Dynamics of Fluctuations in the Indices of Species Diversity of Ichthyoplankton in the Northern Part of the Central-Eastern Atlantic
An integrated indicator of the condition of any community can be its species diversity, expressed by various indices
Editorial
An integrated indicator of the condition of any community can be its species diversity, expressed by various indices [1, 2, 3]. In ichthyoplankton samples collected during complex expeditions at standard stations in the northern part of the Central-Eastern Atlantic (CEA), the number of species of eggs and larvae, as well as the number of each species, changed significantly from year to year. To assess the species diversity of ichthyoplankton, various indices of species diversity were used, widely used in ecological and faunal studies [1, 3, 4, 5, 6]. Changes in the indices of species diversity of ichthyoplankton in the northern part of the Central-Eastern Atlantic were analyzed using retrospective data and new materials.
- Formula
- Author
- N
- W ln
- 1
- −
- Margalef, et al. [11]
- N n
- N n i w i
- 2
- 1 log
- ⋅
- −∑
- = i
- 1
- )1
- (
- )1
- (
- 1
- Simpson, et al. [10] i i
- N
- N n n
- ∑
- −
- −
- ⋅
- −
- ⋅
- − w i log n
- -
- 1 i
- 2 i
- ∑
- =
- ⋅
- N n
- N w i
- Pielou, et al. [6]
- W log
- 2
- W – number of species; ni – number of the i-th species; N – total number of individuals.
Table 1: Indices of species diversity.
21 was used to collect materials. Stepwise oblique fishing was carried out at the horizons of 100, 50, 35, 25, 10 and 0 m for 1.5-3.0 minutes. on each horizon at the speed of the vessel 2.0-3.0 knots [7]. Plankton stations in the water areas of the survey were located above depths from 20 to 1000 m. Further processing of the materials was carried out under laboratory conditions under MBS-10 binocular microscopes (magnification 8 x 2, 8 x 4). During the cameral processing, the qualitative and quantitative compositions of fish eggs and larvae were determined. The species diversity of ichthyoplankton was estimated using the Shannon information and statistical imperative index, which is the most common in hydrobiology [8]. Although, as a measure of heterogeneity, the Shannon index takes into account the equalization of species abundance, very often, for a visual assessment of the degree of “polydominance”, the equalization index (imperative index) proposed by Pielou E, et al. [9] is used separately and represents the ratio of the observed diversity (Shannon index) to the maximum. In addition, the imperative Simpson index [10] and the parametric Margalef index Margalef, et al. [11] were calculated. Imperative indexes are mainly borrowed from information theory and have the advantage over parametric ones that they are applicable even when the form of distribution of species diversity by stations and/or by years varies significantly. Parametric indices show the presence of a larger or smaller number of species in the compared biocenoses, regardless of the number of each species (Table 1).
To assess the contribution of each species to the composition of ichthyoplankton complexes, the Paliy- Kovnatsky dominance index was used [12, 13].
i i i m n D 100 M N = ´ ´
Di – the dominance index for the i-th specie (%), mi - the number of samples in which the specie i was found, M - total number of samples, ni - the number of individuals of the i-th species, N - total number of individuals.
A total of 25 ichthyoplankton surveys were performed and analyzed during the period under review.
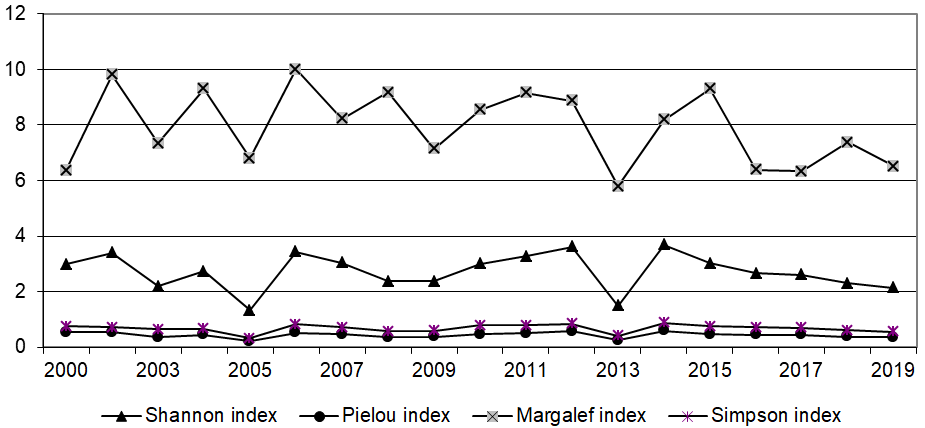
We will consider the dynamics of fluctuations in the indices of species diversity of ichthyoplankton in the northern part of the Central-Eastern Atlantic. Changes in the indices occurred synchronously, and the greatest fluctuations were observed in the parametric Margalef index (5.79-10.00) (Figure 1), analyzing which we can note the general trend of increasing species diversity from the northern part of Morocco to the region of Mauritania. Fluctuations in imperative indices were more smoothed out. The Simpson index varied in the range from 0.32 to 0.87 (Figure 1), i.e. according to the Odum classification [14], the ichthyoplankton complex in the northern part of the CEA had medium (index higher than 0.2) and high (index higher than 0.7) biotic diversity. The equalization index Pielu varied in the range from 0.22 to 0.59 (Figure 1). Basically, a situation of a certain monodominance was observed (the equalization index was ≤ 0.5 in 13 out of 19 years of observations), in the remaining years there was a relatively equal abundance of all species in a relatively complex ichthyoplankton complex.
According to the assessment of the degree of dominance according to the Palia-Kovnatsky index, the dominant species in the ichthyoplankton in the considered areas were the european sardine (Sardina pilchardus) and round sardinella (Sardinella aurita), the subdominants were the european anchovy (Engraulis encrasicolus), spar (family Sparidae) and rhombus (family Bothidae) (Figure 2).
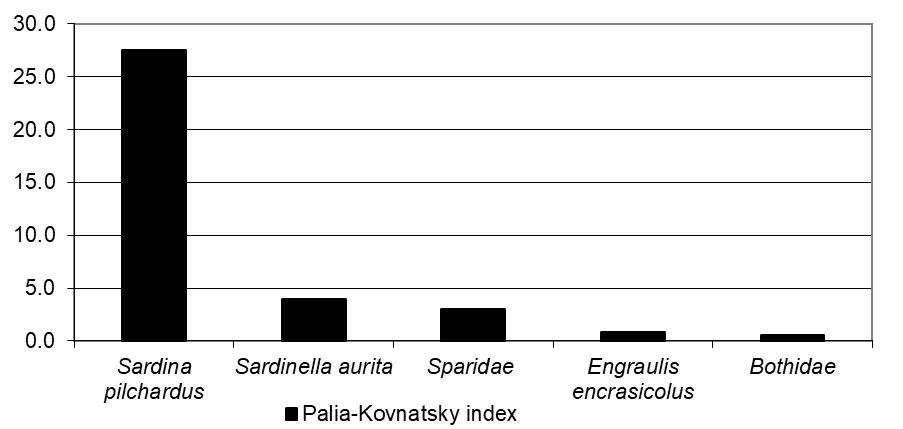
Moreover, the percentage of the contribution to the ichthyoplankton of the european sardine (subtropical species) decreased, and the percentage of the contribution of the round sardinella (tropical species) increased from Morocco to Mauritania.
References
-
Pesenko (1982) Principles and methods of quantitative analysis in faunistic research. Moscow Nauka, pp: 288.
-
Serebryakov (1984) Reproduction and early ontogeny of commercial fish in the North Atlantic. VNIRO, pp: 48.
-
Jacobs I (1975) Diversity stability and maturity in ecosystems influenced by human activities. Springer pp: 187-207.
-
Odum (1986) Ecology. pp: 376.
-
Burkovsky (2006) Marine biogeocenology. Organizing communities and ecosystems. pp: 285.
-
Arkhipov (2006) Dynamics of the abundance and peculiarities of the distribution of ichthyoplankton communities in the northern part of the central-eastern Atlantic and the seas of the Mediterranean basin. Kaliningrad: Publishing house AtlantNIRO pp: 232.
-
Methodological guidelines (1983) Methodical instructions for the collection of samples of zoo- and ichthyoplankton by the plank-collecting device «Bongo» and their processing. pp: 36.
-
Shannon CE (1948) A mathematical theory of communication. Bell System Tech JV 27(3): 623-656.
-
Pielou EC (1966) Species-diversity and pattern-diversity in the study of ecological succession. J Theor Biol 10(2): 370-383.
-
Simpson EH (1949) Measurement of diversity. Nature 163: 688.
-
Margalef R (1951) Species diversity in natural communities. Pablnes Inst Biol Apl Barcelona 9(5): 5-28.
-
Paliy (1961) Quantitative indicators in the processing of faunistic materials. Zoological journal 40(1): 3-6.
-
Kownacki A (1971) Taxocens of Chironomidae in streams of the Polish Hight Tatra Mts. Acta Hydrobiol 13(2): 439-463.
-
Odum EP (1975) Diversity as a function of energy flow. Ibid pp: 11-14.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework