Nucleolar Organizer Region (NOR) Polymorphism in Relation to Interspecific and Intraspecific Variability in Some Indian Anurans-A Review
The amphibians being originated from Crossopterygian fishes in the late Devonian constitute one of the most interesting groups of vertebrates. Indian subcontinent is rich in amphibian fauna but the cytogenetical studies in this field are scanty. The present review has been oriented to analyse the nucleolar organizer region polymorphism in some Indian anuran bufonids, ranids and rhacophorid. Conventional staining of mitotic metaphase stages revealed that three Bufonid species studied possess symmetrical karyotypes with 2n=22 and NF=44. The four ranid species and a single sp. of Rhacophorus studied possess symmetrical karyotypes with 2n=26 and NF=52. Nucleolar organizer region or secondary constriction (SC) region staining by HCl-trypsin digestion and silver impregnation technique revealed that the number and position of NORs are variable not only among the different species but also in different individuals of the same species. Considerable variation in the length of NOR or SC has been noted in different specimens of the same species. An uneven distribution of NOR has also been found in Bufo stomaticus where males possess one NOR bearing chromosome in the chromosome pair # 5 and female with two NOR bearing chromosome that is in the chromosome pair # 7. The NOR polymorphism in relation to interspecific and intraspecific variability in the size and number is interesting which would be of cytotaxonomic value to trace the phylogenetic relationship among the different species.
Introduction
The nucleolus organizer regions (NORs) or Secondary constrictions (SCs), have been recognized as sites containing ribosomal DNA (rDNA) cistrons [1, 2]. Biochemical studies have shown that the silver stained parts of chromosomes consist of acidic proteins [3]. The NORs constitute important landmarks in identifying a particular chromosome or chromosomal segment in the karyotype of a species [4, 5, 6]. The NOR polymorphism seems to be a general phenomenon in different vertebrates such as fishes [7, 8, 9], amphibians [6, 10, 11, 12] and reptiles [13]. In conventional stained metaphases, the precise identification of the NOR is difficult due to extreme condensation of chromatids. The silver staining method is often used to demonstrate the position of the NOR in the chromosome [14].
Amphibians, the amazing group of vertebrates, have been the subject of various cytogenetic investigations [4, 10, 15, 16, 17] because of their transitional phylogenetic position in the animal kingdom, low diploid numbers with enormous in length and ambiguous nature of sex chromosome. There are 7350 amphibians in the world out of which 6458 are anurans [18]. Infact, very little has been added to the cytogenetics of Amphibia distributed in Indian subcontinent. The number and location of NORs have been studied in a number of Anuran species [6, 19] to solve various phylogenetic riddles in amphibian cytotaxonomy. The karyotypes of Bufo andersonii and B.melanostictus (=Duttaphrynus melanostictus ) are almost identical, differing only in the presence of a prominent secondary constriction in the short arm of both the chromosomes of pair number 7 in the former and its complete absence in the latter [20]. In majority of the Anuran species investigated so far, the NORs occupy an interstitial or subterminal position in the chromosome arms. It was also noticed that the size of Ag-stained NORs is extremely variable in different or closely related species [6, 10]. The NORs in the chromosomes of Pyxicephalus adspersus revealed a high frequency of inter-individual variations (duplications or triplications etc.). The high degree of interindividual variability in respect to size and number of AgNO3 stained NOR seems to be a characteristic feature in different groups of vertebrates such as fishes [21] , amphibians [4,5,10 ] and mammals [14, 22]. Moreover, the number and position of NOR have been used as markers in karyosystematic studies of Anurans (5,6). Present review has, therefore, been oriented to analyze the chromosomal morphology of some Indian Anurans with respect to Nucleolar organizer region polymorphism that not only add further cytological data on Amphibian karyosystematics but also readjust and refine the existing knowledge gathered in this field of special interest.
Distribution of Anuran Amphibians in Some Localities of West Bengal, India
The review work of NOR polymorphism has been oriented on the basis of some cytological observations [10, 19] of some anuran species (Table 1 & Figure 1).
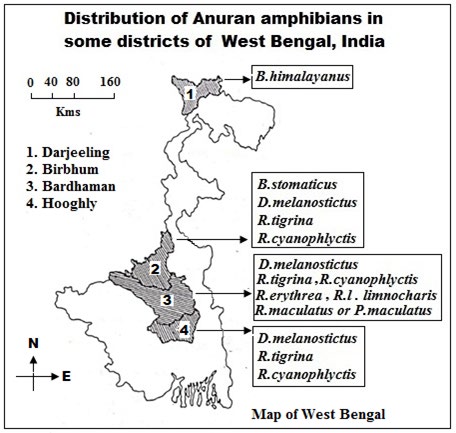
| Order | Family | Scientific name | Distribution |
|---|---|---|---|
| Anura | Bufonidae | Bufo melanostictus(=Duttaphrynus melanostictus | Bardhaman, Birbhum, Hooghly districts, West Bengal, India |
| B. himalayanus | Darjeeling District, West Bengal, India | ||
| B. stomaticus | Bakreshwar, Birbhum District, West Bengal, India | ||
| Ranidae | Rana tigrina | Bardhaman, Birbhum, Hooghly districts, West Bengal, India | |
| R.cyanophlyctis | Bardhaman,Birbhum and Hooghly districts, West Bengal, India | ||
| R. erythrea | Bardhaman district, West Bengal, India | ||
| R. limnocharis limnocharis | Bardhaman district, West Bengal, India | ||
| Rhacophoridae | Rhacophorus (Polypedates) maculatus | Bardhaman district, West Bengal, India |
Table: 1: Systematic position and related particulars of eight anuran species.
HCl-Trypsin Digestion Technique to Detect Satellite and Satellite Stalk
Since amphibian chromosomes remain in a highly condensed state at metaphase (Figure 2a,b) it becomes difficult to detect NOR or satellite stalk and satellite chromosomes. In order to expose the NOR or satellite region the HCl-Trypsin digestion technique was standardized [19].
NOR banding_:_ N-band was performed earlier [10] by slight modifications of the technique described by Howell and Black [23].
Conventional Stained Mitotic Chromosomes of Bufo and Rana
Conventional staining of mitotic chromosomes of the species studied under genus Bufo: B. melanostictus, B.himalayanus and B.stomaticus revealed an identical diploid number (2n=22 and fundamental number = 44) with a symmetrical karyotype [10].
Conventional staining of mitotic chromosomes revealed that 2n number of all the species studied under the genus Rana and Rhacophorus is 26 with NF=52 (Figure 2). The karyotype is typically symmetrical with all biarmed chromosomes. No morphologically distinguishable heteromorphic sex chromosome pair could be identified in conventional Giemsa stained preparations [10].
Both NOR and satellite bearing chromosomes constitute important chromosome landmarks and provide important clues in tracing karyological evolutionary trend in anurans. The metaphases of Duttaphrynus melanostictus and B. himalayanus subjected to subsequent acid and enzyme treatment showed one pair chromosome with distinct satellite and satellite-stalk [10, 19]. In Duttaphrynus melanostictus it is located in the long arm of chromosome pair # 11 where as in B. himalayanus it is located in the short arm of chromosome pair # 11 (Figure 3a,b). In female specimens the NOR is prominent in both the homologues whereas in male one chromosome with well-developed NOR and other one of this pair is small. In B. stomaticus the SC or NOR region is visualized in one chromosome of pair # 5 in male and both the homologues of the pair # 7 in case of female individuals (3c,d). The application of HCl-Trypsin digestion method in all the species studied under the genus Rana and Rhacophorus revealed that one pair of chromosomes always showed secondary constriction region. It is located at the middle region of its long arm of 7th pair in Rana tigrina, proximal region of the short arm of the 8th pair in R. cyanophlyctis, short arm of 8th pair in R. erythrea and short arm of 7th pair in R. limnocharis limnocharis. In case of Rhacophorus maculatus the secondary constriction is visualised in the short arm of 11th pair (Figure 4c-g)
Visualisation of NOR by AgNO3 staining
In metaphase chromosomes stained with Ag, the dark Ag label is clearly localised exactly within the nucleolar constriction of the light brown stained chromosomes (Figure 4a,b). The Ag-banding coincided exactly within the chromosomal secondary constrictions detected earlier by the application of HCl Trypsin digestion technique. Due to the extreme length of the NORs, the terminal region of the NOR carrying chromosome arms appear as satellites in Duttaphrynus (Bufo) melanostictus. A comparison of the NOR carrying chromosomes of different individuals of the same species revealed considerable differences in the lengths of the NOR (Figure 3e-j). Moreover, variation in respect to distribution of NOR in chromosome pair is sex specific. The HCl Trypsin digestion banding patterns showed that NOR in the middle region of the short arm of both the chromosome pair 7 in the female individuals whereas in the male it was recorded interstitially in the long arm of a single chromosome no.5 (Figure 3c,d).
It is evident from the existing knowledge [6] and also from the present study [10, 24] that the difference in NOR size in Anura could either be due to deletion or duplication. This is a good example of NOR heteromorphism. Interestingly, variations in number and size of the Ag-stained nucleoli were also recorded in the diploid interphase nuclei (Figure 4a).
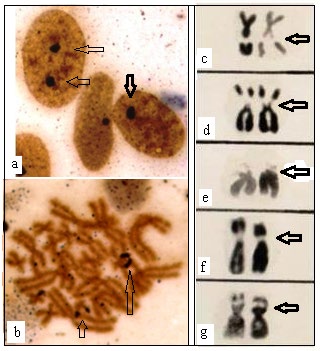
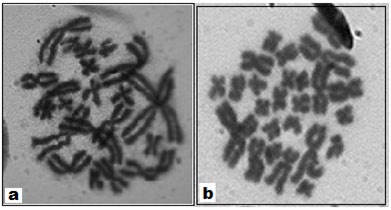
Figure 3: a-j: NOR detection by Silver staining and HCl- Trypsin digestion method (Individual chromosomes are arranged from different photographic plates showing less extended NOR and highly extended NOR) a = Silver stained NOR in Bufo melanostictus; b = HCl_-Trypsin treated Giemsa stained NOR in _B. himalayanus. c = HCl -Trypsin treated Giemsa stained NOR in B. stomaticus (male) with one NOR. d = HCl – Trypsin treated Giemsa stained NOR in B. stomaticus (female) with two NOR. e, f = HCl_-Trypsin treated Giemsa stained NOR in _B. melanostictus . NOR is less extended to highly extended state. g, h, i, j = Silver stained NOR in B. melanostictus . NOR is less extended to highly extended state ( (Modified after Banerjee,1986; Banerjee and Chakrabarti, 1986; 1987).
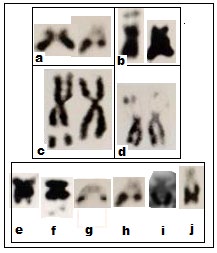
Figure 4: a-g: a = Silver stained cells in Bufo melanostictus (B.m). Variations in number of the Ag-stained nucleoli in interphase cells (arrowed); b = Silver stained metaphase with two NOR bearing chromosomes in B.m. Interestingly, NOR is extended (large arrow) in one chromosome of the homologue but other is dotted (small arrow). NOR is exposed (arrowed) by HCl-trypsin digestion method in Rana tigrina (c); R. cyanophlyctis (d); R. erythrea (e); R. limnocharis limnocharis (f) and Rhacophorus maculatus (g) (Modified after Banerjee, 1986; Banerjee and Chakrabarti, 1986)
Conclusion
Interspecific and intraspecific polymorphism in NORs has been demonstrated in different groups of vertebrates [6, 7, 8, 9, 10, 11, 12, 13]. Most of these variations are related to the chromosome specific location of NOR, number of active NOR per cell and relative size of specific NOR on chromosome.
In the present investigations and data from previous works [10, 19, 24, 25] all the species belonging to the genus Bufo, Rana and Rhacophorus were found to possess a single pair of SC or NOR distributed interstitially with some exceptions. A species specificity in distribution of SC / NOR has been noted. Because in Rana tigrina the SC is located in the long arm of chromosome # 7, in R. cyanophlyctis in the short arm of chromosome # 8, in R.erythrea in the short arm of chromosome # 8, , in R. limnocharis limnocharis in the short arm of chromosome # 7 and in Rhacophorus maculatus in the short arm of chromosome # 11 ( Figure 4c-g). In Duttaphrynus melanostictus it is located in the long arm of chromosome # 11 and in B.himalayanus it is located in the short arm of chromosome # 11(Figure 3a,b). Earlier Schmid [6] identified species specific distribution of NOR in different anurans. Interestingly, in the present study an apparent sex specific difference in the number of NOR or SC has been found in B. stomaticus in which male is heteromorphic in having a single NOR bearing chromosome and the females two. This situation receives further attention because in male the NOR is localised in the long arm of chromosome # 5 where as in female the NOR is localised in the short arm of chromosome # 7 (3c,d). This uneven localisation of NOR in male and female karyotypes casts doubt on the possibility of sex specific NOR polymorphism in this species. Instead it recalls the possibility of NOR polymorphism resulted due to breakage and fusion of or near nucleolar organizer regions. Some authors [26, 27] suggested that unequal meiotic crossovers or sister chromatid exchanges are the probable causes for polymorphism in NOR distribution.
In the present investigation, intraindividual size differences of NOR were frequently noted in Duttaphrynus melanostictus and B.himalayanus. It is evident from the existing knowledge and also from the present study that the difference in NOR size in Anura could either be due to deletion or duplication. Experiment conducted on Ambystoma mexicanum [28] showed pronounced differences in the length of nucleolar constriction are directly related to differences in the number of rRNA cistrons. Miller, et al. [22] advocated that the Ag-staining in the mitotic metaphase chromosomes demonstrates only those NORs which were active i. e, engaged in ribosomal RNA synthesis in the preceding interphase. Lack of Ag-staining on the chromosome arm indicates that either this NOR was inactive in the preceding interphase, or the NOR was completely deleted and lost. Lopez et al. [29] have stated that the size variation between the two NOR bearing homologs should be more functional than structural. Therefore, the present review indicates that in Duttaphrynus melanostictus, the large duplicated or triplicated Ag blocks are localised in the very extended nucleolar constrictions and it may be assumed that the number of the rRNA cistrons have greatly increased with a corresponding increase in ribosomal RNA synthesis at these loci.
It is evident from present analysis and previous study [10, 19, 24, 25] that all five species viz. Rana tigrina, R.cyanophlyctis , R. erythrea, R. limnocharis limnocharis and Rhacophorus maculatus or Polypedates maculatus show unique resemblance in their identical chromosome number ( 2n = 26) and NOR number (one pair) with some deviations exhibited mainly in the location of NOR in different pair of chromosomes. In addition, chromosomal features analysed here indicate that Duttaphrynus melanostictus, Bufo himalayanus and B. stomaticus possess an identical chromosome number (2n=22) and NOR number with minor differences with respect to NOR distribution in different chromosome. Considering various information about the different species studied under the genus Rana, Rhacophorus and Bufo or Duttaphrynus indicated that they are to some extent conservative with respect to chromosome number and NOR number with little deviations exhibited in the location of NOR. Perhaps new approaches in light of banding technology in this field will focus new information about the anuran cytotaxonomy.
Acknowledgement
Acknowledgement is due to University Grants Commission (Grant no- F/PSW–080/09–10 dated 26-10- 2009), Eastern Regional Office, Salt Lake, Kolkata, India for financial support. The paper is dedicated to my teacher and research guide Late Prof. Samar Chakrabarti, Cancer Cytogenetics Unit, Department of Zoology, Burdwan University, Burdwan, India. Author is grateful to Late Dr. N. M. Bose, Former Emeritus Lecturer, Department of Zoology, Rammohan College, Kolkata, India. Author is grateful to Samar Bejoy Das for help. Author is also thankful to Principal, Dr. Saswati Sanyal, Rammohan College, Kolkata, India for support and encouragement. References
1. Pardue ML, Hsu TC (1975) Locations of 18s and 28s ribosomal genes in the chromosomes of the Indian Muntjac. Journal of Cell Biology 64: 251-254.
2. Ghosh S (1976) The nucleolar structure. International Review of Cytology 44: 1-24.
3. Sumner AT (1990) Chromosome banding. Publication academic Division of Unwin Hymen Ltd, London.
4. Schmid M (1978) Chromosome banding in Amphibia. I Constitutive heterochromatin and nucleolar organizer regions in Bufo and Hyla. Chromosoma 66: 361-388.
5. Schmid M (1980) Chromosome banding in Amphibia V. Highly differentiated ZW/ZZ sex chromosomes and exceptional genome size in Pyxicephalus adspersus (Anura, Ranidae) Chromosoma 80: 69-96.
6. Schmid M (1982) Chromosome banding in Amphibia VII. Analysis of the structure and variability of NORs in Anura. Chromosoma 87: 327-344.
7. Phillips R, Zajicek KD, Utter FM (1986) Chromosome banding in salmonid fishes: nucleolar organizer regions in Oncorrynchus. Canadian Journal of Genetics and Cytology 28: 502-510.
8. Sanchez L, Martinez P, Vinas A, Bouza C (1990) Analysis of the structure and of nucleolar organizer regions of Salmo trutta by C-, Ag-, and restriction endonuclease banding. Cytogenetics Cell Genetics 54(1-2): 6-9.
9. Carvalho R, Dias AL (2007) Interindividual size heteromorphism of NOR and chromosomal location of 5s rRNA genes in Iheringichthys labrosus. Brazilian Archives of Biology and Technology 50(1): 141-146.
10. Banerjee SN (1986) Chromosomal endophenotype of some Indian Anura with reference to C- band distribution and sensitivity of heterochromatin to induced aberration. Ph.D. Thesis, University of Burdwan, Burdwan, West Bengal, India, pp : 1-149.
11. Silva APZ, Haddad CFB, Galassi GG, Kasahara S (2006) Multiple NORs in Leptodactylus mystacinus ( Amphibia, Anura) and comments on its systematic position in the L. fuscus group based on cytogenetic and molecular analyses. Genetica 127(1-3): 35-44.
12. Quindere YR, Lourenco LB, Andrade GV, Tomatis C, Baldo D, et al. (2009) Polytypic and polymorphic cytogenetic variations in the widespread anuran Physalaemus cuvieri (Anura, Leiuperidae) with emphasis on nucleolar organizing regions. Biological Research 42(1): 79-92.
13. Bickham J W, Rogers DS (1985) Structure and variation of the nucleolus organizer regions turtles. Genetica 67: 171-184.
14. Goodpasture C, Bloom SE (1975) Visualization of NORs in mammalian chromosomes using silver staining. Chromosoma 53(1): 37-50.
15. Morescalchi A (1979) New developments in vertebrate cytotaxonomy I. Cytotaxonomy of the amphibians. Genetica 50(3): 179-193.
16. Chakrabarti S (1979) Somatic chromosomes of Indian burrowing frog Uperodon globulosum (Gunther Anura: Amphibia). Experientia 35: 743-744.
17. Manna GK (1983) Cytogenetic studies on fish and amphibian.Conference Proceedings, Genetical Research in India, XV International Genetics Congress, New Delhi, pp: 243-273.
18. Frost DR (2011) Amphibian species of the world: an Online Reference. New York, USA.
19. Banerjee SN, Chakrabarti S (1987) A simple technique to detect satellite and satellite stalk in amphibian. Current Science 56: 41-42.
20. Singh L (1974) Present status of sex chromosomes in amphibians. The Nucleus 17: 17-27.
21. Foresti F, Almeida Toledo LF, Toledo FSA (1981) Polymorphic nature of nucleolus organizer regions in fishes. Cytogenetics Cell Genetics 31: 137-144.
22. Miller DA, Dev VG, Tantravahi R, Miller OJ (1977) Frequency of satellite association of human chromosomes is correlated with amount of Ag-staining of nucleolus organizer regions. American Journal Human Genetics 29: 490-502.
23. Howell WM, Black A (1980) Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: a I-step method. Experientia 36: 1014-1015.
24. Banerjee SN (2015) Nucleolar Organizer Region (NOR) Polymorphism in relation to interspecific and intraspecific variability in some Indian Anurans. Invited talk delivered at Kasetsart University, Bangkok, Thailand, 5th Asian Chromosome Colloquium pp: 53.
25. Banerjee SN, Chakrabarti S (1986) The present status of sex chromosomes in some Indian Anurans. In: Manna GK, Sinha U, et al. (Eds.), Perspectives in Cytology and Genetics 5: 119-124.
26. Gold JR (1984) Silver staining and heteromorphism of chromosomal nucleolus organizer regions in North American Cyprinid fishes. Copeia 1: 133-139.
27. Castro J, Sanchez L, Martinez P (1998) Analysis of the inheritance of NOR size variants in brown trout (Salmo trutta). Journal of Heredity 89: 264-266.
28. Sinclair JH, Carrol CR, Humphrey RR (1974) Variation in rDNA NOR length in normal and variant lines of the Mexican axolotl. Journal of Cell Science 15: 239-257.
29. Lopez JR, Alvarez MC, Thode G, Martinez G (1989) Karyotype divergence in Symphodus melops and S. roissali (Labridae, Perciforms): C- banded and Ag-NOR karyotypes. Genome 32: 35-39.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework