referred specimens, we aim to propose Parahydraspis paranensis as a fully valid species of the Phrynops genus.
Materials and Methods
Materials
The holotype of “Parahydraspis” paranensis in the study proceeds from “Conglomerado osifero” Ituzaingó Formation (Entre Rios province, Argentina), and it is housed at the Paleontological Vertebrates Collection of the Yale Peabody Museum (YPM 1565), New Haven, USA. The refereed material to “Parahydraspis” paranensis belongs to the collection of the Division Paleontologia Vertebrados, Museo de La Plata (MLP), Argentina, under the collection numbers: MLP 41-XII-13-782; MLP-41-XII-13-793; MLP-41-XII-13-805; MLP-62-XII-10-9; MLP-79-VIII-12-18; MLP-79-VIII-21-2; MLP-79-VIII-21-5; MLP-79-VIII-21-6; MLP-79-VIII-21-7; MLP-79-VIII-21-8; MLP-79- VIII-21-9; MLP-79-VIII-21-10; MLP-79-VIII-21-1; MLP-79-VIII-21-12; MLP-79-VIII-21-13; MLP-79- VIII-21-14; MLP-79-VIII-21-16; MLP-79-VIII-21-17; MLP-79-VIII-21-18; MLP-79-VIII-21-19; MLP-79- VIII-21-20; MLP-80-IV-9-1 (see below).
The anatomy of the carapace is based on the terminology of [73]. Chronological issues follow the International Chronostratigraphic Chart [74, 75].
We used specimens of different extant species of Phrynops sensu stricto (e.g., P. geoffroanus, P. hilarii and P. williamsi) and Mesoclemmys (e.g., M. vanderhaegei, M. gibba) belongs to different institutions for comparisons with the fossil material examined (see institutional abbreviations, description, and discussion). Anatomical Abbreviations: ABSS: Anterior Bridge Strut Scar, C: Cervical Scute; CO: Costal Bone; M: Marginal Scute; Ne: Neural Bones; Nu: Nuchal Bone; PE: Peripheral Bone; P: Pleural Scute; PBSS: Posterior Bridge Strut Scar; R: Rib/ Gomphosis; V: Vertebral Scute. Institutional Abbreviations: AMNH: American Museum of Natural History, New York, NY, USA; FCDPV: Coleccion de Paleontologia Vertebrados, Facultad de Ciencias, Universidad de la Republica, Uruguay; FMNH: Field Museum of Natural History, Chicago, USA; MACN-H, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia,” Herpetological, collection, Ciudad Autonoma de Buenos Aires, Argentina; MCT, Museu de Cie^ ncias da Terra, Serviço Geologico do Brasil, Rio de Janeiro, Brazil; MHNSR-Z, Museo de Historia Natural San Rafael, Zoological collection, San Rafael, Mendoza; Argentina; MNHN, Museum National dHistoire Naturelle de Paris, France; MLP, Museo de La Plata, Vertebral Paleontological collection, La Plata, Argentina; MZUC, Museo de Zoologia, Facultad de Ciencias Exactas, Fisicas y Naturales, Universidad Nacional de Cordoba, Argentina; MZUSP, Museu de Zoologia Universidade de Sao Paulo, Sao Paulo, Brazil; NHMUK, Natural History Museum, London, UK.; USNM, National Museum of Natural History, Smithsonian Institution, Washington DC, USA; YPM, Yale Peabody Museum of Natural History, Yale University, New Haven, USA.
Results
Systematic Paleontology
Testudinata [76], sensu [77], Testudines [78], sensu [79], Pleurodira [80], sensu [81], Pan-Chelidae [82], Chelidae [83], sensu [84], Chelinae [85], Phrynops [58], Phrynops paranensis [46].
Synonymy
Parahydraspis paranensis [46, 86, 87], Phrynops geoffroanus [88], Phrynops complex geoffroanus [89, 48], Phrynops s.l. [Phrynops s.s.] sp. [29], Phrynops cf Phrynops geoffroanus complex [90, 91], Phrynops paranensis [14, 26, 72].
Holotype
YPM 1565, most of the carapace except the caudal portion [46] (Figure 1).
Refereed Material
MLP 41-XII-13-782, medial fragment of the left costal 1; MLP-41-XII-13-793, left hypoplastron; MLP-41-XII-13-805, right hyoplastron; MLP-62-XII-10-9, plastral fragment; MLP- 79-VIII-12-18, anterior left portion of the carapace (left nuchal portion, left peripherals 1-4 and left costal 1), visceral carapace surface with an anterior bridge strut scar (= axillary buttress) that extends to the posterior margin of costal one, with a central constriction and a medial expansion, and an acute angle between rib/gomphosis and anterior bridge strut scar about 15 degree (a diagnostic condition of P. paranensis as listed below); MLP-79-VIII-21-2, proximal end of the left humerus, fragment of the right hyoplastron; MLP-79-VIII-21-5, left hyoplastron; MLP-79-VIII-21-6, two medial fragments of the costal bones, two fragments of the right epiplastra, four fragments of right and two left hypoplastra, one left hypoplastron, one fragment of right hyo plus xiphyplastron, one fragment of left xiphyplastron; MLP- 79-VIII-21-7, left peripherical bone 8, left peripherical bone 3; MLP-79-VIII-21-8, right peripherical bone 8, a fragment of left costal 1 and a right fragment of costal 8; MLP-79- VIII-21-9, three neural bones; MLP-79-VIII-21-10, left costal 1, two right costal bones; MLP-79-VIII-21-11, four peripheral bones of the posterior carapace, left peripheral 2; MLP- 79- VIII-21-12, suprapygal; MLP-79-VIII-21-13, distal end of left xiphyplastron; MLP-79-VIII-21-14, entoplastron; MLP-79- VIII-21-16, left peripheral bones 3-4 and a fragment of the right costal 1; MLP-79-VIII-21-17, lateral extreme of the left costal 8 and a neural bone; MLP-79-VIII-21-18, a left femur, proximal extreme of right humerus, five left pelvic halves, two left ilium, four right ilium, eleven neurals, one lateral fragment of the right costal 1 with an attached axillar pillar of the plastral bridge, a fragment of another costal 1, and one left hypoplastron; MLP-79-VIII-21-19, two right hyoplastra, left peripheral 9; MLP-79-VIII-21-20, left hyoplastron, a costal bone, and a portion of the carapace; MLP-80-IV-9-1, fragment of the carapace.
Stratigraphic and Geographic Provenance
Lower levels “Conglomerado osifero” from Ituzaingó Formation (late Miocene), see geographic location map and generalized stratigraphic section of the late Cenozoic strata at the left side cliff along the Parana river near the city of Parana, Entre Rios, Argentina [91] (Figures 1 & 2).
Enmended Diagnosis
Parahydraspis paranensis can be diagnosed as a chelid pleurodiran turtle due to the presence of a cervical scute on the cranial margin of the carapace nuchal bone and the loose attachment between carapace and plastron. It can be attributed as a representative of Phrynops sensu stricto by the presence of a continuous neural series and a likely nuchal neural 1 contact. It differs from the remaining Phrynops spp. such as P. geoffroanus, P. tuberosus, P. hilarii, and P. williamsi by the presence of a shorter vertebral scute I, a wider cervical scute, and an anterior bridge strut scar (= axillary buttress) that extends to the posterior margin of costal 1, with a central constriction and a medial expansion, and a low acute angle (about 15 degrees) between rib/gomphosis and the strut anterior bridge scar. The posterior bridge strut scar (= inguinal buttress) extends to the most lateral end of costal bones 5 and 6 as other Phrynops spp.; but in contrast, reaches the medial ends of the peripheral bones 8-9.
Redescription of Holotype of Parahydraspis paranensis
The carapace in YPM 1565 (Figure 1A) is broadly oval and low-domed as the extant specimens assigned to Phrynops sensu stricto such as Phrynops geoffroanus (AMNH 79048, MZUSP 2635) (Figure 2), P. hilarii (FMNH 22481, MHNSR- Z- 1550, NHMUK 86-3-10-1) (Figure 3), or P. williamsi (MACN H-38395, USNM 306646) (Figure 4).
The odd bones series in YPM 1565 starts with a, heptagonal nuchal bone (Figures 1A & 1B). This is in contrast to a typical hexagonal nuchal present in chelid species. The caudal margin width of the nuchal is more than twice the cranial margin. The nuchal is followed by a series of six neural bones in YPM 3536 (Figure 1A). Although the area where the neural 1 is positioned is damaged, the nuchal bone-neural 1 contacts can only be suggested.



Although the typical arrange of a continuous series with nuchal-neural 1 contacts are diagnostic characters of Phrynops sensu stricto, (Figure 5A) as shown in a specimen (NHMUK 84-2-5-1) assigned to Phrynops williamsi [64] (Figure 6). An intraspecific variation is seen in MACN 35395 referred to P. williamsi shown [92], where the medial portions of costal bone 1 make contact, preventing contact between the nuchal and neural 1 (Figure 5B). Although the anterior portion of neural 2 is not recognizable in YPM 1565, the neural bones 3 to 6 are hexagonal with short anterolateral margins, displaying a coffin outline appearance. As is typical in turtles, YPM 1565, the right and left costal bones 1-6 are positioned on both sides of the carapace, with at least 8 pairs of peripheral bones on both sides and only a peripheral 9 on the right. As is typical in chelids, costal 1 is twice longer than the costal 2 and trapezoidal. The remaining costal bones (2-6) are subrectangular and similar in size. Anterior free peripheral bones 1 -2 and the lateral portion of peripheral 3 seem slightly shorter than the posterior ones. The peripheral bones 1-3 on both sides are trapezoidal. 4-6 are subrectangular, whereas 7-8, and 9 on the right side are trapezoidal.
![Figure 5: Schematic drawing of intraspecific variation in nuchal-neural 1 contact in different specimens of Phrynops williamsi (NHMUK 84-2-5-1) redrawing from Rhodin, et al. [64] (Figure 6) and (MACN 35395) redrawing from Cabrera [92] (Figure 4).](/fulltextimages/13603/fig_5.png)
On the visceral surface, YPM 1565 (Figure 1B), the nuchal bone shows a hexagonal outline in contrast to the heptagonal dorsal shape. The cranial margin of this bone shows the impression of the cervical and a small portion of the marginal scutes 1; the impression of the remaining marginal scutes extends to the anterior peripheral bones. Although the two anterior neural bones are not easily distinguished, they seem to be present, and the posterior portion of the first thoracic vertebra and the posterior part of the second one are preserved. The remaining visceral neural surfaces (3- 6) show scars on the upper portion of the neural arch. The anterior bridge strut scar (= axillary buttress) extends from peripherals 3 and 4 to most of the posterior margin of coastal 1, almost in contact with the suture with costal 2 on the right side and nearly close to this sutural contact on the left one, with a central constriction and a medial end expansion.
Besides, YPM 1565 shows a 15-degree angle between rib/gomphosis and strut anterior bridge structure scar. In contrast, a subparallel margin of anterior bridge strut scar in P. geoffroanus (AMNH 79048, MZUSP 2635), P. hilarii (FMNH 22481, MHNSR-Z- 1550), and P. williamsi (MACN H-38395, USNM 306646) or some extinct Phrynops spp., as some specimens MCT (ex DGM) 302 R, 1472-R, are referred as superspecies (Phrynops) (Phrynops) (geoffroanus) spp. or specimen PVL 3351 from El Morterito Formation, Valle del Cajon (Catamarca province) (Figures 1 & 2) [8, 10, 12, 48, 93]. The posterior bridge strut scar (= inguinal buttress) in the holotype (YPM 1565) of P. paranensis (Figure 1B) extends on most of the lateral third extremes of posterior costal 5 and anterior costal 6 as other Phrynops spp. such as P. hilarii (FMNH 22481, MHNSR-Z-1550) and P. williamsi (MACN-H- 38395, USNM 306646, see Figures 2B & 3B), but only on costal 5 and peripheral 7-8 in P. geoffroanus (AMNH 79048, MZUSP 2635, see Figure 4B); but in contrast to them, the posterior bridge strut scar reaches in P. paranensis (YPM 1565) the medial ends of the peripheral bones 8-9.
The cervical scute is slightly longer than wide and covers most of the cranial part of the nuchal bone in YPM 1565 (Figure 1A). In contrast, extant P. geoffroanus (AMNH 79048, MZUSP 2635), P. hilarii (FMNH 22481, MHNSR-Z- 1550), P. williamsi (MACN-H- 35395) show a cervical scute longer than wide (Figures 2A,3A & 4A), as is also in some specimens of extinct species such as PVL 3351 from El Morterito Formation, Valle del Cajon (Catamarca province Argentina) (48, plate 1 Figures 1 & 2), whereas the specimen MCT (ex DGM) 301 R shows a shows a cervical scute significantly wider than long as was illustrated by Kischlat [93], plates 8, from late Oligocene-early Miocene, Tremembe Formation, Tabuate Basin, Sao Paulo, Brazil.
In YPM 1565 (Figure 1A), the intervertebral sulcus V1- V2 seems to cross the caudal portion of neural 1. In contrast, the intervertebral sulcus V2-V3 crosses the neural 3 and the intervertebral sulcus V3-V4 crosses the caudal portion of the neural 5.
The vertebral scute 1 is more expansive than long and shorter than the remaining vertebral scutes 2-3 and probably vertebral 4; a similar pattern is seen in Phrynops sp. (PVL
3351) from El Morterito Formation Late Miocene and in the identical specimens of P. williamsi (MACN-H-35395) or P. geoffroanus (AMNH 79048), but not in other specimens of the same species (USNM 65116) or in P. hilarii (MHNSR-Z-1550, MZUC III. 1212) the latter as illustrated by Cabrera [92]. In YPM 1565, vertebral 1 covers the nuchal, a slight portion of caudal of peripheral 1, a possible anterior portion of neural 1 if present, and a medial part of both costal 1. The vertebral 2 covers the caudal portion of neural 1, neural 2, the cranial portion of neural 3, and the medial portion, both costal 1-3. The vertebral 3 covers the caudal portion of neural 3, neural 4, and most of neural 5, as well as medial parts, both costal 3 to 5. The cranial portion of vertebral 4 covers the most caudal portion of neural 5 and neural 6 and a short medial portion of costal bones 5 and 6.
Both pleural scutes 1 are trapezoidal, and the remaining are subrectangular, covering both sides of the carapace. Pleural 1 covers most of costal 1, the cranial portion of costal 2, and the slightly caudal portion of peripheral 2 and 3. Pleural 2 covers the caudal portion of costal 2, costal 3, and cranial portion of costal 4. Finally, pleural 3 covers the caudal portion of costal 4, costal 5, and the cranial portion of costal 6. As is typical in turtles in YPM 1565 (Figure 1A) the marginal scutes cover both sides of lateral extremes of the carapace. The marginal scutes 1-2 are trapezoidal, and marginals 3-7 are subrectangular, whereas the remaining posterior preserved are trapezoidal. The marginal scute 1 covers a lateral portion of the nuchal bone and the same portion of peripheral 1, whereas the remaining marginal scutes cover two alternative successive peripheral bones.
Discussion
Taxonomic Comments
The inclusion of the holotype of Parahydraspis paranensis plus the refereed specimens to the same taxon [48, plate II] in Pleurodira Chelidae is recognized by having the left half pelvic girdle (pubis, ischium and ilium) attached by suture to the shell as is seen in [48], plate II in specimen MLP 70-VIII-21-18, absence of mesoplastron (MLP-79- VIII-21.20), a cervical scute on the cranial margin of the carapace nuchal bone, and the loose attachment between carapace and plastron in the holotype YPM 1565 (Figures 1A & 2B). When Wieland, a century before, described Parahydraspis paranensis, he interpreted several pit bridges present in the peripheral bridge margin (Figures 1B & 1C) for the supposed reception of short plastral digitations and the existence of lateral hyo-hypoplastral fontanella (Figure 1C). However, the plastron is missing in the holotype. Considering that the Parahydrapis paranensis was erected by Wieland based on supposed structures not present in the preserved plastral bridge of extant Phrynops spp., where minor pitting is present in the peripheral border. The validation of the genus Parahydraspis is not possible. Besides, the presence of a continuous series of at least the last five neural plates and a possible contact between the nuchal and the neural 1 in YPM 1565 differentiates the material under study from Platemys, Acanthochelys, and Mesoclemmys [26, 55]. The holotype and same referred specimen of P. paranensis, in turn, differs from Platemys by the absence of the well-developed longitudinal ridges of the carapace and from Acanthochelys by the absence of a shallow midline depression of the carapace and from both aforementioned short-necked South American general by the presence of neural bones, and mid-size shell in comparison with the small size in Platemys platycephala and Aconthochelys spp., except for Acanthochelys macrocephala. YPM 1565 differs from most species of Mesoclemmys by having an uninterrupted neural series and a shell middle size. [55, 59, 94, 95] most species of Mesoclemmys [96], M. dahli [97], M. nasuta [65], M. raniceps [98], M. tuberculata [99], and some specimens of M. zuliae [67] lack neural bones or have a low number of neurals displaying in an interrupted series such as M. gibba, (MHNSR-Z -1551), and M. vanderhaegei (MZUC Ac-399) as is shown in [92], they do not reach the nuchal, allowing the medial contact between the costal 1 [55]. Only M. vanderhaegei is distributed in Argentina [92, 100, 101]. These extant Mesoclemmys species differ from extinct and extant Phrynops spp. in the carapace by the presence of three (MZUC- Ac 399) or four (MNHN- P unnumbered) discontinuous neurals as shown [102].
Acanthochelys and Phrynops sensu stricto is, until now, the only genera of Chelinae recorded from the late Miocene and Pliocene of Argentina. The specimens previously assigned to the latter genus come from outcrops of the El Morterito Fm. (late Miocene); see [103] at Valle del Cajon, Catamarca Province, and from the Andalhuala Fm (late Miocene) at Tio Punco locality, Tucuman Province [48]. As stated above, the genus Phrynops sensu stricto is currently comprised of the following extant species: P. geoffroanus, P. hilarii, P. tuberosus and P. williamsi, which are part of a supra-specific complex named by [64] as the P. geoffroanus complex [14, 25, 59, 92]. In addition, the extinct species P. paranaensis from the Late Miocene of Mesopotamia was also referred to in the aforementioned complex by [48]. Posteriorly, Kischlat, in its Magister Thesis, described in 1993 several specimens refereed, as unnamed spp., to the superspecies (Phrynops) (Phrynops) (geoffroanus) from Temembe Formation (late Oligocene-early Miocene), Tabuate Basin, Sao Paulo State, Brazil. The differences among species of Phrynops sensu stricto are mainly observed in characteristics of the scutes of the carapace and the carapace axillary buttress = anterior bridge strut scar; showing differences between the design of this structure and the angle between the rib/gomphosis and the anterior bridge strut suture scar on the first costal bone as is shown in Australasian chelids such as Elseya spp. and other short-necked taxa from Australasia [104, 105, 106, 107]. Differences in this structure on the visceral carapace are also distinguished in extant and extinct Phrynops spp. (see below).
Considering the similar plates and scute carapace and plastral morphological patterns between the Phrynops specimens from late Miocene El Morterito and Andalhuala Formations (PVL 3351, 3152 respectively), but the different morphology of the bridge strut suture scar between the specimen PVL 3351 from El Morterito Formation, Valle del Cajon (Catamarca province), [48, plate 1, figures. 1 & 2], some specimens MCT (ex DGM) 302 R, 1472-R, referred to as superspecies (Phrynops) (Phrynops) (geoffroanus) spp. (plates Figures 2, 4 & 5) than Phrynops paranensis (YPM 1565) [48, 93]. These conditions allow us to suggest that different Miocene species seem to be present in South America. P. paranensis is characterized by a diagnostic design of the anterior bridge structure with an enlarged medial end in contact with the suture between costal 1-2, and a different angle between rib/gomphosis and the anterior bridge strut scar in the holotype (YPM 1565) from upper Miocene “Conglomerado osifero” from Ituzaingó Formation at Parana (Entre Rios province). In contrast to P. paranensis (Figures 6A & 6B), a different design of the anterior bridge strut scar, and the angle between rib/gomphosis and the anterior bridge strut scar is shown in extant P. geoffroanus (AMNH 79048, MZUSP 2635) (Figures 6C & 6D), P. hilarii (FMNH 22481, MHNSR Z- 1550, NHMUK 86-3-10-1), (Figures 6E & 6F) and P. williamsi (MACN-H 38395, USNM 306646) (Figures 6G & 6H).
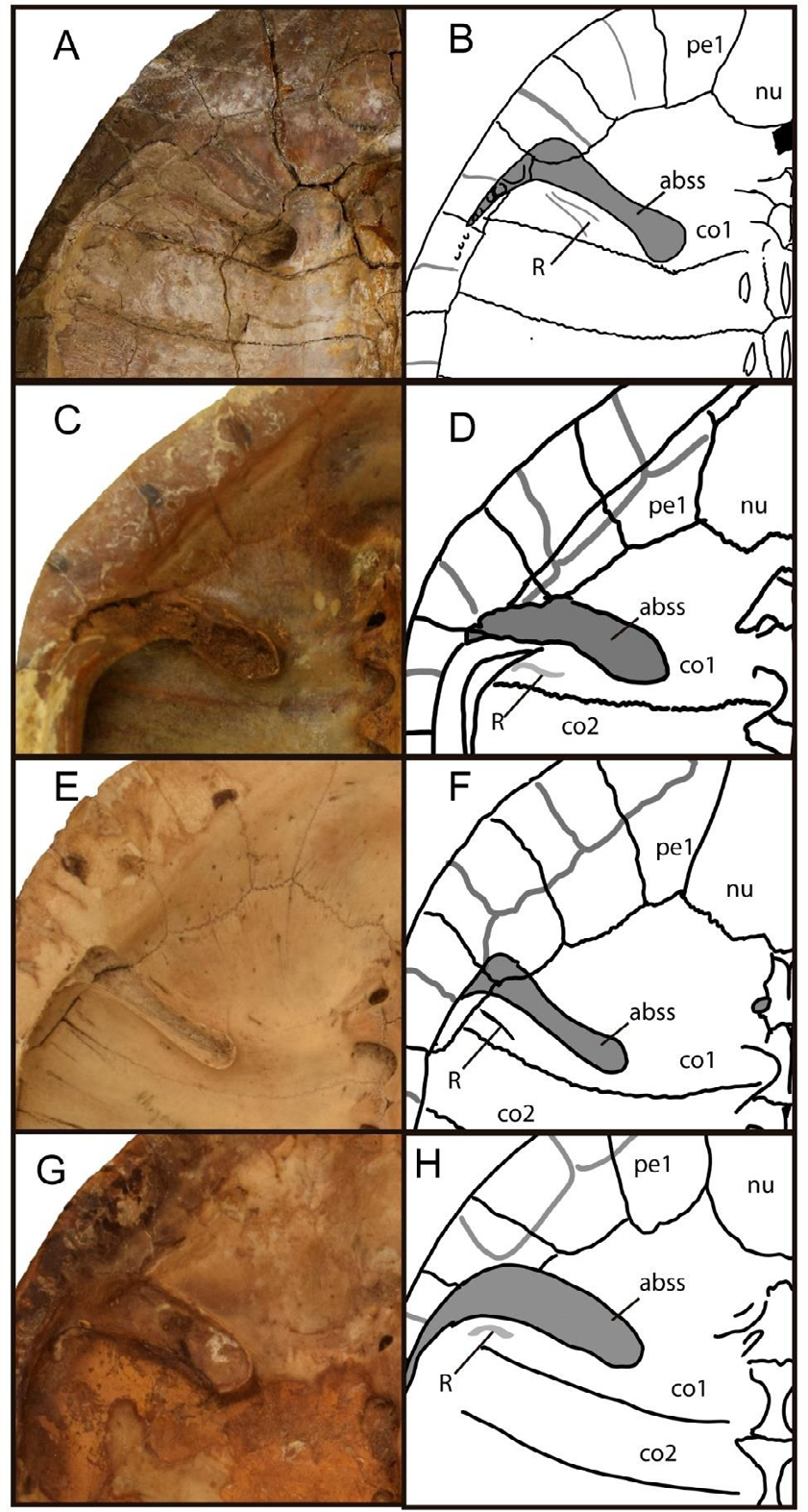
Figure 6: Photographs and drawings of visceral views of the anterior carapace in Phrynops spp., showing the anterior bridge carapace scar (abss) design and the angular relationships between it and the rib/ gomphosis (R) (A-B) Phrynops paranensis (YPM 1565), (C-D) P. geoffroanus (AMNH 79048), (E-F) P. hilarii (MHNSR-Z 1550), and (G-H) P. williamsi (MACN-H-38395).
Likely, the Phrynops sp. (FCVPV-319) described by [108] resembles P. paranensis. However, its assignation to P. paranensis seems to be possible, as was previously suggested by [108] in the relatively greater width of the cervical scute and the proportions of the last preserved marginals in the holotype of P. paranensis (YPM 1565) and FCVPV-319. However, until the visceral surface of the carapace of the Uruguayan specimen is exposed, the diagnostic condition of the anterior bridge strut scar present in the holotype YPM 3536 could not be identified in FCDPV-319, this assessment cannot be supported.
Although a recent total-evidence phylogeny [28] nested P. paranaensis inside a stem panchelid node composed of Cretaceous taxa with mesoplastra from Patagonia, this seems to be a spurious result due to the poor preservation of the shell of the holotype of Phrynops paranaensis. Whether a mesoplastron is absent in this Miocene taxon, as in post- Paleocene chelids, can only be determined with a preserved plastron in the holotype of P. paranaensis. However, from the hyo-hypoplastral contact as is seen in referred specimens (MLP 79-VIII-21-3 and MLP-79-VIII-21-20) recovered at “Conglomerado osifero” lower levels included in Ituzaingó’ Formation outcrop along the Parana river at Parana city suggest the absence of mesoplastron in these exemplars that can be referred to P. paranensis.
Paleoenvironment Setting from the “Conglomerado osifero” Ituzaingo Formation
A panoramic paleoenvironmental scenario from the “Conglomerado osifero” of Ituzaingó Formation was presented by [91]. According to these authors, the paleogeographic location of Parana, in the subtropical to temperate belts, is confirmed by the absence of crocodilids, podocnemidid turtles, and primates. The taxonomic reptile diversity, particularly, allows these authors to suggest a varied paleoenvironment to these lower levels of the Ituzaingo lithostratigraphic unit. For instance, the predominance of aquatic birds in these lower levels suggests the presence of woody lowlands and swamps along the riverbanks [91]. In addition, rheas and phorusrhacoids support a savannah-like near the river sides [109, 110]. The presence of glyptodonts allows [91] to suggest the existence of gallery forests outside of the pre-Parana River.
Considering tardigrade diversity must have been related to a heterogeneous landscape controlled by the Pre-Parana River and its affluents. The Interatheriidae Interatheriinae and proterotheriids notoungulates also seem related to woody and wet environments. The occurrence of porcupine rodents and procyionids also suggests forested paleoenvironments.
Conclusion
The re-study of holotype and the referred specimens to “Parahydraspis” paranensis from “Conglomerado osifero” Ituzaingó Formation crop out along Parana river at Parana city allows us to propose the validation of Phrynops paranensis as a full valid species. The continental terrestrial and aquatic fauna from the “Conglomerado osifero” indicate a warmer climate than the present one and suggest the presence of forested areas along the coasts of the pre-Parana River and its affluents.
This paleoenvironment was inhabited by the chelid turtle Phrynops paranensis.
Acknowledgments
We thank Juliana Sterli (Museo Paleontologico Egidio Feruglio, Trelew, Argentina) for sharing photographs of extant and extinct chelid turtles and Walter Joyce (University of Fribourg, Switzerland) for accessing of holotype of Parahydraspis paranensis. We also acknowledge for giving us access to extant and extinct chelids to Julian Faivovich, and Santiago Menda from the Herpetological collections (Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina) and Marcelo Reguero, Yanina Herrera, Sergio Vizcaino, Susana Bargo (Museo de la Plata, Argentina) and Jaime Powell (Instituto Miguel Lillo, Universidad Nacional de Tucuman, Argentina) from the Vertebrate Paleontological ones respectively. Finally, we thank David Kizirian. (American Museum of Natural History, New York, USA). and Vanessa Rhues (Peabody Museum, Yale University, New Haven, USA) for permitting us to publish photographs of specimens under their care. Besides, the editors of IZAB improved the quality of the original manuscript draft.
References
-
Broin FD (1984) Proganochelys ruchae n. sp., chélonien du Trias supérieur de Thaïlande [Proganochelys ruchae n. sp., chelonian from the Upper Triassic of Thailand]. Studia Geologica Salmanticensia 1: 87-97.
-
Gaffney ES (1990) The comparative osteology of the Triassic turtle Proganochelys. Bulletin of the American Museum of Natural History 194: 259-263.
-
Jenkins FA, Shubin NH, Amaral WW, Gattes SM, Schaff CR, et al. (1994) Late Triassic continental vertebrates and depositional environments of the Fleming Fjor Formation, Jameson Land, East Groenland. Meddelelser om Gronland Geosciences 32: 32-54.
-
Rougier GW, Fuente MS, Arcucci A (1995) Late Triassic turtles from South America. Science 268 (5112): 855- 858.
-
Sterli J, Fuente MS, Rougier G (2007) Anatomy and relationships of Palaeochersis talampayensis, a Late Triassic turtle from Argentina. Palaeontographica A 28: 1-61.
-
Joyce WG, Lucas SG, Scheyer TM, Heckert AB, Hunt AP (2009) A thin-shelled reptile from the Late Triassic of North America and the origin of the turtle shell. Proceedings of the Royal Society B 276(1656): 507-513.
-
Szczygielski T, Sulej T (2016) Revision of the Triassic European turtles Proterochersis and Murrhardtia (Reptilia, Testudinata, Proterochersidae), with the description of new taxa from Poland and Germany. Zoological Journal of the Linnean Society 177(2): 395- 427.
-
Joyce WG (2017) A Review of the Fossil Record of Basal Mesozoic Turtles. Bulletin of the Peabody Museum of Natural History 58(1): 65-113.
-
Sterli J, Martinez RN, Cerda IA, Apaldetti C (2021) Appearances can be deceptive: bizarre shell microanatomy and histology in a new Triassic turtle (Testudinata) from Argentina at the dawn of turtles. Papers in Palaentol 7(2): 1097-1132.
-
Fuente MS, Sterli J, Krapovickas V (2021) Triassic Turtles from Pangea: The legacy from South America. Journal of South American Earth Sciences 105.
-
Lyson TRG, Bever S (2020) Origin and evolution of the turtle body plan. Annu Rev Ecol Evol Syst 51: 143-166.
-
Benson RBJ, Domokos G, Varkonyi PL, Reisz RR (2011) Shell geometry and habitat determination in extinct and extant turtles (Reptilia: Testudinata). Paleobiology 37(4): 547-562.
-
Nicholson DB, Holroyd PA, Benson RB J, Barrett PM (2015) Climate-mediated diversification of turtles in the Cretaceous. Nat. Commun 6: 78-48.
-
Rhodin AGJ, Iverson JB, Bour R, Fritz U, Georges A, et al. (2021) Turtles of the World: Annotated Checklist and Atlas of Taxonomy, Synonymy, Distribution, and Conservation Status. In: Rhodin AGJ, Pritchard PCH, et al. (Eds.), 9th (Edn.), Conservation Biology of Freshwater Turtles and Tortoises: A Compilation Project of the IUCN/SSC Tortoise and Freshwater Turtle Specialist Group. Chelonian Research Monographs 8: 1-472.
-
Wilson DE, Reeder DAM (2005) Mammal Species of the World: A Taxonomic and Geographic Reference. Johns Hopkins University Press, 88(3): 824-830.
-
Clements JF, Rasmussen PC, Schulenberg TS, Iliff MJ, Fredericks TA, et al. (2020) The eBird/Clements Checklist of Birds of the World. Cornell University.
-
Gill F, Donsker D, Rasmussen P (2020) IOC World Bird List, volume 10(2).
-
Uetz P, Freed P, Hoseck J, Aguilar R, Reyes F, et al. (2020) The Reptile Database.
-
Thomson RC, Spinks PD, Broadly Shaffer H (2021) A global phylogeny of turtles reveals a burst of climate- associated diversification on continental margins Proceedings of the National Academy of Sciences 118(7): e2012215118.
-
Joyce WG, Parham JF, Lyson TR, Warnock CM, Donoghue PCJ (2013) A divergence dating analysis of turtles using fossil calibrations: An example of best practices. J Paleontol 87(4): 612-634.
-
Shaffer HB, McCartney-Melstad ET, Near J, Mount GG, Spinks PQ (2017) Phylogenomic analyses of 539 highly informative loci dates a fully resolved time tree for the major clades of living turtles (Testudines). Mol Phylogenet Evol 115: 7-15.
-
Pereira AG, Sterli J, Moreira, FRR, Schrago CG (2017) Multilocus phylogeny and statistical biogeography clarify the evolutionary history of major lineages of turtles. Mol Phylogenet Evol 113: 59-66.
-
Warnock RCM, Parham JF, Joyce WG, Lyson TR, Donoghue PCJ (2015) Calibration uncertainty in molecular dating analyses: There is no substitute for the prior evaluation of time priors. Proc Biol Sc 282(1798): 20141013
-
Near TJ, Meylan PA, Shaffer HB (2005) Assessing concordance of fossil calibration points in molecular clock studies: An example using turtles. Am Nat 165(2): 137-146
-
de la Fuente MS, Sterli J, Maniel I (2014) Origin, Evolution and Biogeographic History of South American Turtles. Springer, Earth System Sciences, Germany, pp: 168.
-
Maniel IJ, de la Fuente MS (2016) A review of the fossil record of turtles of the clade Pan-Chelidae. Bulletin of the Peabody Museum of Natural History 57(2): 191-227.
-
Vlachos E, Randolfe E, Sterli J, Leardi JM (2018) Changes in the diversity of turtles (Testudinata) in South America from the Late Triassic to the Present. Ameghiniana 55(6): 619-644.
-
Holley JA, Sterli J, Basso NG (2020) Dating the origin and diversification of Pan-Chelidae (Testudines, Pleurodira) under multiple molecular clock approaches. Contributions to Zoology 89(2): 146-174.
-
Broin F de, de la Fuente MS (1993) Les tortues fósiles d’Argentine: Synthèse. Annales de Paléontologie 79, 169–232.
-
de la Fuente M, Lapparent de Broin F de, Manera de Bianco T (2001) The oldest and first nearly complete skeleton of a chelid, of the Hydromedusa group (Chelidae, Pleurodira), from the Upper Cretaceous of Patagonia. Bulletin de la Societe Geologique de France 172(2): 237- 244.
-
Lapparent de Broin F de, de la Fuente M (2001) Oldest world Chelidae (Chelonii, Pleurodira), from the Cretaceous of Patagonia. Comptes Rendues Academie des Sciences de Paris 333: 463-470.
-
Bona P, de la Fuente MS (2005) Phylogenetic and paleobiogeographic implications of Yaminuechelys maior (Staesche, 1929) new comb. A large long-necked chelid turtle from the Early Paleocene of Patagonia. Journal of Vertebrate Paleontology 25(3): 569-582.
-
de la Fuente MS (2003) Two new Pleurodiran turtles from the Portezuelo Formation (Upper Cretaceous) of northern Patagonia, Argentina. Journal of Paleontology 77(3): 559-575.
-
Bona P (2006) Paleocene (Danian) chelid turtles from Patagonia, Argentina: taxonomic and biogeographic implications. N Jahr Geol Palaont 241(3): 303-323.
-
de la Fuente MS (2007) Testudines in Patagonian Mesozoic Reptiles. Gasparini Z, Coria R, et al. (Eds.), Indiana University Press, Indiana, USA, pp: 50-86.
-
de la Fuente MS, Umazano AM, Sterli J, Carballido JI (2011) New chelid of the lower section of the Cerro Barcino formation (Aptian-Albian?), Patagonia. Cretaceous Research 32(4): 527-537.
-
de la Fuente MS, Maniel I, Jannello JM, Filippi L, Cerda I (2015) Long-necked chelid turtles from the Campanian of northwestern Patagonia with comments on K/P survivorship of the genus Yaminuechelys. Comptes Rendus Palevol 14(6-7): 563-576.
-
de la Fuente MS, Maniel IJ, Jannello JM, Sterli J, Riga BJG, et al. (2017a) A new large panchelid turtle (Pleurodira) from the Loncoche Formation (upper Campanian-lower Maastrichtian) of the Mendoza Province (Argentina): Morphological, osteohistological studies, and a preliminary phylogenetic analysis. Cretaceous Research 69: 147-168.
-
de la Fuente MS, Maniel IJ, Jannello JM, Sterli J, Garrido A, et al. (2017) Unusual shell anatomy and osteohistology in a new Upper Cretaceous panchelid turtle from northwestern Patagonia (Argentina). Acta Palaeontologica Polonica 62: 585-601.
-
Maniel IJ, de la Fuente MS, Sterli J, Jannello JM, Krause MJ (2018) New remains of the aquatic turtle Hydromedusa casamayorensis (Pleurodira: Chelidae) from the Middle Eocene of Patagonia: Taxonomic validation and phylogenetic relationships. Papers in Palaeontology 4(4): 537-566.
-
Maniel IJ, de la Fuente MS, Apesteguia S, Perez Mayoral J, Sanchez ML, et al. (2020) Cranial and postcranial remains of a new species of Prochelidella (Testudines: Pleurodira: Chelidae) from ‘La Buitrera’ (Cenomanian of Patagonia, Argentina), with comments on the monophyly of this extinct chelid genus from southern Gondwana. Journal of Systematic Palaeontology 18(12): 1033-1055.
-
Maniel IJ, de la Fuente MS, Filippi LF (2023) A new chelid turtle, Prochelidella palomoi, sp. nov. from Campanian Anacleto Formation, Neuquen Basin, north-western Patagonia, Argentina. The Anatomical Record 306: 1365-1376.
-
Lapparent de Broin F, Molnar R (2001) Eocene chelid turtles from Redbank Plains, Southeast Queensland, Australia. Geodiversitas 23(1): 41-79.
-
Smith ET (2010) Early Cretaceous chelids from Lightning Ridge, New South Wales. Alcheringa 34: 375-384.
-
De Vis C W (1897) The extinct freshwater turtles of Queensland. Annals of the Queensland Museum 3: 3-7.
-
Wieland G (1923) A new Parana pleurodiran. American Journal of Science 5: 1-14.
-
Wood RC (1976) Two new species of Chelus (Testudines: Pleurodira) from the late Tertiary of Northern South America. Breviora 435: 1-26.
-
de la Fuente MS (1992) Las tortugas Chelidae del Terciario superior y Cuaternario del territorio argentino. Ameghiniana 29(3): 211-299.
-
Gaffney ES (1979) Fossil chelid turtles of Australia. American Museum Novitates 2681: 1-23.
-
Gaffney ES, Archer M, White A (1989) Chelid turtles from the Miocene freshwater limestones of Riversleigh Station, northwestern Queensland, Australia. American Museum Novitates 2959: 1-10.
-
White AW, Archer M (1994) Emydura lavarockorum, a new Pleistocene turtle (Pleurodira: Chelidae) from fluviatile deposits at Riversleigh, northwestern Queensland. Records of the South Australian Museum 27: 159-167.
-
Megirian D, Murray P (1999) Chelid turtles (Pleurodira, Chelidae) from the Miocene Camfield Beds, Northern Territory of Australia, with a description of a new genus and species. The Beagle 15: 75- 130.
-
Thomson SA, MacKness BS (1999) Fossil turtles from the early Pliocene Bluff Downs Local Fauna, with a description of a new species of Elseya. Transactions of the Royal Society of South Australia 123(3): 101-105.
-
Yates AM (2013) A new species of long-necked turtle (Pleurodira: Chelidae: Chelodina) from the late Miocene Alcoota Local Fauna, Northern Territory, Australia. PeerJ pp: e170.
-
Cadena EA, Vanegas A, Jaramillo C, Cottle JM, Johnson TA (2020) A new Miocene turtle from Colombia sheds light on the evolutionary history of the extant genus Mesoclemmys Gray, 1873. Journal of Vertebrate Paleontology 39(5).
-
Gray JE (1873) Observations on chelonians, with description of new genera and species. Journal of Natural History 11: 289-308.
-
Dumeril AMC (1806) Analytical zoology, or natural method of classifying animals, made easier using synoptic tables. Allais, Paris, USA, pp: 1‐344.
-
Wagler J (1830) Natural system of amphibians, preceded by the classification of mammals and birds, a contribution to comparative zoology. JG Cottaschen Buchhandlung, Germany.
-
McCord WP, Joseph-Ouni M, Lamar WW (2001) A taxonomic reevaluation of Phrynops (Testudines: Chelidae) with the description of two new genera and a new species of Batrachemys. Revista de Biologia Tropical 49(2): 715-764.
-
Rhodin AGJ, Mittermeier RA, McMorris JR (1984) Platcmys macrocephala a new species of chelid turtle from central Bolivia and the Pantanal region of Brazil. Herpetologica 40: 38-46.
-
Freiberg M (1945) A new species of the genus Platemys Wagler 20(55): 19-23.
-
Dume ́ril AMC, Bibron G (1835) Erpe ́tologi ́e Generale ou Histire Naturelle des Reptiles. In: Nouvelles suites à Buffon. (Edn.), Biodiversity Heritage Library, Paris, USA.
-
Mikan JC, Sandler W, Knapp J, Miken JC, Strauss A (1820) A selection of Brazilian flora and fauna: researched by order and under the auspices of Francis I, emperor of Austria. In: Nissen C (Ed.), 2nd (Edn.), Botanical Book Illustration, Vienna, Austria 723: 789.
-
Rhodin AGJ, Mittermeier RA (1983) Description of Phrynops williamsi, a new species of chelid tunle of the South American P. geoffroanus complex. In: Rhodin AGJ, (eds). Advances in Herpetology and Evolutionary Biology. (Edn.), Mus Comp Zool, Harvard Cambridge, Massachussets, USA, pp: 58-73.
-
Schweiger AF (1812) Prodomus Monographiae Cheloniorum. Konigsberg Arch Naturwiss Math 1: 171- 368.
-
Peters WCH (1870) Uber Platemys tuberosa, a new species of turtle from British Guiana. Monatsber co ̈ nigl Acad Wiss Berlin pp: 311-313.
-
Pritchard PCH, Trebbau P (1984) The Turtles of Venezuela. New York, USA.
-
Ernst CH, Barbour RW (1989) Turtles of the World. In: Smithsonian Institution Press, Washington DC, London, UK.
-
Froes OM (1957) Notas chelonologicas Update of the nomenclature of the Brazilian chelo. nios Inheringia Zool 2: 1-24.
-
David P (1994) List of current reptiles of the world. I: Chelonii Dumerilia 1: 7-27.
-
Fritz E, Obst FJ, Gunther R (1994) Kritischer Typen Katalog der Shildkro¨ tensammlung (Reptilia: Testudines). des Zoologischen Museums Berlin Mitt Zool Mus Berlin 70: 157-175.
-
Rhodin AGJ, Thomson S, Georgalis G, Karl HV, Danilov IG, et al. (2015) Turtles and tortoises of the world during the rise and global spread of humanity: first checklist of extinct Pleistocene and Holocene chelonians. In: Rhodin AGJ (Ed.), Chelonian Research Monographs (Edn.), Switzerland 5(8): 1-66.
-
Zangerl R (1969) The turtle shell. In: Gans C (Ed.), Biology of the Reptilia. Academic Press, New York, USA, pp: 311-339.
-
Gibbard PL, Harper DAT (2020) International Commission on Stratigraphy. Annual Report pp: 1-3.
-
Cohen KM, Finney SC, Gibbard JX (2013) The ICS International Chronostratigraphic Chart. Geology 36(3): 199-204.
-
Klein T (1760) Klein’s Classification and Short History of Four-Feded Animals. In: Behn FD, (Eds.), Lübeck, Germany, pp: 381.
-
Joyce WG, Parham JF, Anquetin J, Claude J, Danilov IG, et al. (2020) Phylonyms A Companion to the PhyloCode. In: De Queiroz (Ed.), CRC Press, Boca Raton, USA, pp: 1045- 1048.
-
Batch AJGC (1788) Versuch einer Anleitung zur Kenntniss und Geschichte der Thiere und Mineralien: für akademische Vorlesungen entworfen, und mit den nöthigsten Abbildungen versehen. Jena, in der Akademischen Buchhandlung, Germany.
-
Joyce W G, Parham J F, Anquetin J, Claude J, Danilov I G, Iverson J B, Kear B, Lyson T R, Rabi M, Sterli J (2020b). Testudines. 1049–1051. In De Queiroz, K. Cantino, P. D. and Gauthier, J. A (eds.) Phylonyms – A Companion to the PhyloCode. CRC Press, Boca Raton.
-
Cope ED (1865) Third Contribution to the Herpetology of Tropical America. Proceedings of the Academy of Natural Sciences of Philadelphia 17(4): 185-198.
-
Joyce WG, Parham JF, Anquetin J, Claude J, Danilov IG, et al. (2020c) Phylonyms A Companion to the PhyloCode. In: De Queiroz K, Cantino PD, et al. (Eds.), CRC Press, Boca Raton, Testudines pp: 1055-1057.
-
Joyce WG, Parham JF, Gauthier A (2004) Developing a protocol for the conversion of rank-based taxon names to phylogenetically defined clade names, as exemplified by turtles. Journal of Paleontology 78: 989-1013.
-
Lindohlm WA (1929) Revised list of the genera of modern snail toads along with notes on the nomenclature of some species. Zoolgischer Anzeiger 81: 272-275.
-
Joyce WG, Anquetin J, Cadena E A, Claude J, Danilov IG, et al. (2021) A nomenclature for fossil and living turtles using phylogenetically defined clade names. Swiss Journal of Palaeontology 140: 1-45.
-
Gray JE (1825) A synopsis of the genera of reptiles and amphibians with description of new 85. species. Annales of Philosophy 2(10): 193-217.
-
Gasparini Z, Baez AM (1975) Contributions to the knowledge of the tertiary herpetofauna of Argentina. In: Proceedings of the Argentine Congress of Paleontology and Biostratigraphy, Tucuma, Argentina 2: 377-415.
-
Baez AM, Gasparini Z (1977) Origenes y evolucion de los anfibios y reptiles del Cenozoico de America del Sur. Acta Geologica Lilloana 14: 149-232.
-
Mlynarski M (1976) Handbuch der Pala¨oherpetologie, Part 7, Testudines. Stuttgart: Gustav Fischer Verlag.
-
Gasparini ZB, de la Fuente MS, Donadio O (1986) Argentine Cenozoic Reptiles: Paleoenvironmental implications and biogeographic evolution. Minutes 4th Congress. Arg Paleontol Bioestr 2: 119-130.
-
Arganaraz B, Pina CI (2000) Chelonians from the Ituzaingo Formation (upper Miocene-Pliocene), in Villa Urquiza, Entre Rios, Argentina. In: AcenOolaza FG, Herbst R (Eds.), El Neogeno de Argentina Serie Correlacion Geologica 14: 239-244.
-
Cione AL, Azpelicueta MM, Bond M, Carlini AA, Casciotta JR, et al. (2000) Miocene vertebrates from Entre Rios province, eastern Argentina. In: Acenolaza FG, Herbst R (Eds.), El Neogeno de Argentina. Serie Correlacion Geologica 14: 191-237.
-
Cabrera MR (2022) The continental turtles of southern South America. Universidad Nacional de Cordoba, pp: 216.
-
Kishlat EE (1993) Contribution to the knowledge of recent fossils from Brazil. Quelidas (Chelonii, Pleurodira) from the Taubate Basin, Cenozoic of the State of São Paulo, Brazil [master’s Rio de Janeiro. Federal University of Rio de Janeiro, Brazil, pp: 188.
-
Pritchard PCH (1988) A survey of neural bone variation among recent chelonian species, with functional interpretations. Acta Zoologica Cracoviensia 31: 625- 686.
-
Smales I (2019) A review of neurals in Chelidae (Testudines: Pleurodira) with reference to phylogeny of the family. Chelonian Conservation and Biology 18: 32- 61.
-
Gray JE (1873) Hand-list of the specimens of shield reptiles in British Museum. British Museum, London, UK.
-
Zangerl R, Medem F (1958) A new species of chelid turtle, Phrynops (Batrachemys) dahli, from Colombia. Bulletin of the Museum of Comparative Zoology 119: 375-390.
-
Gray J E (1855) Catalogue of Shield Reptiles in the Collection of the British Museum. Part 1. Testudinata (Tortoises). Taylor and Francis, London, UK.
-
Luederwaldt H (1926) Brazilian chelonians with the list of species from the Museu Paulista. Revista Museu Paulista 14: 405-470.
-
Bour R, Zaher H (2005) A new species of Mesoclemmys, from the open formations of Northeastern Brazil (Chelonii: Chelidae). Pap Avul Zool 45: 295-311.
-
Baldo D, Martinez P, Boeris JM, Giraudo R (2007) Reptilia, Chelonii, Chelidae, Phrynops geoffroanus Schweigger, 1812 and Mesoclemmys vanderhaegei (Bour, 1973): distribution extension, new country record, and new province records in Argentina. Check List 3(4): 348-352.
-
Bour R, Pauler I (1987) The identity of Phrynops vanderhaegei Bour 1973 and related species. Mesogee 47: 3-23.
-
Bossi GE, Vides ME, Georgieff SM, Muruaga CM, Ibanez LM (2000) Analysis of paleocurrents and the variance of the components at three levels, Neogene of the Cajon Valley, Catamarca, Argentina. Magazine of the Argentine Association of Sedimentology 7: 23-47.
-
Thomson SA, White A, Georges A (1997) Re- evaluation of Emydura lavarackorum: Identification of a living fossil. Memoirs of the Queensland Museum 42: 327-336.
-
Thomson SA (2015) Diagnosis of living and fossil short-necked turtles of the genus Elseya using skeletal morphology. Master of Applied Science Thesis, University of Canberra, Canberra, Australia, pp: 112.
-
Joshep-Ouni M, Cann J, McCord WP, Frewer J (2023) Discovery of extant Elseya (Elseya) lavarackorum and confirmation of two lineages of “ living fossil” in the Nicholson-Gregory drainage, with a visual guide to scutation and osteological suture characters in northern Australia Elseya (Testudines: Chelidae). The Batagur Monographs 10: 7-119.
-
Thomson SA, Friol NR, White A, Wedd D, Georges A (2023) The Australian gulf snapping turtle Elseya lavarackorum (Testudines: Chelidae) revisited―Is the late Pleistocene fossil species extant?. Vertebrate Zoology 73: 237-256.
-
Perea D, Ubilla M, Pineiro G (1996) First Fossil Record (Late Miocene) of Phrynops (P. geoffroanus) from Uruguay: biostratigraphical and Palaeoenvironmental Context. Copeia 1996(2): 445-451.
-
Noriega JI (1994) The Birds of the “Mesopotamian” of the province of Entre Rios, Argentina. National University of La Plata, Argentina 611: 162.
-
Noriega JI (1995) The Avifauna from the “Mesopotamian” (Ituzaingo Formation: Upper Miocene) of Entre Ríos Province. Courier Forschungsinstitut Senckenberg, Argentina 181: 141-148.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework