A Preliminary Study on the Potential Mechanism of Notch Signaling Pathway in the Regulation of Sudden Sensorineural Hearing Loss
Sudden sensorineural hearing loss (SSHNL) is a sudden onset of unexplained sensorineural hearing loss. Studies have shown that the number of patients with SSNHL is increasing year by year, and the average age is decreasing, which seriously affects the quality of life of patients. Its pathogenesis is complex and the mechanisms of hearing loss caused by various causes are quite different, but no matter what the cause is, it is ultimately related to the damage and loss of the inner ear structural hair cells (HCs) and their associated spiral ganglion cells (SGCs). Studies have shown that the Notch signaling pathway can regulate the development of the inner ear and nervous system in mammals. Hes family, Math1, DNER, Wnt signaling, and the lateral induction and lateral inhibition of Notch signaling related to the Notch signaling pathway are all key factors for the differentiation and development of HCs and SGCs. Based on the existing research, this paper makes a comprehensive and systematic summary, summarizes the importance of HCs and their afferent partner SGCs in SSNHL, and the relationship between Notch signaling pathway and the existence of the two, to provide a reference for clinical treatment of SSNHL in the future.
Congli Han1, Jiqin Tang*1, Jialin Gai1, Junfang Lei1, Jienuo Pan1, Jinchao Du2 and Bingshun Tang1
Introduction
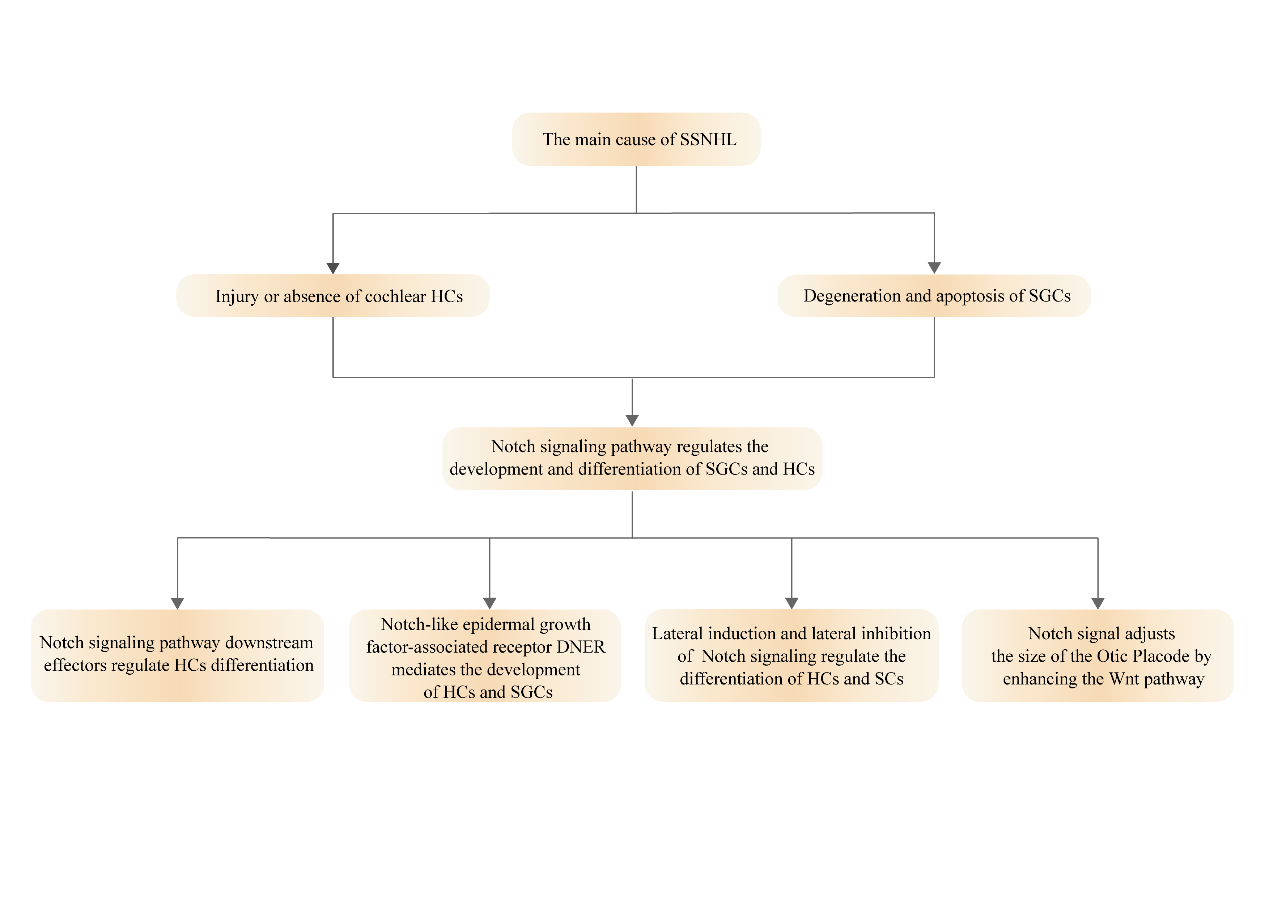
Sudden sensorineural hearing loss (SSNHL) is also known as idiopathic sensorineural hearing loss or Sudden Investigation Paper hearing loss, which belongs to the category of sensorineural hearing loss. SSNHL is most commonly defined as a hearing loss of 30dBHL or more on at least three consecutive audiometry frequencies occurring within 72 hours [1]. It is mainly manifested as hearing loss caused by sound perception and conduction disorders caused by hair cells (HCs) and/or spiral ganglion cells (SGCs) injury in the cochlea [2]. In the United States, there are at least 5 to 27 people diagnosed with SSNHL per 100,000 people, and about 66,000 new cases of SSNHL are reported every year. According to recent epidemiological surveys, the number of SSNHL patients is increasing year by year, and the average age has decreased [3]. Some patients have a poor prognosis, and may even cause secondary acoustic nerve injury. Hearing loss and accompanying symptoms will seriously affect their normal life and work. At present, the specific pathogenesis of SSNHL is still unclear, and no treatment can fully explain the pathogenesis of SSNHL. The most common suspected causes include viral infection, drug poisoning, blood circulation disorders, immune dysfunction, stress theory and so on. The mechanism of hearing loss caused by different etiologies varies greatly, but no matter which etiology, it is ultimately related to the damage or loss of HCs and their associated SGCs. Studies have found that Notch signaling pathway plays an important role in the regulation of the nervous system and the development of the mammalian cochlea [4], but there is no study on the regulation of SSNHL through Notch signaling pathway so far. Based on the existing studies, this paper comprehensively and systematically summarized the importance of HCs and its afferent partner SGCs in SSNHL, and summarized the relationship between Notch signaling pathway and the two, to provide new ideas for future clinical treatment of SSNHL. Figure 1 is the overall venation diagram of this article.
Materials and Methods
Inclusion Criteria
Published articles at home and abroad on SSNHL, Notch signaling pathway, HCs, and SGCs, including guidelines or consensus, reviews, clinical experiments, animal experiments, and retrospective studies.
Exclusion Criteria
(1)Articles with incomplete data or the full text of which cannot be obtained; (2) Articles published repeatedly; (3) Articles in languages other than Chinese and English.
Database and Search Strategies
Search CNKI and PubMed databases to collect articles on SSNHL, Notch signaling pathway, HCs, and SGCs published since the establishment of the database to March 2022. Search terms included: Sudden sensorineural hearing loss, Sudden Deafness, Notch, Hair Cells, Spiral Ganglion Cells, Reviews, Guidelines, Clinical Experiments, Animal Experiments, Retrospective Studies. Retrieving “fields” in different databases is adjusted.
Literature Screening
The EndNote X9 software was used for literature management, and two researchers independently screened the literature. In case of disagreement, the literature was discussed with a third party.
Literature Screening Process and Results
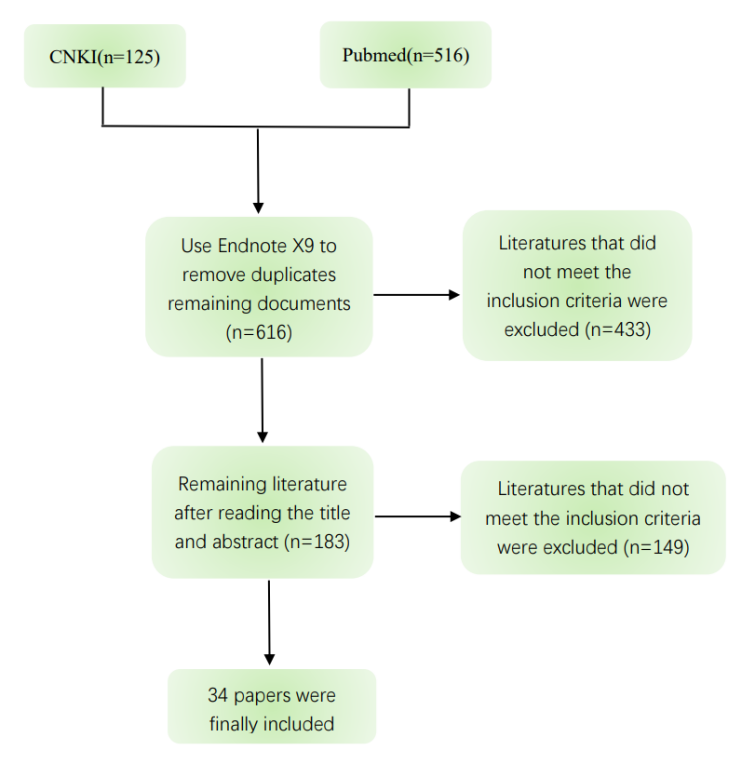
According to the preset search terms and inclusion and exclusion criteria, the literature was retrieved and screened, a total of 641 literatures were retrieved, and 616 literatures remained after excluding duplicate literatures. After further reading the relevant information of the title and abstract, the remaining literatures were 183 literatures, and finally included 34 articles. The retrieval process is shown in Figure 2.
The following is a comprehensive and systematic summary of the included literature, summarizing the importance of HCs and their afferent partner SGCs in SSNHL and the relationship between Notch signaling pathway and the two.
HCs and SGCs
HCs, which can convert sound into nerve impulses, are the primary receptors of auditory stimuli. There are two types of HCs and at least six associated non-sensory Sertoli cells (SCs) in the cochlear helix. The outer hair cells (OHCs), three rows of HCs located on the Outer side of the cochlea, are important for amplifying sound-induced vibrations. The first row of HCs located in the center or medial part of the cochlea is called inner hair cells (IHCs), which can contact afferent neuronal fibers and transmit sound signals to the brain [5]. The two kinds of HCs arranged in rows are separated from each other by SCs of different forms, forming a highly ordered asymmetric Mosaic pattern along the base of the cochlea to the apical axis, and constituting the sensory plaque of the inner ear [6]. Coyat C, et al. [7] demonstrated in their study on the morphological effects of the auditory injury on cochlear HCs that sustained noise exposure would cause loss of cochlear OHCs and fusion of IHCs stereocilia, thus leading to permanent hearing loss in the rat model.
SGCs are the first-level neurons of the auditory system, whose dendrites start from the base of helical HCs, and axons combine with HCs through banded synapses to transmit local electrical signals sent by HCs to secondary sensory neurons in the cochlear nucleus for sound processing [8]. There are at least two subtypes of type Ⅰ and type Ⅱ neurons in normal mammalian spiral ganglion neurons. Type Ⅰ SGCs, which account for about 95%, are bipolar cells connected to IHCs, which can encode sounds and transmit complex sound signals to the auditory brainstem for sound processing. Type Ⅱ SGCs, which account for about 5%, are bipolar or pseudobipolar cells connected to OHCs [9]. Ungar OJ, et al. [10] and colleagues selected the temporal bone histopathology bank cases that were diagnosed with unilateral SSNHL and did not recover to observe the decay rate of SGCs after SSNHL. The results showed that the number of SGCs had been in a declining state since the diagnosis of SSNHL, which was linearly proportional to the time of diagnosis.
HCs and their afferent chaperones SGCs are one of the major sensory cell types in the auditory system. After experiencing loud noise, certain drugs and aging wear and tear, some non-mammalian damaged HCs can proliferate and differentiate into new HCs through SCs, while mature mammalian HCs and SGCs are unable to regenerate, and severe loss of HCs can cause secondary damage to SGCs. Permanent hearing loss may eventually result [11, 12]. Otologists at home and abroad have conducted a large number of relevant studies to find ways to repair damaged and missing HCs or SGCs. Existing results show that inhibition of Notch signaling can force SCs to differentiate into HCs and increase the number of regenerated HCs [13]. In the cochlea and vestibular organs of embryonic or neonatal mice cultured in vitro, the use of drugs or γ-secretase inhibitors to interrupt the activation of Notch signaling can cause SCs to differentiate into HCs [14]. Other studies have shown that regenerated HCs can not only promote the growth of adult neurites but also re-establish connections with SGCs [15].
Notch Signaling Pathway
Notch signaling pathway is an evolutionarily highly conserved signaling pathway, which was first discovered in Drosophila in 1917 and has attracted widespread attention because of its important role in the development of biological embryos and cell life cycle [16]. This signaling pathway is composed of Notch receptors and ligands, CsL transcription factors, other effectors, and Notch regulatory molecules. Mammals have four Notch receptors (Notch1-4) and five ligands (Deltalike1,3,4, Jagged1 and Jagged2) [17, 18]. The receptor of Notch signal in the mammalian auditory receptive epithelium is mainly Notch l, and the ligands are mainly Deltalike1, Jagged1 and Jagged2 [11].
In the embryonic neural stem cells can express Notch receptors, and this phenomenon persists in the nervous system of adult animals, studies showed that Notch signaling pathways in the development of the nervous system plays a different role, including the proliferation of neural stem cells, cell fate specification and migration of neural stem cells, maintain differentiation and death [18, 19]. Secondly, Notch receptors bind to different ligands, participate in a series of morphological development of cells, play an important role in the activation of cell differentiation programs, promote or inhibit cell proliferation, embryonic and tissue development, tissue function and repair, and exist throughout the development of organisms [20, 21]. Thus, dysregulation or loss of Notch signaling may underlie a wide range of diseases from developmental syndromes to noncancerous and cancerous diseases in adults [22].
The transmission of Notch signaling does not depend on second messengers and protein kinases and can receive ligand binding from neighboring cells through intercellular contact and transfer it to the nucleus to activate relevant transcription factors [23]. When Notch receptors bind to ligands on neighboring cells, the receptors are broken down by ADAM family metalloproteinases (γ-secretase). The remaining transmembrane segments become substrates that are further cleaved by γ-secretase complexes, which release the intracellular domain (NICD) of Notch receptors, which then enter the nucleus and bind to DNA-binding proteins. Generate transcriptional complexes that activate downstream target genes, which affect gene transcription of repressor basic Helix-Loop-Helix (bHLH) gene subtypes such as Hes1 and Hes5.
Notch Signaling Pathway Regulates the Development and Differentiation of SGCs, HCs
• Notch signaling pathway downstream effectors regulate HCs differentiation The Hes family, a downstream effector of the Notch signaling pathway, is one of the key genes controlling the differentiation of HCs in the inner ear, and its expression is upregulated by activation of the Notch signaling pathway. The proto-neural gene Math1 is also expressed in the inner ear and encodes a transcription factor that is important in the differentiation and maturation of HCs, the deletion of which leads to disruption of HCs development. In postnatal rat explant culture experiments, it was found that the differentiation of HCs induced by Math1 was inhibited by Hes1 and Hes5, both of which negatively regulate HCs differentiation by antagonizing Math1 [24]. It has been shown that the Hes binding site at the Math1 promoter inhibits Math1 expression and reduces differentiation toward hair cells, and when this inhibition is lifted, it stimulates Math1 expression [25, 26]. In addition, two repressive bHLH proteins encoded by Hes1 and Hes5 inhibit the overexpression of the pre-HCs gene Atoh1 upon differentiation of SCs, thereby inhibiting the differentiation of SCs into new HCs [27]. Thus, the generation of HCs can be achieved by repressing the transcription of Notch signaling.
• Notch-like epidermal growth factor (EGF)-associated receptor DNER mediates the development of HCs and SGCs DNER, a mammalian Delta/Notch-like epidermal growth factor (EGF)-associated receptor, is a neuron-specific transmembrane protein with 10 extracellular EGF-like structural thresholds. These structural domains have high similarity to the Notch receptor and ligand Delta-like and can bind to Notch1 receptors at cell phase contacts to activate Notch signaling [28]. In previous studies, it was known that DNER mediates the development of HCs and that it plays a key role in the development of the central nervous system and seems to be involved in the establishment of neuronal polarity and neurosynaptic extension processes through the Notch signaling pathway [29]. It has been shown that DNER is robustly expressed in type I and type II SGCs and HCs during cochlear embryonic development and that this expression can be maintained into adulthood [30]. One experiment performed double immunolabeling experiments in a rat model using three markers of the anti-DNER and SGCs groups and found that all SGCs bodies in the adult mouse cochlea were labeled with DNER [31]. Future studies on the promotion or inhibition of DNER expression directed to the generation of HCs and SGCs are needed through numerous experiments.
• Lateral induction and lateral inhibition of Notch signaling regulate the differentiation of HCs and SCs The Notch signaling pathway is characterized by lateral induction and lateral repression. In lateral inhibition, ligand- producing cells inhibit neighboring cells from becoming the same cell type to reduce ligand expression, whereas lateral induction is a positive feedback loop in which ligand- expressing cells induce their neighbors to increase ligand expression, thereby promoting Notch signaling activation and coordinating various cellular behaviors [32]. During neuronal generation in the mammalian nervous system, there is an expression of a family of bHLH transcription factors based on the proto-neural genes encoded by the bHLH transcription factor family, which is controlled by a mechanism of lateral inhibition and feedback mediated by the Notch ligand Deltalike1 [33]. Notch signaling through lateral inhibition forms a mosaic of HCs and SCs arranged pattern, which is considered to be one of the most prominent functions of Notch signaling. Lateral induction plays a major role in the early stage of differentiation of HCs and SCs, maintaining the cells in a pre-sensory state so that they can readily differentiate into HCs and SCs, whereas lateral inhibition plays a major role in the late stage of differentiation of HCs and SCs, limiting the proportion of pre-sensory cells that continue to differentiate into HCs and SCs.
• Notch signal adjusts the size of the Otic Placode by enhancing the Wnt pathway The main sensory organs of vertebrates, such as vision, hearing, taste and olfactory system, were all developed from the placode or play a certain role in these sensory organs. Otic placode (OP) was derived from the oval thickened tissue of the ectoderm around the neural plate, namely the placode precursor, which generated the body’s complex sensory organ spiral apparatus, as well as cells such as HCs, SCs and sensory neurons, thereby transforming sound into hearing [34]. From the origin of OP to the formation of mature mechanical sensory structure, the Notch signal pathway was involved in the development of cochlea [35].
OP, extrabranchial basal plate and cranial epidermis form the preauricular area. Studies have found that Notch signal can affect the size of OP by regulating the proliferation of preauricular progenitor cells. In further bird embryo animal model experiments, it was observed that Notch signaling pathway may play a role after preauricular area induction and before morphological basal plate formation [36]. In the study, Ohyama, T. and his colleagues found that ectoderm was expressed by Pax2+, a member of the family of transcription factors related to evolution. Precursor cells were induced to differentiate into OP epithelium, cranial epidermis and extrabranchial basal plate by Pax2+. The activation of Wnt signaling pathway could guide Pax2+ to differentiate into OP rather than cranial epidermis [37]. The activation of Notch1 receptor in the precursor of Pax2+ expands the size of OP epithelium at the cost of reducing the expression of the cranial epidermis, and the loss of Notch1 receptor will lead to a decrease in OP size [38]. In this process, Notch signal can positively regulate the Wnt signaling pathway to specify the size of OP.
Conclusion
SSNHL is mainly due to the irreversible damage of cochlear HCs caused by various reasons, which results in degeneration and apoptosis of SGCs. The age of onset ranges from children to the elderly, and the specific pathogenesis is unknown. It is often accompanied by symptoms such as dizziness and tinnitus. The study of its pathogenesis and treatment methods has long been an important topic at home and abroad. The inner ear of the human newborn is fully developed and mature. The lack of cell proliferation in the damaged adult inner ear is a major obstacle to the regeneration of inner ear cells. Therefore, the key to adult inner ear cells after the injury is their re-proliferation and/ or differentiation and regeneration.
Previous studies have shown that Notch signaling pathway plays an important role in the development, injury and repair of the inner ear [39]. This study summarized the role of Notch signaling pathway in the development and differentiation of the inner ear by consulting the relevant literature at home and abroad. Notch signal can expand the size of the ear basal plate by enhancing Wnt pathway, and its lateral induction and lateral inhibition can regulate the differentiation of HCs and SCs. Notch signal target genes such as Hes1 and Hes5 combine with Math1 to negatively regulate the differentiation of HCs. In addition, Notch EGF-related receptor DNER is also steadily expressed in the development of HCs and SCs, and the specific regulatory mechanism needs to be further studied. At present, there is no research to verify the regulation mechanism of Notch signaling pathway in SSNHL-related disease model, so it is necessary to further establish an animal model, and on this basis, clarify the mechanism of Notch signal pathway in the SSNHL model, to establish a systematic relationship between Notch signaling regulation and SSNHL mechanism, and provide a more reliable theoretical basis for the clinical treatment of SSNHL.
Conflict of Interest Statement
The authors have no conflicts of interests to disclose.
Author Contributions
Jiqin Tang conceived and designed the research as corresponding authors . Congli Han performed the main manuscript text. Jialin Gai was responsible for literature retrieval. Junfang Lei and Jienuo Pan were responsible for the classification and sorting of the included literature. Jinchao Du was responsible for verifying the authenticity of the retrieval steps and contents. Bingshun Tang made Figure 1 and Figure 2, and revised the full text. All authors reviewed the manuscript.
Funded Projects
- Evaluation of the mechanism and efficacy of the first batch of classical prescriptions in the treatment of major diseases in Shandong University of traditional Chinese Medicine of scientific research innovation team fund- funded projects (220316)
- Development of Medical and Health Science and Technology in Shandong Province (2018WS205)
- The Development Program of traditional Chinese Medicine Science and Technology in Shandong Province
(2017-011).
References
-
Chandrasekhar SS, Tsai DO BS, Schwartz SR (2019) Clinical Practice Guideline: Sudden Hearing Loss (Update). J Otolaryngol Head Neck Surg 161(s1): S1-s45.
-
Haijun C, Zongrun S, Yan G (2021) Advances in the molecular mechanism of sensorineural hearing loss. J Journal of Anatomy 44(06): 526-529.
-
Chen X, Fu YY, Zhang TY (2019) Role of viral infection in sudden hearing loss. J The Journal of international medical research 47(7): 2865-2872.
-
Samarajeewa A, Jacques BE, Dabdoub A (2019) Therapeutic Potential of Wnt and Notch Signaling and Epigenetic Regulation in Mammalian Sensory Hair Cell Regeneration. J Molecular therapy: the journal of the American Society of Gene Therapy 27(5): 904-911.
-
Zheng W, Holt JR (2021) The Mechanosensory Transduction Machinery in Inner Ear Hair Cells. J Annual review of biophysics 50: 31-51.
-
Driver EC, Kelley MW (2020) Development of the cochlea. J Development (Cambridge, England) 147(12).
-
Coyat C, Cazevieille C, Baudoux V (2019) Morphological consequences of acoustic trauma on cochlear hair cells and the auditory nerve. J The International journal of neuroscience 129(6): 580-587.
-
Sun S, Babola T, Pregernig G (2018) Hair Cell Mechanotransduction Regulates Spontaneous Activity and Spiral Ganglion Subtype Specification in the Auditory System. J Cell 174(5): 1247-63.e15.
-
Shanshan L, Qixuan W, Zhiwu H (2018) Single-cell sequencing found that the activity of inner hair cells induces type I spiral ganglia to be divided into three types. J Journal of Audiology and Speech Diseases 26(6): 685-688.
-
Ungar OJ, Handzel O, Santos F (2018) Rate of Spiral Ganglion Cell Loss in Idiopathic Sudden Sensorineural Hearing Loss. J Otology & neurotology 39(10): e944-e949.
-
Lihong X, Menglin W, Anzhou T (2018) Research progress of inner ear hair cell regeneration and its signaling pathway. J Journal of Audiology and Speech Diseases 26(05): 547-550.
-
Wagner EL, Shin JB (2019) Mechanisms of Hair Cell Damage and Repair. J Trends in neurosciences 42(6): 414-424.
-
Mcgovern MM, Zhou L, Randle MR (2018) Spontaneous Hair Cell Regeneration Is Prevented by Increased Notch Signaling in Supporting Cells. J Frontiers in cellular neuroscience 12: 120.
-
Yamamoto N, Tanigaki K, Tsuji M (2006) Inhibition of Notch/RBP-J signaling induces hair cell formation in neonate mouse cochleas. J Journal of molecular medicine (Berlin, Germany) 84(1): 37-45.
-
Shu Y, Li W, Huang M (2021) Renewed proliferation in adult mouse cochlea and regeneration of hair cells. J Nature communications 10(1): 5530.
-
Xi Y, Peng C, Xia J (2021) Research progress of Notch signaling pathway in insects. J Acta Entomology 64(02): 250-258.
-
Pancewicz J, Niklinska W, Eljaszewicz A (2022) Anti- Jagged-1 immunotherapy in cancer. J Advances in medical sciences 67(2): 196-202.
-
SAlazar JL, Yang SA, Yamamoto S (2020) Post- Developmental Roles of Notch Signaling in the Nervous System. J Biomolecules 10(7).
-
Engler A, Zhang R, Taylor V (2018) Notch and Neurogenesis. J Advances in experimental medicine and biology 1066: 223-234.
-
Yujie L, Hong L, Hongyu L (2022) Study on the relationship between NOTCH2NLC gene and nervous system diseases. J Chinese Journal of Practical Neurological Diseases 25(01): 128-132.
-
Xiaoting H, Yuzhu L, Changqian Z (2020) Notch Signaling Pathway and Neurodegenerative Diseases. J Chinese Medicine Guide 18(12): 33-34.
-
Zhou B, Lin W, Long Y (2022) Notch signaling pathway: architecture, disease, and therapeutics. J Signal transduction and targeted therapy 7(1): 95.
-
Yangyang W, Yi Z, Jing L (2021) Advances in the effects of Notch signaling on inner ear hair cell regeneration. J Chinese Journal of Ophthalmology and Otolaryngology 21(04): 309-311.
-
Zheng JL, Shou J, Guillemot F (2000) Hes1 is a negative regulator of inner ear hair cell differentiation. J Development (Cambridge, England) 127(21): 4551- 4560.
-
Ran F, Feilong M, Gaopeng L (2019) Functions of Math1 in the inner ear hair cell development and regeneration. J Journal of Zhejiang University (Science Edition) 46(01): 78-91.
-
Abdolazimi Y, Stojanova Z, Segil N (2016) Selection of cell fate in the organ of Corti involves the integration of Hes/Hey signaling at the Atoh1 promoter. J Development (Cambridge, England) 143(5): 841-850.
-
Du X, Li W, Gao X (2013) Regeneration of mammalian cochlear and vestibular hair cells through Hes1/Hes5 modulation with siRNA. J Hearing research 304: 91-110.
-
Wang Z, Li Z, Wu Q (2020) DNER promotes epithelial- mesenchymal transition and prevents chemosensitivity through the Wnt/β-catenin pathway in breast cancer. J Cell death & disease 11(8): 642.
-
Hsieh FY, Ma TL, Shih HY (2013) Dner inhibits neural progenitor proliferation and induces neuronal and glial differentiation in zebrafish. J Developmental biology 375(1): 1-12.
-
Du J, Wang X, Zhang X (2018) DNER modulates the length, polarity and synaptogenesis of spiral ganglion neurons via the Notch signaling pathway. J Molecular medicine reports 17(2): 2357-2365.
-
Hartman BH, Nelson BR, Reh TA (2010) Delta/notch- like EGF-related receptor (DNER) is expressed in hair cells and neurons in the developing and adult mouse inner ear. J Journal of the Association for Research in Otolaryngology JARO 11(2): 187-201.
-
Brown RM, Nelson JC, Zhang H (2020) Notch-mediated lateral induction is necessary to maintain vestibular prosensory identity during inner ear development. J Developmental biology 462(1): 74-84.
-
Kageyama R, Ohtsuka T, Shimojo H (2008) Dynamic Notch signaling in neural progenitor cells and a revised view of lateral inhibition. J Nature neuroscience 11(11): 1247-1251.
-
Tambalo M, Anwar M, Ahmed M (2020) Enhancer activation by FGF signalling during otic induction. J Developmental biology 457(1): 69-82.
-
Żak M, Daudet N (2021) A gradient of Wnt activity positions the neurosensory domains of the inner ear. J eLife 10: e59540.
-
Shida H, Mende M, Takano-yamamoto T (2015) Otic placode cell specification and proliferation are regulated by Notch signaling in avian development. J Developmental dynamics : an official publication of the American Association of Anatomists 244(7): 839-851.
-
Ohyama T, Groves AK (2004) Generation of Pax2- Cre mice by modification of a Pax2 bacterial artificial chromosome. J Genesis (New York, NY :2000) 38(4): 195-199.
-
Jayasena CS, Ohyama T, Segil N (2008) Notch signaling augments the canonical Wnt pathway to specify the size of the otic placode. J Development (Cambridge, England) 135(13): 2251-2261.
-
Huang J, Sun X, Wang H (2022) Conditional overexpression of neuritin in supporting cells (SCs) mitigates hair cell (HC) damage and induces HC regeneration in the adult mouse cochlea after drug-induced ototoxicity. J Hearing research 420: 108515.
- 4th Branchial Cleft Sinus Anomaly Presenting as Recurrent Thyroid Abscess in A Child: A Case Report
- Parotid Duct Injury Repaired Using an Angiocatheter Stent: A Case Report
- Organization and Functionality of the Referral and Counter-Referral System for ENT Disorders in District Hospitals of N’Djamena, Chad: A Cross-Sectional Analytical Study
- Facial Metastases from a Gastrointestinal Stromal Tumor: A Case Report
- Panorama of Ent Cancers and Literature Review: Epidemiological Profile and Therapeutic Management
- Could Antimicrobial Resistance Prove to Be Both a Threat and an Opportunity for us?