Serum Phenoloxidase Activity in the Hemolymph of Giant Freshwater Prawn Macrobrachium rosenbergii (De Man, 1879)
Melanization is one of the major immune responses in arthropods, particularly crustaceans. The prophenoloxidase (proPO) catalyzes the oxidation of mono or di or polyphenols, a reaction that is the key initial step of melanin formation. We attempted to study the immune response in M. rosenbergii by melanization reaction as produced by serum phenoloxidase (PO) activity. The substrate affinity of the PO enzyme was determined using different phenolic substrates and it was found that the mono, di and poly phenols were oxidized hence the enzyme was characterized as catecholoxidase and l-3,4-dihydroxyphenylalanine (L–DOPA) showed the highest substrate affinity to the enzyme. The biochemical parameters that determined optimum enzyme activity were found such as 1 mM L–DOPA concentration showing at the absorbance of 470 nm, 10 mM Tris–HCl buffer pH 7.5 (brown colour formation), temperature 40 °C and 10 min incubation. Kinetic characteristics of serum from the prawn were also determined. Determination of optimal conditions of PO activity in the serum has also been attempted. We also identified the serum having a potent PO activity was inhibited by phenylthiourea (PTU) and activation of such as trypsin, sodium dodecyl sulphate (SDS) and laminarin in the freshwater prawn M. rosenbergii.
Introduction
The Macrobrachium rosenbergii commonly known as giant freshwater prawn is one of the most commercially important species of the palaemonidae family. The popularity of increased economic value is accountable to its large size, water quality tolerance, handling stress tolerance, adaptability to the feed and importantly to its increased immune response leading to high disease resistance such as against the white spot syndrome virus [1]. The invertebrate immune response mechanism is very simple and can be considered a precursor to vertebrate immunity. Vertebrate immunity is more complex in understanding due to the participation of cellular components and humoral factors which results in memory and antigen–specificity [2, 3, 4, 5, 6], which is absent in the case of invertebrate immunity. However, from recent studies, it has been identified that some cellular and cell-free hemolymph factors show specificity for non–self or damaged cells in invertebrates [7]. Henceforth, among the two types of immunity, innate or natural immunity and acquired immunity, invertebrates exhibit innate immunity which is further classified into humoral and cellular immunity.
Crustaceans possess an open circulatory system, where nutrients, oxygen, hormones and cells are present in the hemolymph. The circulatory cells which are the hemocytes function analogous to vertebrate leukocytes since they play a role in non–self recognition and elimination [8, 9] and coagulation [10, 11]. During a pathogen attack, the physical barriers such as the exoskeleton detain the pathogenic microbial entry into the tissues [12]. When the physical barrier fails in its function microbial invasion into tissues occurs. Thus, the defense mechanism gets activated based on the characteristics of the pathogen recognized. After the recognition, different effector mechanisms such as prophenoloxidase (_pro_PO) system, phagocytosis, and encapsulation occurs through proteolytic pathways resulting in the dismissal of invasive microbes out of the system [13].
The proPO system in crustaceans exhibits similarities mainly in function with the complement system where the chain of enzymatic reactions takes place [8]. The enzymatic system can be activated by lipopolysaccharides (LPSs) or peptidoglycans and β–glucans which are specific molecules in the cell walls of bacteria, and fungi respectively. These mechanisms are stimulated by the pattern recognition receptors (PRR) which are germ line encoded. They recognize these microbial surface determinants and activate distinct signaling cascades which regulate specific immune related proteins [14]. In the case of peptidoglycan and, β–glucans, the peptidoglycan recognition or binding proteins (PGBPs) and β–glucan recognition or binding proteins (β–GBPs) binds to form a complex which induces activation of proPO cascade [15]. At times, the β–GBPs recognized structure could be a laminarin [16]. In some crustaceans such as the horseshoe crab, these receptors or binding sites do not play a major role in the proPO activation system. This system activation leads to hemocyte degranulation [17] since the proPO defensive mechanism involves cellular populations such as semi- granulocytes and hemocytes with big refractile granules. The proPO gets activated after its release to the haemocoel in the presence of calcium ions [18].
The mechanism process is the proPO in its active state is cleaved by proteolysis to its active form PO. However, PO is copper dependent, catalyses the synthesis of o–phenols (catechols) from mono–phenols, and oxidation of o-_diphenols to _o–quinones [19]. After which the non–enzymatic polymerization of o-_phenols leads to the production of melanin which response to the entry of pathogen i.e., PO catalyses the reaction where tyrosine is converted into DOPA and DOPA to DOPA-quinone, the precursor of melanin [20]. Melanin is a brownish black pigment that has biological properties such as the inhibition of fungal and bacterial enzyme activity [21]. This induces amplification of the proPO system through cell degranulation where some regulatory proteins are present such as the α–macroglobulin. These adhesion proteins result in the stimulation of multicellular response as encapsulation, and phagocytosis which is evident from _Carcinus maenas hyaline cells thus destroying the invasive foreign microbes [22, 23].
It has also been evident that change in absorbance maxima, substrate concentration, ionic strength, pH, temperature and time interval [24, 25, 26], dissolved oxygen [27] causes fluctuation in immune responses, especially in _pro_PO activation. Therefore, the study attempted to understand and unveil the difference in serum PO activity based on the biochemical, physiological and enzymatic condition and thus to determine its activity.
Furthermore, to understand the substrate affinity of the serum PO enzyme activity the kinetics of the enzymes rate of reaction was determined in the Lineweaver-Burk plot. It is evident that the kinetics of the crustaceans phenoloxidases vary among the different components of the hemolymph as well as species [25, 26]. Moreover, to our knowledge, there is no information on the optimized conditions of the serum phenoloxidase enzyme activity. Hence, an attempt has been made to optimize the conditions for determining the PO activity of serum including the Km and Vmax value of M. rosenbergii. Hence, an attempt has been made to optimize conditions for determining serum PO activity and its functional activity and susceptibility to an inhibitor and various activators in freshwater prawn M. rosenbergii.
Materials and Methods
Source of Experimental Animals
Adult intermoult specimens of the giant freshwater prawn, M. rosenbergii weighing around 25–30 ± 1.53 g were collected from private hatcheries in and around Chennai, India. In the laboratory, the prawns were maintained in plastic troughs containing continuously aerated freshwater. The animals were fed with egg white ad libitum and were acclimatized to the laboratory conditions for at least 4 to 5 days prior to use. Fifty percent of water was renewed daily to remove the unfed and fecal materials. The uninjured, intermoult animals were used throughout this study.
Hemolymph Collection and Preparation of Serum
The hemolymph from fifty adult prawns per tank was pooled as a replicate to measure sample each animal 500 µl without an anticoagulant solution was allowed to clot at –20 ˚C for 6 h. The coagulated sample was disturbed with a glass rod and centrifuged at 12, 000 rpm for 20 min, 4 ˚C. The supernatant (= serum) was collected and used for protein estimation and phenoloxidase activity to immediately freeze at –80˚C until assayed.
Determination of Protein Concentration
The protein content in the serum was determined according to Bradford [28] using bovine serum albumin as the standard. All chemicals used in the study were purchased from Sigma–Aldrich, St. Louis, MO, USA.
Oxidation of Phenolic Substrates by Serum
The oxidative activity of serum was tested by incubating 100 µl serum with 1.9 ml of different phenolic substrate solutions (5 mM; tyramine, L–tyrosine, L–DOPA, DL– DOPA, dopamine, catechol, hydroquinone and pyrogallol) for 20 min at 25°C. The color developed was measured spectrophotometrically at 300 - 600 nm against a reagent blank in which suitable substrates were substituted for serum.
Effect of Different Concentrations of L–DOPA by Serum
To serum (100 µl) was mixed with 1.9 ml of L–DOPA at different concentrations (1.0 – 10.0 mM) and incubated for 20 min at 25°C. The color developed was measured spectrophotometrically at 470 nm against a reagent blank of L–DOPA.
Effect of Different Time Interval of L–DOPA by Serum
To serum (100 µl) was mixed with 1.9 ml of 1 mM L–DOPA and incubated for different time intervals for 1 – 45 min at 25°C. The color developed was measured spectrophotometrically at 470 nm against a reagent blank of L–DOPA.
Effect of Ionic Strength on Oxidation of L-DOPA by Serum
The effect of buffer ionic strength on oxidation of L-DOPA (1 mM) by serum was assessed by incubating 100 µl serum with 1.9 ml of L–DOPA prepared in Tris-HCl buffer (pH 7.5) with different ionic strength (5 – 100 mM) at 25°C. After 10 min the optical density of each of these reaction mixtures was determined spectrophotometrically at 470 nm against a reagent blank of L–DOPA.
Effect of pH on Oxidation of L-DOPA by Serum
The ability of serum to oxidize L–DOPA (1 mM) at different pH was tested by incubating 100 µl of serum with 1.9 ml of substrate solutions prepared in 10 mM Tris–HCl buffer at different pH (5.0 – 9.0) for 10 min at 25°C. The color developed was measured spectrophotometrically at 470 nm against a reagent blank of L–DOPA.
Oxidation of L–DOPA by Serum Exposed to Different Temperature
Serum samples (each 100 µl) were held at temperatures ranging from 4°C to 80°C for 10 min. The color developed was measured spectrophotometrically at 470 nm against a reagent blank of L–DOPA.
Kinetic Parameters, Km and Vmax of Serum Phenoloxidase Enzyme
To measure the kinetic parameters of the serum PO enzyme, different concentrations of L-DOPA (1 - 10 mM) were mixed with 100 µl of serum incubated for 10 min and absorbency read at 470 nm. The Michaelis-Menten constant was estimated by plotting substrate concentrations [S] and the rate of PO activity [V]. Lineweaver-Burk plot was plotted as reciprocal of substrate concentration [1/S] and rate of PO activity [1/V]. The resultant plot gave a line that intercepted X-axis to give the -1/Km value and intercepted Y axis to give the 1/Vmax. The slope Km/Vmax was determined and the resultant plot was rechecked from equation Y= mx + c.
Phenoloxidase Activity
The phenoloxidase activity of serum with the phenolic substrate was tested by incubating 100 µl of serum made up to 2 ml of 1 mM L–DOPA (10 mM Tris–HCl, pH 7.5) for 10 min at 40°C. The colour developed was measured spectrophotometrically at 470 nm against a blank reagent of L–DOPA.
Inhibition of Serum PO Activity
In this experiment, 100 μl of serum was separately mixed with 100 μl of 3 mM PTU. In control, PTU was substituted with Tris–HCl buffer (10 mM; pH 7.5) and incubated for 10 min at 40°C. An aliquot of 200 μl of the reaction mixture from the control or experimental was incubated with made up to 2 ml of 1 mM L–DOPA for 10 min at 40°C. The optical density of both the control and experiments was measured spectrophotometrically at 470 nm.
Activation of Serum PO activity
In this experiment, 100 μl of serum was separately mixed with 100 μl of trypsin (2.1 mg.ml–1), SDS (6 mg.ml–1) and laminarin (3 mg.ml–1). In control, these chemicals were substituted with Tris–HCl buffer (10 mM; pH 7.5), and pre- incubated for 10 min at 40°C. An aliquot of 200 μl of the reaction mixture from control or experiments was incubated with made up to 2 ml of 1 mM L–DOPA for 10 min at 40°C. The optical density of both the control and experiments was measured spectrophotometrically at 470 nm.
Statistical Analysis
The serum PO activity data were expressed as mean ± SD of triplicate experimental trials from five determinations. The variation between experimental trials and control were evaluated by SPSS/18 software (version 20; SPSS, New York) by one-way analysis of variance (ANOVA) at 0.05, 0.01 probabilities (*P < 0.05, **P < 0.01).
Results
Serum PO Activity with Various Substrates
The serum separated from the hemolymph of the freshwater prawn M. rosenbergii, was tested for PO activity with various phenolic substrates. Among the diphenolic substrate, the serum showed the highest activity with L– DOPA (470 nm) when compared to DL–DOPA (490 nm), catechol (450 nm) and dopamine (460 nm) as shown in Figure 1. However, the monophenols including tyramine and L–tyrosine, or polyphenols such as pyrogallol and hydroquinone activated to show any oxidation by serum. In all the above experiments, the results were expressed in absorbance at 300 - 600 nm. Since the highest oxidative activity was obtained with L–DOPA, this substrate was used to detect PO activity in all subsequent experiments performed in this study.
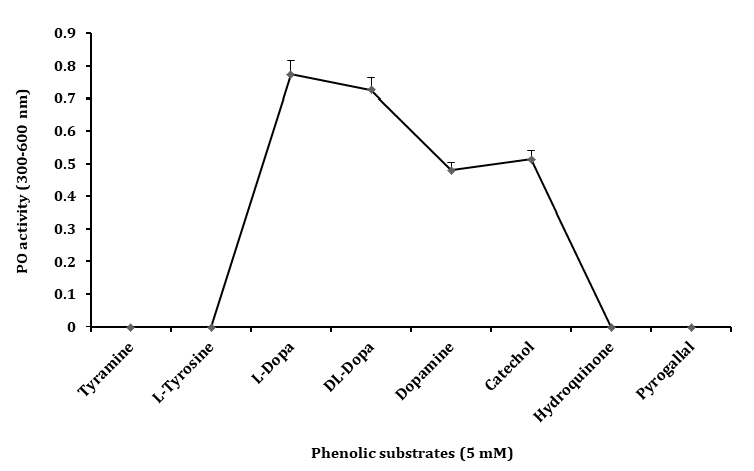
Figure 1: Phenoloxidase activity in the serum of Macrobrachium rosenbergii on different phenolic substrates (5 mM) in Tris- HCl buffer (Tris 10 mM, pH 7.5) incubated at 25°C for 20 min and absorbance at 300–600 nm. The PO activity in optical density obtained at absorbance maxima of respective substrates. Data represent mean of triplicate repeats of five determinations (mean ± SD) in the same way in all further experiments.
Effect of L–DOPA on PO Activity
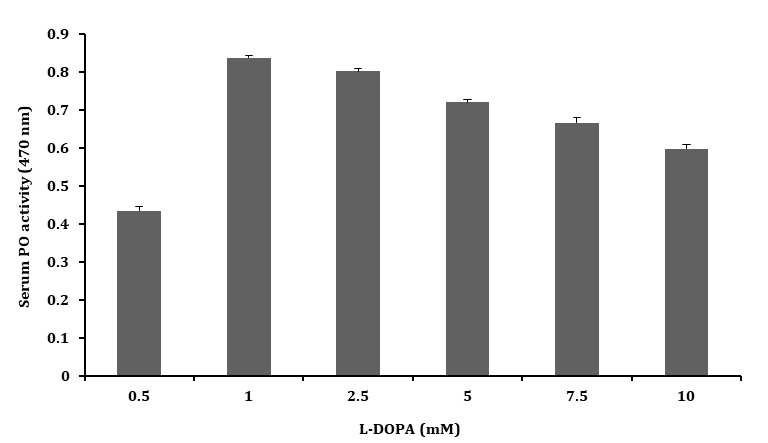
The serum PO activity was tested with different concentrations of L–DOPA (1.0 – 10.0 mM) and the PO activity was found to be higher with L–DOPA of 1 mM concentration than that of higher concentrations (5.0, 7.5 and 10.0 mM) as shown in the Figure 2. This result clearly suggested that the optimum concentration for testing PO activity in serum was 1 mM of L–DOPA.
Effect of Time Interval on PO Activity
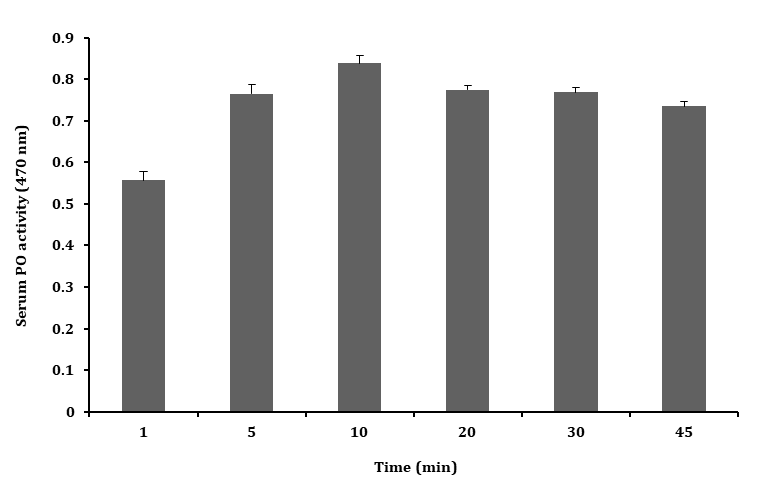
The PO activity of serum was tested oxidation of L–DOPA
at a different time interval (1 – 45 min) and the highest PO activity was found in 10 min when compared to other time intervals tested as shown in the Figure 3.
Effect of Ionic Strength on PO Activity
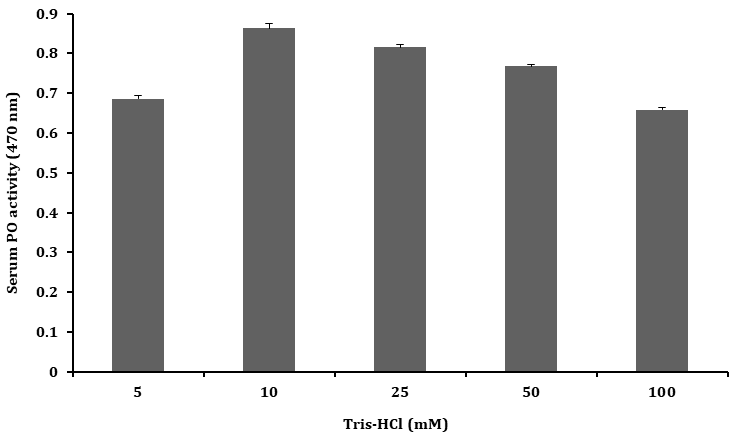
The PO activity of serum was tested with Tris–HCl buffer (pH 7.5) of different ionic strengths (5 – 100 mM) and highest PO activity was found with 10 mM Tris–HCl buffer when compared to other ionic strengths tested as shown Figure 4.
Effect of pH on PO Activity
The PO activity of serum was tested for oxidation of L–DOPA at pH 5.0 to 9.0, pH above 7.5 (brown colour formation), and the observed PO activity decreased as shown in Figure 5. The pH of 7.5 increased the PO activity taking as the optimum pH for the study of serum PO activity.
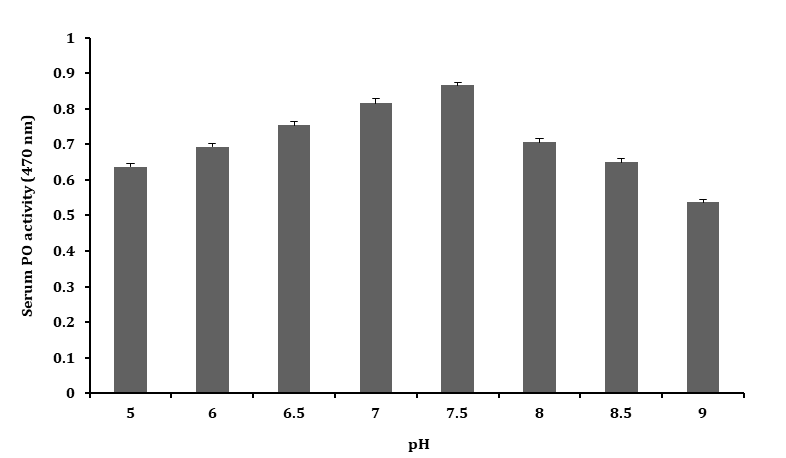
Effect of Temperature on PO Activity
The serum samples in buffer solution (10 mM Tris–HCl, pH 7.5) incubated for 10 min at temperatures ranging from
10 – 80 °C showed, stable and peak PO activity at 40 °C, which was taken as the optimum temperatures for PO activity. At temperature below or above 40 ˚C, a decline in PO activity was observed as shown in Figure 6. In summary, the optimal conditions for measuring serum PO activity on 1 mM L-DOPA were 10 mM Tris-HCl, pH 7.5 at 40 °C for 10 min at 470 nm.
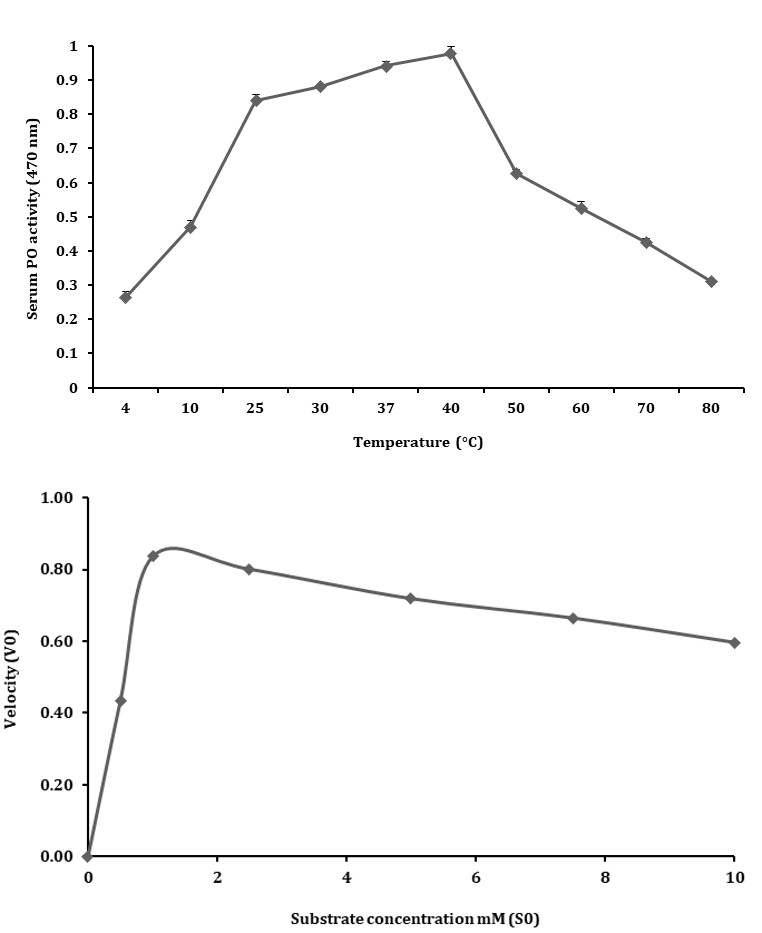
Kinetic Behavior
The kinetic characteristics of serum PO activity were determined from the rate of reactions as calculated from the oxidation of L-DOPA at different concentrations (1 – 10 mM) in 20 min. The Michaelis-Menten constant Km was calculated to be 0.5 and the maximum velocity (Vmax) was 0.014 as shown in Figure 7A. Application of Km and Vmax yielded Lineweaver- Burk plot with a line slope (3.7) which on extrapolation intercepted at -1.8 that was plotted as -1/Km and on Y-axis 1/ Vmax was derived at 2 on X-axis (Figure 7B).
Figure 7A: Kinetic properties of PO activity in serum of freshwater prawn M. rosenbergii at different substrate concentrations of L-DOPA as shown in Michaelis–Menten curve.
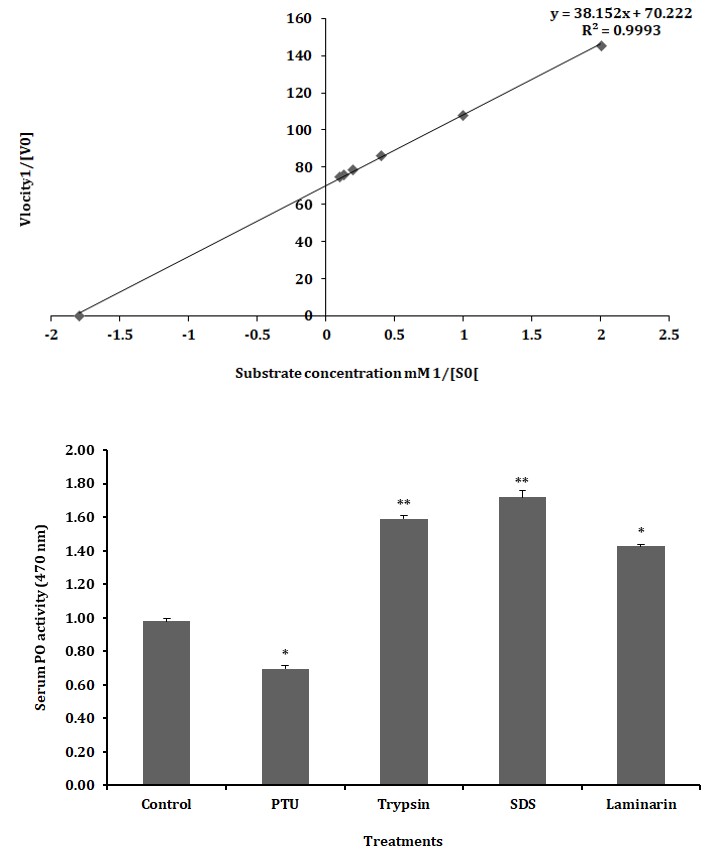
Figure 7B: The Km and Vmax values are calculated using Lineweaver Burk plot of PO activity in serum of freshwater prawn M. rosenbergii with L-DOPA as substrate.
Inhibition and Activation of Serum PO Activity
The treatment of serum with PTU (3 mM) decreased the oxidation of L–DOPA compared to the control and the reduction was found to be about 29% was statistically significant (p < 0.05; Figure 8). Similarly, the treatment of serum with trypsin, SDS and laminarin (fungal cell wall component) resulted in a significant (p < 0.05; p < 0.01) increase in PO activity of L–DOPA respectively, when compared to control (Figure 8).
Asterisk indicates significant variation determined from the value obtained for a treatment to untreated control by one way ANOVA at *P < 0.05, P < 0.01. Data represents mean of triplicate repeats of five determinations (mean ± SD). Figure 8:** Inhibition and activation of serum phenoloxidase activity in the freshwater prawn M. rosenbergii.
Discussion
The PO activity of serum in the freshwater prawn M. rosenbergii has been depicted in the present study. Phenoloxidase, the active form of prophenoloxidase is the key step in melanin formation and plays a major in immune responses in the hemolymph of arthropods [24, 25, 26, 29, 30, 31, 32]. Melanisation occurs during phenol oxidation which comprises two reactions in the melanin synthetic pathway which is the oxidation of tyrosine and DOPA to dopaquinone which produces brown pigment dopachrome through a non- enzymatic reaction.
In the present study, the phenoloxidase activity of serum from M. rosenbergii has been studied. Initially, to characterize the substrate affinity of serum phenoloxidase activity, mono-phenols such as tyramine and L–tyrosine, diphenols as the L–DOPA, DL–DOPA, dopamine and catechol and polyphenols as the pyrogallol and hydroquinone was used. The result produced in Figure 1 indicates that the serum has the highest affinity towards L–DOPA among the other monophenols, diphenols and polyphenols. It has been shown that DOPA works best to synthesize dopachrome by oxidation of tyrosinase to produce melanin [33]. In general, the reactions are catalyzed by the enzyme occurrence which forms an enzyme substrate complex in which non-covalent bonding occurs between the active site of the enzyme and the substrate [34]. Similar to the produced result the use of L–DOPA as substrate was also reported in marine crab Scylla tranquebarica [35], shrimp in Penaeus californiensis [36], freshwater prawn M. rosenbergii [25, 31] and many other crustaceans.
Every enzymatic reaction occurs in a particular biochemical condition. Therefore, biochemical studies were taken up to understand the optimum conditions for serum phenoloxidase activity. The reactions with L–DOPA at different concentrations (1 – 10 mM) as shown in Figure 2. The result obtained shows an increased value in the concentration of 1 mM compared to other concentrations. This result shows the level of substrate binding with respect to substrate concentration. Our results compared to other crustaceans species as in kuruma shrimp Penaeus japonicus [37] and portunid crab Charybdis japonica [38] the reaction occurred at 3.4 mM, 2.5 mM in brown shrimp Penaeus californiensis [39] and 1.5 mM in Chinese white shrimp Penaeus chinensis [40] which is nearly similar to M. rosenbergii.
The time interval required for the substrate to bind to the enzyme for its complete activation is also very important. Therefore, the highest serum activity from M. rosenbergii in accordance with time interval was observed as in Figure 3. It was found that in time intervals of 10 minutes the PO activity showed the highest value. It was also observed that the time interval required for PO activation sometimes varied among the species itself. In the case of C. japonica, the incubation time interval required was 40 minutes [38] while 5 minutes was recorded in Penaeus monodon [41] and Penaeus paulensis [42].
Enzyme activity or activation always gets enhanced at particular physiological and chemical conditions such as differences in pH, different buffer concentrations, temperature and so on. Therefore, an ideal ionic interaction was estimated in serum by using different ionic strengths of Tris–HCl buffer (5 – 100 mM) as shown in Figure 4 and serum PO activity was studied. The favorable ionic strength of Tris–HCl was obtained at 10 mM. The ionic strength of the buffer varies among different decapod crustaceans such as 50 mM Tris–HCl buffer concentration was ideal in Panulirus argus [43] while a similar accordance with M. rosenbergii was noted in spiny lobster Panulirus japonicus [33, 44] with the highest activity in 10 mM concentration. To follow this, the optimum pH was also studied and pH 7.5 was shown as the ideal pH among varying ranges of (5.0 – 9.0) as shown in Figure 5. We can also infer that the pH value is also organism specific because a similar Penaeus setiferus showed a pH of 7.5 while, a pH of 6.0 was also observed in Charybdis japonica phenoloxidase, while Botryllus schlosseri showed the range of pH 7.0 – 7.5 [45, 46].
Among various parameters, temperature plays a major physiological role in enhancing an enzymatic reaction because the intactness of the binding site can be altered with temperature variations. In the case of serum PO activity, various temperatures were set ranging from 10-80°C and as per Figure 6 and the favorable temperature analysed was 40 °C. A resemblance with value was also observed in crab C. japonica [38], spiny lobster Panulirus argus [42], and Japanese prawn Penaeus chinensis [37].
The enzyme kinetics of the serum PO activity was determined using the Michaelis-Menten curve by plotting various concentrations of L-DOPA (1-10 mM) and the rate of reaction was determined in 20 min (1/V). The initial rate of reaction increased up to a maximum reaction velocity after which it stabilized and then declined. The Km value determined for substrate enzyme affinity was 0.5 mM and this suggested a strong affinity between the enzyme and L-DOPA and the Vmax was calculated as 0.014. The Lineweaver-Burk plot showed a slope of 0.014 with a correlation coefficient of R2 = 0.999. This indicated that the enzyme had active sites to maintain a steady increase in the rate of reaction. The kinetic and biochemical characteristics of the serum PO activity demonstrate a distinct PO activity among the other crustaceans [25, 26, 38, 39].
The oxidation of L-DOPA, by serum of freshwater prawn M. rosenbergii showed that serum possesses PO activity. In this connection, it is worthwhile to note that many investigators have reported the presence of phenoloxidase activity in the hemocyte lysate supernatant (HLS) of M. rosenbergii [26, 27, 38, 41, 47, 48, 49, 50, 51, 52, 53, 54]. These investigators, apart from demonstrating PO activity in hemocytes or plasma, have also shown that this PO responds to several environmental factors (such as salinity, temperature, pH, hypoxia), physiological status such as moulting, optimum conditions of enzyme activity [25, 26, 31, 38] and bacterial infectivity [54]. Thus, the hemocytic or plasma phenoloxidase clearly appears to respond to environmental, physiological, enzyme optimum conditions and pathological conditions of the freshwater prawn M. rosenbergii. In addition, most of these workers have also demonstrated that exogenous protease such as trypsin, detergent like SDS or certain non-self molecules (laminarin) activate the proenzymic state prophenoloxidase, thereby implicating its potential role in non-self recognition. On the other hand, the present study, which showed the presence of PO activity in the serum of freshwater prawn M. rosenbergii inhibition of PTU clearly showed that the serum indeed possesses a significant amount of PO activity.
Although, there are reports on the presence of PO activity in the serum of crabs [55], crayfish [56] and anomuran crabs [24] the major hemolymph component of crustaceans that possess PO activity remains unclear. A few investigators claim that the hemocytes appear to be the major repertoire of phenoloxidase [57] which accounts for nearly 90 and 85% of the hemolymph PO activity in shrimp Penaeus paulensis and lobster Panulirus interruptus respectively [42, 58]. However, considering the presence of PO activity in the serum of PTU inhibited clearly indicates that the serum appears to be the hemolymph PO activity in the freshwater prawn M. rosenbergii.
A statistically significant increase in serum PO activity upon treatment with an exogenous protease such as trypsin a total of 38.43% increase with an anionic detergent SDS increase of 43.08%. However, indicate that serum possesses protease and anionic detergent activable proPO activity a common feature observed with hemocyte or plasma PO activity in M. rosenbergii [25, 26, 27, 31, 38, 41, 47, 52]. Thus, these results together with the observations of earlier workers clearly indicate that the freshwater prawn M. rosenbergii possesses PO activity in hemocytes and plasma. The results reveal that the pretreatment of serum resulted in a significant increase in PO activity in the presence of laminarin (a polymer of β-1,3 glucan derived from fungal cell wall materials). It may be noted that while working on crayfish, Astacus astacus reported that the serum PO activity activated to respond to laminarin [56]. These observations show that the serum PO activity found in crayfish A. astacus and M. rosenbergii appear to same in responding to the non-self molecules.
Conclusion
In the present study, we concluded that the above observation is the serum PO enzyme was studied under optimized conditions of catechol oxidase nature. However, the PO activity in serum is very distinct by showing the highest specific binding affinity to diphenol L–DOPA only and activation of trypsin, detergents like SDS and fungal cell wall component laminarin. These results clearly finding provides evidence that the serum of M. rosenbergii act as both cellular and humoral immune system enzymatically functions as phenoloxidase. Thus the serum of freshwater prawn M. rosenbergii possesses differential physiological or immunological functions, apart from their established respiratory function.
Conflict of Interest
The authors declare that they have no conflict of interest.
References
-
Hameed ASS, Charles MX, Anilkumar M (2000) Tolerance of _Macrobrachium rosenbergii_ to white spot syndrome virus. Aquaculture 183(3-4): 207-213.
-
Hoffmann JA, Kafatos FC, Janeway CA, Ezekowitz RAB (1999) Phylogenetic perspectives in innate immunity. Science 284(5418): 1313-1318.
-
Iwanaga S, Lee BL (2005) Recent advances in the innate immunity of invertebrate animals. J Biochem Mol Biol 38(2): 128-150.
-
Coates CJ, Paiva EMC (2020) Multifunctional roles of hemocyanins. Subcell Biochem 94: 233-250.
-
Huang Y, Ren Q (2020) Research progress in innate immunity of freshwater crustaceans. Dev Comp Immunol 104: 103569.
-
Cerenius L, Söderhäll K (2021) Immune properties of invertebrate phenoloxidases. Dev Comp Immunol 122: 104098.
-
Liu S, Zheng SC, Li YL, Li J, Liu HP (2020) Hemocyte mediated phagocytosis in Crustaceans. Front Immunol 11: 268.
-
Sritunyalucksana K, Söderhäll K (2000) The proPO and clotting system in crustaceans. Aquaculture 191(1-3): 53-69.
-
Morales RP, Alejo VM, Perera E (2019) The clotting system in decapod crustaceans: History, current knowledge and what we need to know beyond the models. Fish Shellfish Immunol 84: 204-212.
-
Cerenius L, Liang Z, Duvic B, Keyser P, Hellman U, et al. (1994) Structure and biological activity of a 1,3–β–D– glucan–binding protein in crustacean blood. J Biol Chem 269(47): 29462-29467.
-
Vázquez L, Maldonado G, Agundis C, Pèrez A, Cooper EL, et al. (1998) Participation of asialic acid specific serum lectin from freshwater prawn _Macrobrachium_ _rosenbergii_ hemocytes in the recognition of non-self cells. J Exp Zool 279(3): 265-272.
-
Söderhäll K (1982) Prophenoloxidase activating system and melanization a recognition system of arthropods? A review. Dev Comp Immunol 6(4): 601-611.
-
Ratcliffe NA, Rowley AF, Fitzgerald SW, Rhodes CP (1985) Invertebrate immunity: basic concepts and recent advances. Int Rev Cytol 97: 183-350.
-
Lee SY, Söderhäll K (2001) Characterization of a pattern recognition protein, a masquerade–like protein, in the freshwater crayfish _Pacifastacus leniusculus_. J Immunol 166(12): 7319-7326.
-
Koizumi N, Imamura M, Kadotani T, Yaoi K, Iwahana H, et al. (1999) The lipopolysaccharide–binding protein participating in hemocyte nodule formation in the silkworm _Bombyx mori_ is a novel member of the C– type lectin superfamily with two different tandem carbohydrate-recognition domains. FEBS Lett 443(2): 139-143.
-
Lee SY, Wang R, Söderhäll K (2000) A lipopolysaccharide and β–1,3–glucan binding protein from hemocytes of freshwater crayfish _Pacifastacus leniusculus_: purification, characterization, and cDNA cloning. J Biol Chem 275(2): 1337-1343.
-
Liu H, Jiravanichpaisal P, Cerenius L, Lee BL, Söderhäll I, et al. (2007) Phenoloxidase is an important component of the defense against _Aeromonas hydrophila_ infection in a crustacean _Pacifastacus leniusculus_. J Biol Chem 282(46): 33593-33598.
-
Adachi K, Hirata T, Nishioka T, Sakaguchi M (2003) Hemocyte components in crustaceans convert hemocyanin into a phenoloxidase-like enzyme**.** Comp Biochem Physiol B 134(1): 135-141.
-
Yamamoto K, Yakiyama M, Fujii H, Kusakabe T, Koga K, et al. (2000) Expression of prophenoloxidase mRNA during silkworm hemocyte development. Biosci Biotechnol Biochem 64(6): 1197-1202.
-
Hellio C, Nilles AB, Gagnaire B, Renault T, Guyon HT (2007) Demonstration of a true phenoloxidase activity and activation of a _pro_PO cascade in Pacific oyster, _Crassostrea gigas_ (Thunberg) _in vitro_. Fish shellfish Immunol 22(4): 433-440.
-
Smith VJ, Söderhäll K (1983) β–1,3 Glucan activation of crustacean hemocytes _in vitro_ and _in vivo_. Biol Bull 164(2): 299-314.
-
Aspán A, Söderhäll K (1991) Purification of prophenoloxidase from crayfish blood cells and its activation by an endogenous serine proteinase. Insect Biochem 21(4): 363-373.
-
Cerenius L, Söderhäll K (2004) The prophenoloxidase activating system in invertebrates. Immunol Rev 198: 116-126.
-
Shanthi S, Sivakumar MR, Rayvathy B (2021) Serum phenoloxidase activity in the hemolymph of the anomuran crab Albunea symmysta **(Linnaeus, 1758)** **(**Decapoda: Anomura: Albuneidae). J Crustac Biol 41(1): ruab003.
-
Sivakumar MR, Shanthi S (2022) Plasma and hemocyanin phenoloxidase derived from the hemolymph of the giant freshwater prawn _Macrobrachium rosenbergii_ (De Man, 1879). Crustacea-Nano, Micro and Macro Studies.
-
Sivakumar MR, Abisha Adline F, Shanthi R (2022) Hemocytes lysate supernatant derived phenoloxidase activity in the hemolymph of giant freshwater prawn _Macrobrachium rosenbergii_ (De Man, 1879). Int J Zoo Animal Biol 5(5): 000403.
-
Cheng W, Liu CH, Hsu JP, Chen JC (2002) Effect of hypoxia on the immune response of giant freshwater prawn _Macrobrachium rosenbergii_ and its susceptibility to pathogen Enterococcus. Fish Shellfish Immunol 13(5): 351-365.
-
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principles of protein-dye binding. Anal Biochem 72: 248-254.
-
Ashida M, Brey PT (1997) Recent advances in research on the insect prophenoloxidase cascade. In: Brey PT, Hultmark D (Eds.), Molecular Mechanism and Immune Response in Insects, Chapman & Hall; London, UK, pp: 133-172.
-
Söderhäll K, Cerenius L, Johansson MW (1996) The prophenoloxidase activating system in invertebrates. New Directions in Invertebrate Immunology. In: Söderhäll K, Iwanaga S, et al. (Eds.), (Fair Heaven, NJ: SOS Publications) pp: 335-374.
-
Sivakumar MR, Denis M, Shanthi S, Arumugam M (2017) Phenoloxidase activity in humoral plasma, hemocyanin and hemocyanin separated proteins of the giant freshwater prawn _Macrobrachium rosenbergii_. Int J Biol Macromol 102: 977-985.
-
Sivakumar MR, Denis M, Shanthi S, Arumugam M (2020) Agglutination of plasma, hemocyanin, and separated hemocyanin from the hemolymph of the freshwater prawn _Macrobrachium rosenbergii_ (De Man, 1879) (Decapoda: Caridea: Palaemonidae). J Crustac Biol 40(3): 309-315.
-
Adachi K, Hirata T, Nagai K, Sakaguchi M (2001) Hemocyanin a most likely inducer of black spots in Kuruma prawn (_Penaeus japonicus_) during storage. J Food Sci 66(8): 1130-1136.
-
Dominguez R, Souchon H, Spinelli S, Dauter Z, Wilson KS, et al. (1995) A common protein fold and similar active site in two distinct families of β-glycanases_._ Nat Struct Biol 2(7): 569-576.
-
Sabu AS, Jithendran KP, Gopal C (2016) Properties of phenoloxidase from haemocytes of _Scylla tranquebarica_. J Appl Biol Biotechnol 4(1): 47-52.
-
López JH, Galván TG, Albores FV (1996) Activation of the prophenoloxidase system of the brown shrimp (_Penaeus_ _californiensis_ Holmes). Comp Biochem Physiol C 113(1): 61-66.
-
Zhao J, Qi XY, You YM, Wang JX, Zhou PG (1997) Study on some characteristics of phenoloxidase from Japanese prawn, _Penaeus japonicus_. J Shanghai Fisheries University 6: 157-165.
-
Liu G, Yang L, Fan T, Cong R, Tang Z, et al. (2006) Purification and characterization of phenoloxidase from crab _Charybdis japonica_. Fish Shellfish Immunol 20(1): 47-57.
-
Galván TG, López JH, Albores FV (1999) Prophenoloxidase from brown shrimp (_Penaeus californiensis_) hemocytes. Comp Biochem Physiol B 122(1): 77-82.
-
Fan TJ, Wang XF (2002) Purification and partial biochemical characterization of phenoloxidase from _Penaeus chinensis_. Acta Biochim Biophys Sin 34(5): 589- 594.
-
Sung H, Chang H, Her C, Chang J, Song Y (1998) Phenoloxidase Activity of Hemocytes Derived from _Penaeus monodon_ and _Macrobrachium rosenbergii._ J Invertebr Pathol 71(1): 26-33.
-
Perazzolo LM, Barracco MA (1997) The prophenoloxidase activating system of the shrimp _Penaeus paulensis_ and associated factors. Dev Comp Immunol 21(5): 385-395.
-
Morales RP, Alejo VM, Perera E, Ruiz ZP, Jiménez EA (2007) Phenoloxidase activity in the hemolymph of the spiny lobster _Panulirus argus_. Fish Shellfish Immunol 23(6): 1187-1195.
-
Adachi K, Hirata T, Nagai K, Fujisawa S, Sakaguchi M (1999) Purification and characterization of prophenoloxidase from kuruma prawn _Penaeus japonicus_. Fish Sci 65(6): 919-925.
-
Simpson BK, Marshall MR, Otwell WS (1987) Phenol oxidase from shrimp (_Penaeus setiferus_): purification and some properties. J Agric Food Chem 35(6): 918-921.
-
Frizzo A, Guidolin L, Ballarin L, Sabbadin A (1999) Purification and partial characterisation of phenoloxidase from the colonial ascidian _Botryllus schlosseri_. Mar Biol 135: 483-488.
-
Cheng W, Chen JC (2000) Effects of pH, temperature and salinity on immune parameters of the freshwater prawn _Macrobrachium rosenbergii_. Fish Shellfish Immunol 10(4): 387-391.
-
Cheng W, Wang CH (2001) The susceptibility of the giant freshwater prawn _Macrobrachium rosenbergii_ to _Lactococcus garvieae_ and its resistance under copper sulfate stress. Dis Aquat Organ 47(2): 137-144.
-
Cheng W, Chen JC (2002a) Effects of environmental factors on the immune responses of freshwater prawn _Macrobrachium rosenbergii_ and other decapod crustaceans. J Fish Soc Taiwan 29(1): 1-19.
-
Cheng W, Chen JC (2002b) The virulence of _Enterococcus_ to freshwater prawn _Macrobrachium rosenbergii_ and its immune resistance under ammonia stress. Fish Shellfish Immunol 12(2): 97-109.
-
Cheng W, Wang CH, Chen JC (2003) Effect of benzalkonium chloride stress on immune resistance and susceptibility to _Lactococcus garvieae_ in the giant freshwater prawn _Macrobrachium rosenbergii_. Dis Aquat Organ 53(3): 223- 229.
-
Li JT, Lee PP, Chen OC, Cheng W, Kuo CM (2005) Dopamine depresses the immune ability and increases susceptibility to _Lactococcus garvieae_ in the freshwater giant prawn, _Macrobrachium rosenbergii_. Fish Shellfish Immunol 19(3): 269-280.
-
Chang CC, Lee PP, Hsu JP, Yeha SP, Cheng W (2006) Survival, and biochemical, physiological, and histopathological responses of the giant freshwater prawn, _Macrobrachium rosenbergii_, to short-term trichlorfon exposure. Aquaculture 253(1-4): 653-666.
-
Sahoo PK, Pillai BR, Mohanty J, Kumari J, Mohanty S, et al. (2007) _In vivo_ humoral and cellular reactions, and fate of injected bacteria _Aeromonas hydrophila_ in freshwater prawn _Macrobrachium rosenbergii_. Fish Shellfish Immunol 23(2): 327-340.
-
Wang LG, Li EC, Qin JG, Du ZY, Yu N, et al. (2015) Effect of oxidized fish oil and α-tocopherol on growth, antioxidation status, serum immune enzyme activity and resistance to _Aeromonas hydrophila_ challenge of Chinese mitten crab _Eriocheir sinensis_**.** Aquac Nutr 21(4): 414-
-
Söderhäll K, Unestam T (1979) Activation of serum prophenoloxidase in arthropod immunity. The specificity of cell wall glucan activation and activation by purified fungal glycoproteins of crayfish phenoloxidase. Can J Microbiol 25(3): 406-414.
-
Söderhäll K, Häll L, Unestam T, Nyhlén L (1979) Attachment of phenoloxidase to fungal cell walls in arthropod immunity. J Invertebr Pathol 34(3): 285-294.
-
López JH, Galván TG, Jiméunez SG, Clark GP, Albores FV (2003) In the spiny lobster (_Panulirus interruptus_) the prophenoloxidase is located in plasma not in haemocytes. Fish Shellfish Immunol 14(2): 105-114.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework