Cell Membrane Compartmentalization and Membrane Dynamics during Plasmodium Infection
Plasmodium undergoes a sophisticated invasion process to enter red blood cells (RBCs), a critical step in its life cycle. The invasion process is intricately regulated and involves a series of well-coordinated events. Membrane compartmentalization and dynamics play a central role in this process.
Abbreviations
CD47: Cluter of Differentiation; CR1: Complement Receptor Type 1; EBAs: Erythrocyte Binding Antigens; NPPs: New Permeability Pathways; PfEMP1: Plasmodium Falciparum Erythrocyte Membrane Protein 1; PV: Parasitophorous Vacuole; PVM: Parasitophorous Vacuole Membrane; RBCs: Red Blood Cells; RONs: Rhoptry Neck Proteins.
Plasmodium undergoes a sophisticated invasion process to enter red blood cells (RBCs), a critical step in its life cycle. The invasion process is intricately regulated and involves a series of well-coordinated events. Membrane compartmentalization and dynamics play a central role in this process. The parasite actively reorganizes its membranes and manipulates those of the host cell to facilitate its entry. Once inside, it resides within a parasitophorous vacuole, a membrane-bound compartment that shields it from the host’s immune defenses while allowing it to exploit host resources. Understanding molecular and cellular mechanisms underlying these membrane interactions provides valuable insights into potential therapeutic targets.
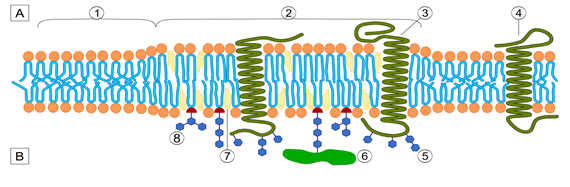
Membrane compartmentalization organizes the plasma membrane into distinct domains with specialized functions. Plasmodium infection takes advantage of this compartmentalization through various mechanisms. Lipids rafts, rich in cholesterol and sphingolipids, allow the parasite’s entry into host cells. These domains serve as platforms for receptor clustering, facilitating the interaction between the parasite and the host cell membrane. After invasion, Plasmodium resides within a parasitophorous vacuole (PV) derived from the host cell membrane. This compartment acts as a protective niche, separating the parasite from the host cytoplasm while allowing nutrient acquisition and waste disposal. The parasite induces compartmentalization within the host cell, forming structures such as Maurer’s clefts. These are membrane-bound compartments essential for protein trafficking and erythrocyte remodeling (Figure 1) [1, 2, 3].
Membrane dynamics encompass the processes of deformation, fusion, and trafficking within the cell membrane. The Plasmodium parasite exploits these mechanisms to support its lifecycle. A key stage is the invasion of erythrocytes by Plasmodium merozoites, a highly coordinated and complex process that induces transient deformations in the host cell membrane. The invasion begins with the merozoite reversibly attaching to the erythrocyte surface by binding to specific receptors, such as glycophorins and complement receptors. It is followed by an irreversible reorientation of the merozoite, aligning its apical pole with the erythrocyte membrane. A tight junction forms a specialized protein complex that establishes close contact between the merozoite and the host cell. This junction acts as a seal, guiding the invagination of the erythrocyte membrane to engulf the parasite, ultimately leading to the formation of the parasitophorous vacuole (PV). The PV is a specialized compartment that protects and nourishes the parasite within the host cell. During the formation of the PV, the parasite deploys proteins such as Rhoptry Neck Proteins (RONs) and Erythrocyte Binding Antigens (EBAs) to mediate membrane fusion events, ensuring a seamless entry into the host cell. These interactions are crucial for the successful invasion and survival of the parasite. The parasitophorous vacuolar membrane is a hybrid structure derived from the parasite and the erythrocyte, enabling the parasite to acquire nutrients and evade immune detection. During the invasion, the parasite sheds its surface coat. It releases contents from secretory organelles, each serving distinct functions. Micronemes facilitate adhesion and junction formation, rhoptries modify the host cell, and dense granules maintain the vacuole. This process showcases the parasite’s sophisticated adaptations for survival and growth within its host (Figure 2) [4, 5, 6].
Once inside an erythrocyte, Plasmodium reprograms the host cell to suit its survival needs. The parasite sends proteins across the parasitophorous vacuole membrane (PVM) into the host’s cytoplasm and membrane, triggering significant changes. These modifications affect the erythrocyte’s stiffness, permeability, and surface protein makeup. By creating New Permeability Pathways (NPPs) facilitated by parasite- formed channels and altered host transporters, the infected cell becomes more accessible to nutrients. Additionally, Plasmodium reshapes the erythrocyte’s cytoskeleton and lipid composition, increasing its flexibility-a vital adaptation that helps the parasite navigate narrow microcapillaries and evade splenic filtration [7].
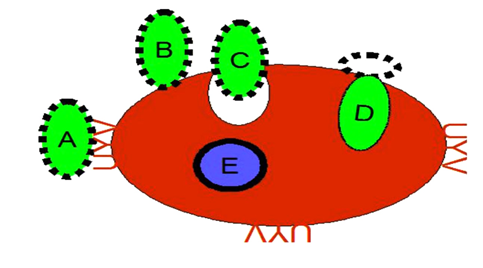
Plasmodium parasites remodel red blood cells (RBCs) to survive and evade the host immune system. They form surface knobs packed with cytoadhesion proteins like PfEMP1 (Plasmodium falciparum Erythrocyte Membrane Protein 1), enabling infected RBCs to adhere to endothelial cells and avoid spleen clearance. Inside RBCs, the parasites reside in parasitophorous vacuoles, which contain tubo- vesicular networks and detergent-resistant membranes rich in lipids, supporting parasite growth and nutrient acquisition. The Maurer’s clefts direct parasite proteins like PfEMP1 to the RBC surface, ensuring functional knob formation and cytoadherence, which can lead to severe complications such as cerebral and placental malaria. Infected RBCs also exhibit increased metabolic activity, enhancing nutrient uptake and waste removal via modified membrane channels. While aiding parasite survival, these adaptations offer potential targets for therapeutic interventions and vaccine development. The ability of Plasmodium to manipulate membrane dynamics is essential to evade immune detection. In Plasmodium falciparum infections, the surface of infected red blood cells develops specialized protrusions known as knobs. These structures anchor cytoadherence proteins like PfEMP1, enabling the infected cells to adhere to microvascular walls and avoid clearance by the spleen. Additionally, the parasite alters or removes specific host membrane proteins, such as CD47 and CR1, to further reduce the likelihood of immune recognition (Figure 3) [8, 9].
A deeper understanding of membrane compartmentalization and dynamics during Plasmodium infection reveals promising therapeutic opportunities. Interfering with lipid rafts or receptor-ligand interactions could block merozoite entry while disrupting the export of parasite proteins, which could hinder erythrocyte remodeling and compromise parasite survival. Targeting membrane permeability, modulating New Permeability Pathways (NPPs), or inhibiting ion channels could deprive the parasite of essential nutrients. These processes are central to Plasmodium’s capacity to invade and thrive within host cells. The complex interaction between parasite and host membranes drives malaria pathogenesis and highlights critical vulnerabilities for intervention [10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39].
Acknowledgments
The author acknowledges Mrs. Norri Zahra and Mr. Regragui Moumaris. The author thinks Prof. Nisen Abuaf (Sorbonne University and AP-HP). The author thinks Tech. Said Youssouf Chanfi (Sorbonne University). The author thinks Ing. Jean-Michel Bretagne (AP-HP). The author thinks Clr. Marie-Hélène Maës and Clr. Monique Abi (Research and Development Company).
References
-
Koudatsu S, Masatani T, Konishi R, Asada M, Hakimi H, et al. (2021) Glycosphingolipid GM3 is localized in both exoplasmic and cytoplasmic leaflets of Plasmodium falciparum malaria parasite plasma membrane. Sci Rep 11(1): 14890.
-
Bindschedler A, Schmuckli-Maurer J, Buchser S, Fischer TD, Wacker R, et al. (2024) LC3B labeling of the parasitophorous vacuole membrane of Plasmodium berghei liver stage parasites depends on the V-ATPase and ATG16L1. Mol Microbiol 121(6): 1095-1111.
-
Omoda A, Matsumoto K, Yoshino KI, Tachibana M, Tsuboi T, et al. (2024) Skeleton binding protein 1 localizes to the Maurer’s cleft and interacts with PfHSP70-1 and PfHSP70-x in Plasmodium falciparum gametocyte- infected erythrocytes. Parasitol Int 100: 102864.
-
Cooke BM, Mohandas N, Coppel RL (2001) The malaria- infected red blood cell: structural and functional changes. Adv Parasitol 50: 1-86.
-
Geoghegan ND, Evelyn C, Whitehead LW, Pasternak M, McDonald P, et al. (2021) 4D analysis of malaria parasite invasion offers insights into erythrocyte membrane remodeling and parasitophorous vacuole formation. Nat Commun 12(1): 3620.
-
Bannister LH, Hopkins JM, Margos G, Dluzewski AR, Mitchell GH (2004) Three-dimensional ultrastructure of the ring stage of Plasmodium falciparum: evidence for export pathways. Microsc Microanal 10(5): 551-562.
-
Fowkes FJ, Michon P, Pilling L, Ripley RM, Tavul L, et al. (2008) Host erythrocyte polymorphisms and exposure to _Plasmodium falciparum_ in Papua New Guinea. Malar J 7:1.
-
Mansouri R, Ali-Hassanzadeh M, Shafiei R, Savardashtaki A, Karimazar M, et al. (2020) The use of proteomics for the identification of promising vaccine and diagnostic biomarkers in Plasmodium falciparum. Parasitology 147(12): 1255-1262.
-
McHugh E, Carmo OMS, Blanch A, Looker O, Liu B, et al. (2020) Role of Plasmodium falciparum Protein GEXP07 in Maurer’s Cleft Morphology, Knob Architecture, and _P._ _falciparum_ EMP1 Trafficking. mBio 11(2): e03320.
-
Moumaris M (2024) Advancements in Magnetic Resonance Imaging: Transforming Non-Invasive Diagnosis and Treatment Monitoring in Radiology. J Life Sci Res and Rev 2(5): 1-3.
-
Moumaris M (2024) Plasmodium’s Secret: How a Complex Endomembrane System Drives Malaria’s Deadly Efficiency. Int J Zoo Animal Biol 7(6): 000631.
-
Moumaris M (2024) Malaria’s Hidden Weapon: How Plasmodium Transforms Red Blood Cells to Evade and Invade. Int J Zoo Animal Biol 7(6): 000629.
-
Moumaris M (2024) Advancements in Diagnosing and Treatments Plasmodium knowlesi: Challenges and Innovations. Int J Zoo Animal Biol 7(5): 000622.
-
Moumaris M (2024) Confronting Plasmodium knowlesi: Challenges and Strategies in Malaria Healthcare. Int J Zoo Animal Biol 7(4): 000607.
-
Moumaris M (2024) Unraveling the Enigma: Tackling Knowlesi Malaria in Southeast Asia. Int J Zoo Animal Biol 7(3): 000585.
-
Moumaris M (2024) Unveiling the Enigmatic Plasmodium knowlesi: Insights, Challenges, and Promises in Malaria Research. Int J Zoo Animal Biol 7(2):000566.
-
Moumaris M (2024) Unlocking the Potential: Overcoming Challenges in CAR-T Cell Therapy for Cancer Treatment. J Biotechnology and Bioprocessing 5(2): 2766-2314.
-
Moumaris M (2023) Revolutionizing Malaria Research: CRISPR unveils New Frontiers. J Biotechnology and Bioprocessing 4(5): 2766-2314.
-
Moumaris M (2024) Lyme Disease: A Zoonosis Tick- Borne Borrelia Bacterium [4/4]. Int J Zoo Animal Biol 7(1): 000549.
-
Moumaris M (2023) Lyme Disease: A Zoonosis Tick- Borne Borrelia Bacterium [3/4]. Int J Zoo Animal Biol 6(4): 000500.
-
Moumaris M (2023) Lyme Disease: A Zoonosis Tick- Borne Borrelia Bacterium [2/4]. Int J Zoo Animal Biol 6(2): 000465.
-
Moumaris M (2022) Lyme Disease: A Zoonosis Tick- Borne Borrelia Bacterium [1/4]. Int J Zoo Animal Biol 5(6): 000425.
-
Moumaris M, Bretagne JM, Abuaf N (2020) Nanomedical Devices and Cancer Theranostics. The Open Nanomedicine and Nanotechnology Journal 6: 1-11.
-
Moumaris M, Bretagne JM, Abuaf N (2019) Biological Membranes and Malaria-Parasites. The Open Parasitology Journal 7: 1-18.
-
Moumaris M, Bretagne JM, Abuaf N (2018) Hospital Engineering of Medical Devices in France. The Open Medical Devices Journal 6: 10-20.
-
Moumaris M, Rajoely B, Abuaf N (2015) Fluorescein Isothiocyanate-Dextran can track Apoptosis and Necrosis induced by heat shock of Peripheral Blood Mononuclear Cells and HeLa Cells. Open Biological Sciences Journal 1: 7-15.
-
Moumaris M, Rajoely B, Abuaf N (2012) The Naïve B Cells are the Lymphocytes with the Highest Anionic Phospholipid Binding Ratios. The Open Immunology Journal 5: 27-35.
-
Moumaris M (2007) Magnetic resonance imaging at the Hôtel-Dieu of Paris. Paris-Descartes University, Unités de formation et de recherche (UFR) biomédicale des Saints- Pères, France.
-
Moumaris M (2005) Identification of a new molecule to monitor apoptosis. Sorbonne-Paris-Nord University, France.
-
Moumaris M (2003) Biomedical research, the law of bioethics relating to the donation and use of elements and products of the human body. Paris-Descartes University, Faculté de Médecine, France.
-
Moumaris M, Abuaf N (2002) Use of labeled dextran for in-vitro assessment of increased cell permeability, cell death and apoptosis. Bulletin officiel de la propriété industrielle (Brevet n°00/09235) 2811682: A3.
-
Moumaris M, Benoliel S, Rouquette AM, Rajoely B, Abuaf N (2000) Phospholipid binding proteins on the plasma membrane of lymphocytes. J Autoimmun 15(2): 81-271.
-
Moumaris M, Ignoti S, Benoliel S, Oghina G, Rajoely B, et al. (1999) Characterization of B-cell adhering to the lamellar phospholipids. French Congress of Antiphospholipid Antibody, Paris, France.
-
Moumaris M (1996) Membranes érythrocytaires dans le paludisme: modèle d’étude: Souris- Plasmodium berghei anka. Université Pierre et Marie Curie, Faculté de Médecine, Paris, France.
-
Moumaris M, Sestier C, Miltgen F, Halbreitch A, Gentilini M, et al. (1995) Effect of Fatty Acid Treatment in Cerebral Malaria-Susceptible and Nonsusceptible Strains of Mice. The Journal of Parasitology 81(6): 997-999.
-
Sabolovic D, Moumaris M, Miltgen F, Sestier C, Halbreich A (1995) A subpopulation of red blood cells induced by bleeding or mosquito sucking. Chinese National Congress of Medical Biophysics, Shanghai, China.
-
Sabolovic D, Moumaris M, Miltgen F, Sestier C, Halbreich A (1995) Characterisation of subpopulation of red blood cells as a preferential target for malaria invasion. French Congress of Electrophoresis, Cell Electrophoresis, Pastor Institute, Paris, USA 19(7): 1215-1219.
-
Moumaris M (1994) Effet des acides gras sur la malaria cérébrale chez des souris susceptible et non susceptible. Université Paris XII, Faculté de Médecine, Créteil, France.
-
Moumaris M (1992) Lyme disease: Serological study. University of Orleans, France.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework