Revisiting Isoglaridacris hexacotyle (Linton, 1897) Mackiewicz, 1968 (Cestoda: Caryophyllidae) from Catostomid Fishes in Rivers of Arizona
Isoglaridacris hexacotyle (Linton, 1897) Mackiewicz, 1968 was found in Catostomus insignis Baird & Girard and Catostomus clarkii Baird & Girard from 3 streams in Arizona. The morphology of I. hexacotyle is revisited using light microscopy revealing new features. Emphasis is placed on the morphological variability of its scolex. The re-examination of 100 more specimens from the Gila River provided an opportunity to revise the taxonomy of the species and correct and update some old statements and concepts by other observers.
Introduction
Mackiewicz JS [1] reviewed the morphology, anatomy, development, zoogeography, and systematics of the Caryophyllidea van Beneden (Cestoidea) and indicated the existence of 89 species and 37 genera in these monozoic cestodes. Ten years later, Mackiewicz JS [2] provided perspectives on the evolution, biology and zoogeography of 128 species in 45 genera of caryophyllids from the Nearctic, Neotropical, Palearctic, Ethiopian, Oriental, and Australian regions. Twelve years later, Mackiewicz JS [3] recognized about 150 species in 41 genera worldwide (except for the Neotropical Region). Of these, 14 genera and 90 species belonging to three families have been described from the Indo-Malayan region from catfishes (Siluriformes: Bagridae, Clariidae, Heteropneustidae, Schilbeidae and Siluridae), cyprinid, and cobitid fishes. A large number of inadequately described species from clariid and hetero-pneustid catfishes in India and neighboring countries have been invalidated by Ash, et al. [4, 5]. Most recently, however, Scholz, et al. [6] and Scholz, et al. [7] recognized only 117 species and 46 genera in order Caryophyllidea in Carus, 1863 parasitizing teleost fish (Cypriniformes, Siluriformes, and some catfish). Chubb JC [8] reviewed the seasonal ecology of the Caryophyllidea in the various climate zone of the world.
The order consists of four families, namely the Balanotaeniidae, Capingentidae, Caryophyllaeidae and Lytocestidae. Classification at the family level is based on the position of the inner longitudinal musculature in relation to the testes and vitelline follicles; “The caryophyllideans are exclusively intestinal parasites of freshwater teleosts with 85 species reported from cypriniform and 29 from siluriform fishes. Only few species of the Lytocestus with 2 species and monotypic Lytocestoides are known from mormyrids (Osteoglossiformes), characiform and cichlid (Perciforms) fishes, respectively.” Within families, 40 % of caryophyllideans parasitize North American suckers (Catostomidae), 26 % cyprinids, 10 % clariid catfishes and just a few caryophyllideans have been reported from 12 additional fish families” [7]. Freshwater annelids of the family Naididae, especially species previously placed in the Tubificidae (Tubifex tubifex, Limnodrilus spp.), serve as the only intermediate hosts [9].
Amin, et al. [10, 11, 12] recognized 14 species in 8 families of caryophyllid cestodes mostly from suckers (Catostomidae) in lake and river systems in Arizona and Wisconsin. These included Isoglaridacris hexacotyle (Linton, 1897) Mackiewicz, 1968 from the Sonora sucker, Catostomus insignis Baird & Girard and from the desert sucker Catostomus clarkii Baird & Girard in the Salt River, Verdi River, and Gila River systems, Arizona. See collections, following, for details. The following is a list of the 14 species of cestodes collected by Amin and organized by host species from Arizona (A) and Wisconsin (W).
- Carpoides Cyprinus (LeSueur); quillback Rowardleus pennensis Mackiewicz and Deutsch, 1976 (W)
- Catostomus clarkii Baird and Girard; desert sucker Isoglaridacris hexacotyle (Linton, 1897) Mackiewicz, 1968 (= Monobothrium hexacotyle (Linton, 1897) Nybelin, 1922, Moghe, 1925; Caryophyllaeus hexacotyle (Linton, 1897) Woodland, 1923, 1926; Glaridacris hexacotyle (Linton, 1897) Hunter, 1927) (A)
- Catostomus commersoni (Lacépède); white sucker Biacetabulum biloculoides Mackiewicz and McCrae, 1965 (= Biacetabulum biloculoides McCrae, 1961 [nomen nudum]; Glaridacris catostomi Van Cleave and Mueller, 1934; Hugghins, 1958; F. Meyer, 1958) (W) Biacetabulum macrocephalum MacCrae, 1962 (W) Biacetabulum sp. Hunter, 1927 (W) Glaridacris catostomi cooper, 1920 (= Caryophyllaeus catostomi Woodland, 1923; Moghe, 1925; Woodland, 1926) (W) Glaridacris laruei (Lamont, 1921) Hunter, 1927 (Glaridacris intermedius Lyster, 1940 fide Mackiewicz, 1965) (W) Hunterrella nodulosa Mackiewicz and McCrae, 1962 (W) Isoglaridacris folius Fredrickson and Ulmer, 1967 (W) Monobothium hunteri Mackiewicz, 1963 (W)
- Catostomus insignis Baird and Girard; Sonora sucker Isoglaridacris hexacotyle (Linton, 1897) Mackiewicz, 1968 (A)
- Cyprinus carpio Linn.; common carp Atractolytocestus huronensis Anthony, 1958 (W) Khawia iowensis Calentine and Ulmer, 1961 (W)
- Erimyzon sucetta (Lacépède); lake chubsucker Glaridacris laruei (Lamont, 1921) Hunter, 1927 (W) Isoglaridacris agminis Williams and Rogers, 1972 (W) Isoglaridacris multivitellaria Amin, 1986 (W) In Arizona, Amin OM, et al. [10] gave a detailed description of I. hexacotyle from C. insignis and C. clarkii which was originally described by Linton E, et al. [13] as Monobothrium hexacotyle from 8 specimens from the same hosts (Catostomus sp.) and localities in the Salt, Verdi, and Gila rivers. Linton’s E [13] description and figures were documented with adequate figures but Hunter’s GW, et al. [14] more detailed description of the same species as Glaridacris hexacotyle was based on Linton’s 8 specimens. “Catostomus sp.” was the type host that we later defined as C. insignis and C. clarkii. Mackiewicz JS [15] redescribed it as Isoglaridacris Mackiewicz, 1965 because it has 1 common gonopore as opposed to 2 gonopores in Glaridacris Cooper AR [16]. While C. clarkii is not the major host, it was apparently the host species instrumental in the transmission of I. hexacotyle outside of Arizona into Utah and Colorado. Catostomus clarkii was found in Nevada, Arizona, Utah, New Mexico, and the lower Colorado River basin particularly in the Gila River, Virgin River basin, White River basin, among other associated streams [17]. In Utah, Brienholt, et al. [18] reported I. hexacotyle from 2 omnivorous suckers: the bluehead or chiselmouth sucker Catostomus discobolis Cope and the flannelmouth sucker Catostomus latipinnis Baird & Girard in La Verkin Creek in southern Utah and the Fremont River near Hanksville. Catostomus latipinnis was also found to harbor I. hexacotyle in Wyoming streams including the Green River [19]. Both the Fremont River in Utah and the Green River in Utah and Wyoming are tributaries of the Colorado River. The identity of C. dicobolis in Utah appears to be the same as that of C. clarkii (= Pantosteus clarkii Baird and Girard) Sigler WF, et al. [20, 21] suggesting that the distribution of I. hexacotyle into Utah may be an extension of its native range in Arizona via C. clarkii. The chiselmouth sucker (also named bluehead suckers in Utah) were correctly identified by Sigler, et al. [20] but were used as common names for Pantosteus delphinus (Cope) which is in error. The same fish, C. clarkii whose native range extends into the White River basin, may also account of the finding of I. hexacotyle from an unnamed host in the “White River, 8 km W. Meeker, Rio Blanco County, Colorado [15]. On the other hand, the non-overlapping endemic range of the Sonora sucker, C. insignis, the other major host of I. hexacotyle, includes northern Sonora, Mexico, the San Francisco drainage in New Mexico, and the Gila and Salt River basins in Arizona [22]. No specimens of I. hexacotyle were reported from C. insignis in Mexico or New Mexico thus restricting the distribution of I. hexacotyle from C. insignis to Arizona locations only. In his description of I. hexacotyle, Amin, et al. [10] also demonstrated the differential growth patterns in various body regions as did Fischthal JH, et al. [23] in Glaridacris laruei from the C. commersoni in New York.
“Molecular phylogenetic studies confirm monophyly of caryophyllideans, but also reveal the non-monophyly of three of the four families (the family Balanotaeniidae is mono-generic)” [8, 24, 25]. In contrast, most genera seem to be monophyletic, with a few exceptions such as Glaridacris Cooper [7, 8].
Collections
A total of 2152 specimens of I. hexacotyle were collected from the carnivorous C. insignis (2127 specimens) and from the herbivorous C. clarkii (only 25 specimens) in the Lower Salt River at Tempe (33°24’53.17” N -111°54’33.52” W) between July, 1966 and January, 1967. Of these, 1031 worms (964 mature, 67 immature) were whole-mounted and measured [10]. Many additional specimens were collected from the Upper Verdi River at Perkinsville (34 54’ 06’’-112 11’ 26’’), the Lower Verdi River at Pinnacle Peak (32°21’7.21” N -112°52’57.56” W), and the Lower San Pedro River of the Gila River system at Aravaipa Creek (32°53’49.24”N,110°29›15.34»W), Arizona mostly from C. insignis in 1966-1967 [9, 10]. The total number of C. insignis and C. clarkii examined in all locations was 375 and 200, respectively. Mackiewicz JS [15] reported it from an additional location in Arizona in the Gila River, Tonto National Forest, Maricopa County as well as from Colorado (White River, Rio Blanco County; collected by G. Schmidt) and Utah from C. clarkii (Birch Creek tributary of the Virgin River, Zion National Park, Washington County). Brienholt JC, et al. [18] reported I. hexacotyle from other species of suckers in different locations in Utah that were discussed in the Introduction (above). A selection of 18 specimens from 288 newly examined specimens from the Gila River was used for microscopy (Figures 1-18).
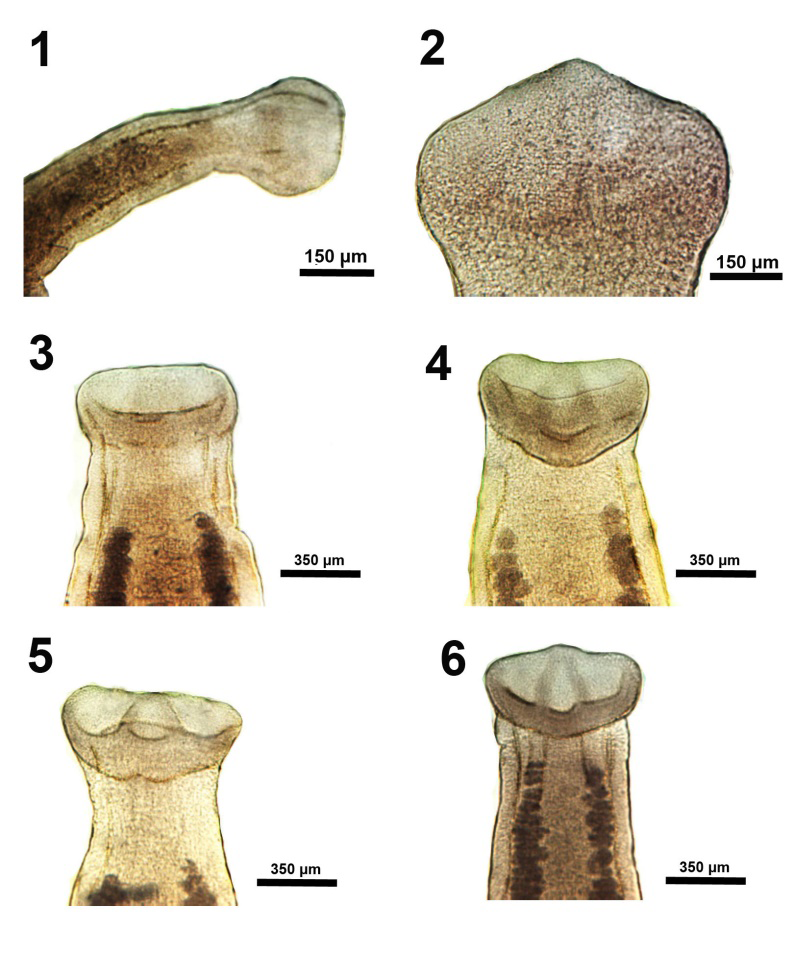
Figures 1-6: Ventral views of variations in the morphology of the scolex in specimens of Isoglaridacris hexacotyle collected from Catostomus insignis in the Gila River, Arizona in 1966-1967. 1. Anterior end of a young specimen with underdeveloped scolex having flat apex and no loculi but with early development of young testes and 2 column of vitellaria. 2-6. Progressive development of the septa between the loculi that are not fully developed.
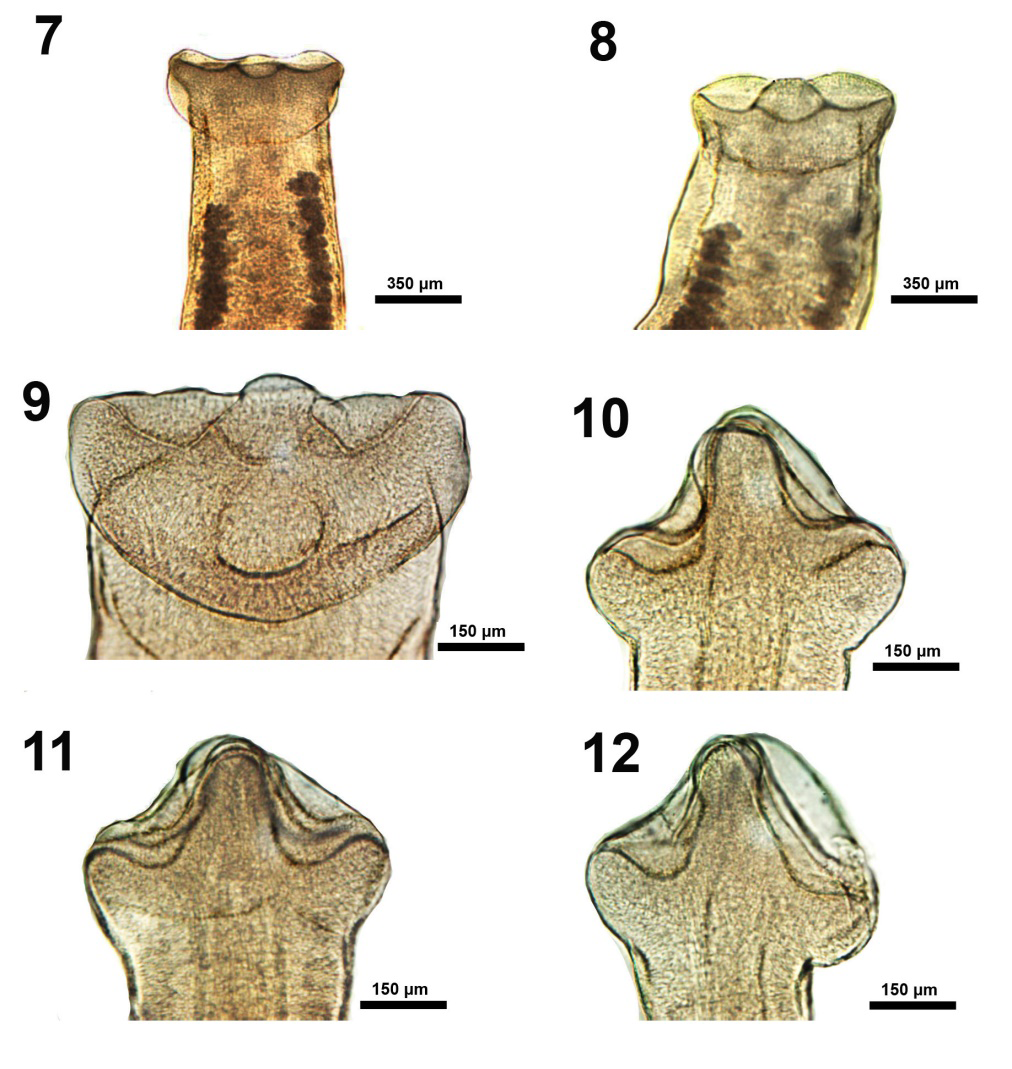
Figures 7-12: Ventral and lateral views of variations in the morphology of the scolex in specimens of Isoglaridacris hexacotyle collected from Catostomus insignis in the Gila River, Arizona in 1966-1967. 7-9. Further development in more mature cestodes showing different ventral perspectives of the developing loculi and the complete formation of the apical papilla. 10-12. Lateral perspectives of the scolex in 3 individuals showing the 2 dorsal and ventral lobes and the dividing ribs (septa) meeting apically to form the retractable conical papilla. The indentations marking the depth of the loculi are clearly visible.
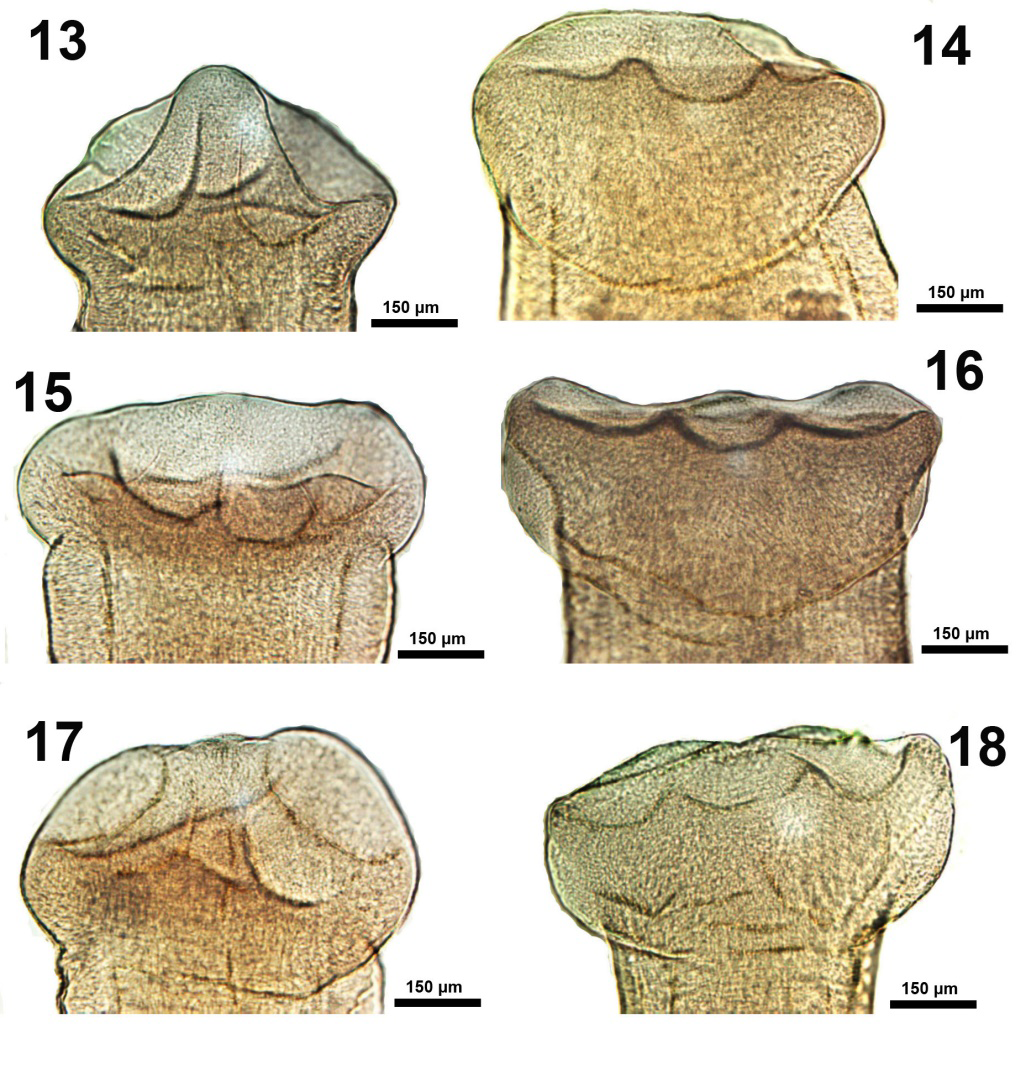
Figures 13-18: Ventral and lateral views of variations in the morphology of the scolex in specimens of Isoglaridacris hexacotyle collected from Catostomus insignis in the Gila River, Arizona in 1966-1967. An additional perspective of a scolex with elevated conical papilla (Figure 13) and others with retracted papillae (Figures 14-18) showing the scolex anatomy when the papillae are retracted with variable expressions related to functionality associated with attachment of worms at the time of capture. The margins and depths of loculi are variable in each worm.
Methods
Worms were initially stored in cold water then fixed in cold 70% ethanol. They were subsequently punctured with a fine needle then stained in Semichon’s carmine, destained in 4% hydrochloric acid in 70% ethanol, dehydrated in ascending concentrations of ethanol and cleared in 100% xylene. Whole worms were mounted in Canada balsam. Measurements are in micrometers or mm as noted in Table 1; the range is followed by the mean values between parentheses. Width measurements represent maximum width. Microscope images were created using 10X and 40X objective lenses of a BH2 light Olympus microscope (Olympus Optical Co., Osachi-shibamiya, Okaya, Nagano, Japan) attached to an AmScope 1000 video camera (United Scope LLC, dba AmScope, Irvine, California), linked to an ASUS lab top equipped with HDMI high definition multimedia interface system (Taiwan-USA, Fremont, California). Images from the microscope are transferred from the lab top to a USB and stored for subsequent processing on a computer.
| Character | Linton (1897) | Hunter (1930) | Mackiewicz (1969) | Amin (1968, 1969, this paper) |
|---|---|---|---|---|
| N | 8 | 8 | 30 | 694 + 100 |
| Total length (mm) | 8.0-14.5 | 8.8-18.0 | 7.0-15.5 (10.4) | 1.53-12.04 (5.75) |
| Post-vitelline: pre-vitelline region | --- | --- | --- | 7.53 : 1.00 |
| Scolex L (mm) | -- X 0.7 | 0.51-0.61 | --- | 0.16-0.73 (0.44) |
| Scolex W (mm) | -- X 0.72 | 0.51-0.61 | --- | 0.24-1.31 (0.70) |
| Neck W (mm) | -- X 0.68 | 0.45-0.0.76 | --- | 0.18-1.26 (0.53) |
| Scolex W: neck W | --- | --- | --- | 1.32 : 1.00 |
| W at gonopore (mm) | -- X 1.0 | 1.03-1.23 | 0.50-1.01 | 0.14-1.10 (0.56) |
| Cirrus sac W (mm) | -- X 0.27 | 0.17-0.23 | --- | 0.06-0.31 (0.18) |
| No. of testes | --- | 175-200 | 132-205 (162) | 172-220 (194) |
| Testis L (mm) | --- | 0.144-0.264 | --- | 0.125-0.277 (0.186) |
| Testis W (mm) | --- | 0.072-0.199 | --- | 0.094-0.135 (0.114) |
| Egg L X W (μm) | 38-40 X 20 | 37-41 X 23-30 | 32-40 X 28-30 | 27-38 (33) X 20-25 X (24) |
Table 1: Comparative measurements of mature Isoglaridacris hexacotyle from Catostomus insignis collected from the Lower Salt Rive
Results
Amin OM, et al. [9] initially described I. hexacotyle from C. insignis and C. ckarkii in the Gila, Salt, and Verdi Rivers as Glaridacris hexacotyle guided by Linton E [13] and Hunter GW [14] who studied the same 8 specimens collected from Catostomus sp. in the same localities. Linton’s material were deposited in Vial No. 4793 of the U.S. National Museum, Washington, D.C. and the label read “From sucker (Catostomus sp.) inhabiting the Gila River and the Salt River, Arizona; E. Palmer.” The above mentioned “Catostomus sp.” must have been C. insignis, the normal host of this cestode especially that C. clarkii which commonly inhabits the same waters as C. insignis, belonged in genus Pantosteus Cope then. The locality where E. Palmer made his collection must have been at the old junction of the Gila and Salt rivers that have been dry since, with the closest locality being the Lower Salt River irrigation canal at Tempe where Amin [9] made his collections and near where Mackiewicz made his 1964 collections [15]. Mackiewicz JS and Amin OM [15] agreed that I initially publish this cestode in the Genus Glaridacris as the change in nomenclature to Isoglaridacris that Mackiewicz JS [15] proposed has not been published yet [9]. The scolex in both genera are similar but some important generic differences exist. For instance, there are 2 gonopores in Glaridacris but 1 in Isglaridacris and the ovarian lobes are H-shaped in Glaridacris but have inverted A- shape in Isoglaridacris. Hunter GW [14] created a much more elaborate description than the modest account of Linton’s E [13] but he also created some confusion. For instance, he described the ovarian arms of his Glaridacris hexacotyle as “lobate, H-shaped, possessing no unusual characteristics” [14]. Linton E [13] did not describe the ovary except to mention that it is” with lateral folds extending nearly to cirrus.” In “the inverted A-shaped ovary often observed in the 2,000 plus specimens examined, the ovarian material in the apex of the A is frequently obscured by the post-ovarian vitellaria” thus becoming confused with the H-shaped ovary of Hunter [14].
We have re-examined 100 additional specimens of I. hexacotyle collected from C. insignis in the Gila River, other than the 1031 specimens (964 mature and 67 immature) described and measured by Amin OM [10]. Our new observations 55 years after the latest treatment of that species were generally in agreement with our descriptions and comparable to those of Linton [13], Hunter [14] and Mackiewicz [15] with some variations. We provide below a brief revised account of our qualitative morphological findings and a comparative morphometrical data (Table 1) high-lightening variations from the other accounts of Linton, Hunter, et al. [13, 14, 15].
Morphological Description of Adults
Body of medium length, dorso-ventrally flattened, covered with very thin cuticle enclosing a layer of cortical parenchyma surrounding internal medullary parenchyma containing glandular-like cells, reproductive, muscular, and excretory systems. Anterior scolex conical, rounded posteriorly, divided in 2 dorsal and ventral lobes each with 3 loculi divided by 2 ribs meeting at apex to form a retractable conical papilla displaying a wide range of diversity (Figures 1-18). Neck, narrowest part of body, not especially differentiated. Vitelline glands in 2 regions; the first beginning just posterior to neck anteriorly and extending in 2 lateral bands reaching cirrus and the second in a separate cluster posterior to ovary (post ovarian vitellaria). Over 200 oblong testes in medullary region occupy central space shorter than that of vitellaria anteriorly and posteriorly. Coils of the vas deference at mid-body dilate posteriorly into a muscular ejaculatory duct enlarging into the seminal vesicle which narrows ventrally into cirrus. Cirrus joins utero-vaginal canal which opens ventrally in a single gonopore. Lobate ovary shaped like inverted letter-A often appearing H-shaped when posterior apex is obscured by post-ovarian vitellaria and beginning just posterior to level of cirrus. Convoluted uterus between arms of ovary empties into female cloaca before forming vagina and subsequently joining the oviduct. Posterior end bluntly rounded where excretory pore drains. Operculate eggs with rough surface.
Discussion
Linton E [10] did not justify the inclusion of I. hexacotyle in the genus Monobothrium Diesing, 1863 except to mention that “In specimens made transparent by oil of cloves, the anatomy of the reproductive organs was seen to agree closely with that made out for Monobothrium terebrans Linton, found in a sucker (Catostomus adrens) from Heart Lake, Yellow Stone National Park, Wyoming.” Hunter GW [14] relegated this cestode to the genus Glaridacris Cooper [16] “which is characterized by the possession of three pairs of loculi or bothria. The type of genital atrium, ovary, position of uterine coils, presence of post-ovarian vitellaria, and external seminal vesicle clearly mark it as belonging to this genus.” Mackiewicz JS [15] placed it in his new genus Isoglaridacris Mackiewicz JS [15] “based on having a single gonopore; Glaridacris has two gonopores and therefore, cannot receive this species. All of the other characters, such as vitellaria in lateral rows, cuneiform scolex and type of ovary, are clearly those of Isoglaridacris.” Amin OM [9] temporarily placed it in Glaridacris waiting for Mackiewicz’s JS [15] redescription under Isoglaridacris before Amin OM [10] described his specimens as I. hexacotyle with mutual agreement with Mackiewicz.
Morphometrically, the size and width of our specimens, collected from C. insignis in the Lower Salt River at Tempe, and of corresponding structures such as testes, cirrus sac and eggs were smaller than those reported by other observers (Table 1) who collected their specimens from the same locality and the same host species, C. insignis, but not the same individuals, earlier. Amin OM [10] documented heavier and more frequent infections in the largest of 261 hosts classified in 5 size classes ranging between 100 and 350 mm long with the largest gravid cestodes (15 mm long) found in the largest hosts (300-350 mm long). Most immature cestodes (85% of 2127 worms collected) were recovered in the smallest 2 host size classes (100-150 & 150-200 mm long). Our smaller cestodes (Table 1) were younger and more slender worms that were mostly collected from smaller hosts. “The fact that more immature cestodes were recovered from smaller fish hosts suggest that there is a correlation between host size and parasite size …. and that the larger cestodes reported by Linton E [13] and Hunter GW [14] were similarly obtained from large hosts” (Amin OM [9], p.80-81, Table 14, Fig. 5). Neither Linton E [13] nor Hunter GH [14] indicated the size of their hosts but Mackiewicz JS [15] did. Most (160) of the 177 specimens collected in December, 1964 in the Salt River at Tempe from 225-494 mm long (mean 359 mm) Sonora suckers by Mackiewicz JS [15] were stained and measured of which 30 gravid adults were reported. Hosts examined by Mackiewicz JS [15] were considerably larger than ours which accounts for the larger sizes of his material (Table 1). Younger worms in smaller fish clearly attain further development and larger size as fish become older and larger in size.
The scolex is of particular interest. Linton E [13] concluded his description of it by stating that “The six loculi meet at the apex of the head and form a central papilla which may project forward as a sharp conical elevation or be contracted to a low eminence ; doubtless other and diverse shapes may be assumed by this versatile organ.” Hunter GW [14] had similar, but less colorful, remarks of the scolex which he concluded by stating “it should be noted that there is probably more variation in living material for all scolices were fairly well contracted.” Mackiewicz JS [15] showed line drawings of only 2 scolex variations and none was portrayed by other observers. We have provided the first microscopic images of all possible scolex variation encountered in our extensive collection of non-contracted specimens (Figures 1-18). Descriptions are provided in the legends to figures.
In his description of I. hexacotyle, Amin OM [10] also studied its growth patterns and demonstrated the acceleration of growth of the post -vitelline region, the region of reproductive activity, compared to the pre-vitelline region, as well as the growth in the scolex, cirrus sac, and width at the common gonopore region. Fischthal, et al. [23] showed comparable growth of the post-vitelline region of Glaridacris laruei from the C. commersoni in New York [23].
While Linton’s E [13] description was sufficiently adequate to provide clear diagnostic recognition of I. hexacotyle, it was incomplete and lacking much detail on the extent of anterior and posterior distribution of vitellaria and testes, shape of ovary and commissure, position of post-ovarian vitellaria, and measurements of various structures such as the scolex and testes. In his detailed revised description of the same material, Hunter GW [14], included comprehensive treatments of the reproductive, muscular and excretory systems largely left out by Linton’s E [13] description. Both Linton E [13] and Hunter GW [14] did not recognize the extent of the anterior distribution of vitellaria and testes, even though Hunter GW [14] clearly showed the anterior end of a specimen with lateral vitellaria more distinctly anteriorly positioned than the anterior-most testes which is characteristic of this species as acknowledged by Mackiewicz [15]. Hunter’s GW [14] detailed revision was additionally marred by a few inaccuracies and errors: 1. Describing the cestode as having “monotonous appearance” then as “one of the most unique which have come under my observations” (p.61,62). 2. Has “practically no neck” then “neck is very short and shows little differentiation” (p.61). 3. Ovary “H-shaped.” (p.66). It is actually inverted A-shaped with occasional obscuring of the posterior apex of the A with post-ovarian vitellaria. 4. “The ovarian commissure is V-shaped” (p.66). It is actually usually straight, just a little wider at interface with ovarian lobes; see Fig. 1c in Amin OM [10] and Figures 4&5 in Mackiewicz JS[15]. In a sample of 68 of the 100 newly examined specimens, 64% of specimens had straight commissures and 36% had commissures that were mostly straight posteriorly but arched anteriorly like a crescent, but never V-shaped. 5. Egg shell “non-operculate” (p.67). Eggs are actually operculate and include a few vitelline cells each; see also Mackiewicz [15]. 6. “common genital atrium is lacking or inconspicuous” (p.68); it is actually always present but sometimes unremarkable. In his specific diagnosis, Hunter GW [14] himself implied that there is one gonopore as the term “common genital atrium” was used.
Acknowledgments
This project was supported by an Institutional Grant from the Parasitology Center, Inc. (PCI), Scottsdale, Arizona.
Declarations
- Compliance with Ethical Standards
- Conflict of interest: The authors declare no conflicts of interest or competing interests.
- Ethical approval: The authors declare that they have observed all applicable ethical standards.
- Availability of data: All presented and related data are available by contacting the senior author.
References
-
Mackiewicz JS (1972) Caryophyllidea (Cestoidea): a review. Exper Parasitol 31(3): 417-512.
-
Mackiewicz JS (1982) Caryophyllidea (Cestoidea): Perspect Parasitol 84(2): 397-417.
-
Mackiewicz JS (1994) Order Caryophyllidea van Beneden in Carus, 1863. In: Khalil LF, Jones A, Bray RA, et al. (Eds.), Keys to the cestode parasites of vertebrates. CAB Intern Wallingford UK, pp: 21-43.
-
Ash A, Scholz T, Oros M, Kar PK (2011a) Tapeworms (Cestoda: Caryophyllidea), parasites of _Clarias batrachus_ (Pisces: Siluriformes) in the Indomalayan region. J Parasitol 97(3): 435-459.
-
Ash A, Scholz T, Oros M, Levron C, Kar PK (2011b) Cestodes (Caryophyllidea) of the stinging catfish _Heteropneustes fossilis_ (Siluriformes: Heteropneustidae) from Asia. J Parasitol 97(5): 899-907.
-
Scholz T, Kuchta R, Oros M (2021) Tapeworms as pathogens of fish: A review. J Fish Dis 44(12): 1-18.
-
Scholz T, Kuchta R (2017) A digest of fish tapeworms. Vie et milieu-Life and environment 67(2): 43-58.
-
Chubb JC (1982) Seasonal occurrence of helminths in freshwater fishes Part IV. Adult Cestoda, Nematoda and Acanthocephala. Adv parasitol 20: 1-292.
-
Amin OM (1968) Helminth fauna of Suckers (Catostomidae) of the Gila River System, Arizona. PhD Thes Ariz St Univ (No 68-1644) Univ Microfilms. Ann Arbor, Michigan; Dissert Abstr, USA. 28: 3521.
-
Amin OM (1969) Helminth fauna of suckers (Catostomidae) of the Gila River System, Arizona. II. Five parasites from _Catostomus_ spp. Am Midland Nat 82: 429- 443.
-
Amin OM (1974) Intestinal helminthes of the white sucker, _Catostomus commersoni_ (Lacépède), in SE Wisconsin. Proc Helminthol Soc Wash 41: 81-88.
-
Amin OM (1986) Caryophyllaiedae (Cestoda) from lake fishes in Wisconsin with a description of _Isoglaridacris multivitellaria_ sp. n. from _Erimyzon sucetta_ (Catostomidae). Proc Helminthol Soc Wash 53: 48-58.
-
Linton E (1897) Notes on cestode parasites of fishes. Proc U S Nat Museum 20: 423-456.
-
Hunter III GW (1930) Studies on the Caryophyllaeidae of North America. Illinois Biological Monographs 11 (1927). pp: 186.
-
Mackiewicz JS (1968) _Isoglaridacris hexacotyle_ comb. N. (Cestoidea: Caryophyllidea) from catostomid fishes in southwestern North America. Proc Helminthol Soc Wash 35: 193-196.
-
Cooper AR (1920) _Glaridacris catostomi_ n. g., n. sp., a cestodarian parasite. Transact Amer Microscop Soc 39: 5-24.
-
Arizona Game & Fish (2007) Desert Sucker.
-
Briendolt JC, Heckmann RA (1980) Parasites from two species of suckers (Catostomidae) from southern Utah. G Bas Natur 40(2): 149-156.
-
Mitchum DL (1995) Parasites of fishes in Wyoming. 1995. Wyoming Game and Fish Dept Cheyenne. pp: 304.
-
Sigler WF, Miller RR (1963) Fishes of Utah. Utah State Department of fish and game. Salt Lake City. pp: 203.
-
Bailey RM, Fitch JE, Herald ES, Lachner EA, Lindsey CC, et al. (1970) A list of common and scientific names of fishes from the United States and Canada. Amer Fisher Soc Spec Publ No 6 Washington DC , USA, pp: 174.
-
Minckley WL (1973) Fishes of Arizona. Ariz Game and Fish Depart Phoenix. pp: 160-162.
-
Fischthal JH, Carson DO, Vaught RS (1982) Size allometry of the caryophyllidean cestode _Glaridacris laruei_ from the white sucker. J Parasitol 68: 1175-1177.
-
Olson PD, Scholz T, Poddubnaya LG, Littlewood DTJ (2008) On the derived position of _Archigetes_ and the early evolution of the tapeworms (Platyhelminthes: Cestoda). J Parasitol 94: 898-904.
-
Brabec J, Scholz T, Králová-Hromadová I, Baszalovicsová E, Olson PD (2012) Substitution saturation and nuclear paralogs of commonly employed phylogenetic markers in the Caryophyllidea, an unusual group of non-segmented tapeworms (Platyhelminthes). Int J Parasitol 42(3): 259- 267.
-
Scholz T, Oros M (2017) Caryophyllidea. In: Caira JN, Jensen K, et al. (Eds.), Planetary Biodiversity Inventory (2008-2017): Tapeworms from Vertebrate Bowels of the Earth. Univ Kansas, Nat Hist Mus, Spec Publ No 25 Lawrence Kansas, USA, pp: 47-64.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework