Nosocomial Infection in Veterinary Medicine: A Rare Event or Neglected by Veterinarians?
In humans, nosocomial infections (HI), also known as healthcare-associated infections (HAIs), are those acquired by patient during hospital stay. They are unrelated to the reason for which the patient was hospitalized, and may manifest, mostly due to bacteria, during or after hospitalization. The environment has a strong connection with cases of nosocomial infection in human medicine, however, in veterinary medicine, there is still not much information regarding this connection, since there is no effective data on these infections. The main multidrug-resistant microorganisms involved in nosocomial infections in humans are: Methicillin-resistant Staphylococcus (MRS), Vancomycin-resistant Enterococcus (VRE), Gram-negative bacteria (Enterobacteriaceae family, Pseudomonas aeruginosa and Acinetobacter baumanii) that produce extended-spectrum betalactamase (ESBL) and those producing carbapenemases. As previously mentioned, bacteria are the main microorganisms involved in cases of nosocomial infections, which may come from the patient himself (endogenous contamination) after an immunological decline or come from the environment or cross-contamination (other animals or via health professionals). Generally, the bacteria involved in nosocomial infections are multi-resistant to antimicrobials, especially those most used in hospitals, which makes treatment and patient improvement even more difficult. Despite research related to antimicrobial resistance, the reality of the clinical routine of animals does not seem to be in line with the world reality. There are few studies in veterinary medicine regarding nosocomial infections (some extremely punctual), In the other hand, these infections are already a reality in veterinary hospitals, mainly of small animals, and, unfortunately, they are most of the time neglected due to the difficulty of their traceability or even by the lack of knowledge.
Introduction
Nosocomial Infection
In humans, nosocomial infections (HI), also known as healthcare-associated infections (HAIs), are those acquired by patient during hospital stay. They are unrelated to the reason for which the patient was hospitalized, and may manifest, mostly due to bacteria, during or after hospitalization [1, 2].
In veterinary medicine, there are still few studies related to nosocomial infections [3]. However, some important studies have contributed to assist this subject, such as the study by Ruple-Czerniak, et al. [4] which evaluated five school veterinary hospitals. Of the 1535 dogs and 416 cats treated in 12 weeks, 28.3% (16.3% dogs and 12% cats) had at least one case of nosocomial infection. Benedict KM, et al. [5] using a questionnaire carried out in 38 veterinary hospitals (European and North American), showed that 82% (31) reported the occurrence of at least one outbreak of nosocomial infection in the last five years preceding the interview.
In human hospitals, it is estimated that tens of thousands of people die each year of nosocomial infections [6]. The annual cost related to these infections ranges from 28 to 45 billion dollars, not including indirect costs such as community care, lost productivity and lost wages [7]. In veterinary medicine, treatment costs, indemnities due to owner dissatisfaction, loss of business and outbreaks of nosocomial infections are increasingly drawing the attention of veterinarians around the world [8, 9].
Actions like the conscious use of antimicrobials, choosing the correct route of administration and monitoring the resistance profile through the antibiogram technique, must be taken to combat multidrug-resistant microorganisms and avoid the impact on public health [10].
The main factors that contribute to nosocomial infections, both in human medicine and in veterinary medicine are: the increased intensive care, a longer hospitalization, the surgical procedures, the use of intravascular devices and probes, the use of antimicrobials and the use of immunosuppressive drugs [8, 11, 12]. However, an aggravating factor in veterinary medicine that compromise the paciente hygiene is the act of licking wounds and the low concern with the control of nosocomial infections [3].
The pathogens responsible for nosocomial infections in animals could come from other patients, veterinarians, employees and/or the hospital environment [2]. These pathogens have a great zoonotic potencial, which points to the need for control strategies [13].
The environment has a strong connection with cases of nosocomial infection in human medicine, however, in veterinary medicine, there is still not much information regarding this connection, since there is no effective data on these infections [9]. In Brazil, Sfaciotte, et al. [14] isolated methicillin-resistant Staphylococcus (MRS), vancomycin- resistant Enterococcus (VRE), Gram-negative extended- spectrum beta-lactamase (ESBL) and carbapenemase- producing bacteria in a school veterinary hospital. Despite the authors not having made the connection with cases of nosocomial infections, this result could represents a risk factor for hospitalized animals and people circulating in that environment.
Some critical contamination points in a hospital are: doorknobs, light switches, computers, cell phones, cage doors, stethoscopes, thermometers, gags and, mainly, the hands of doctors and employees, which, when hygiene and disinfection fails, increase the chances of hospital infections [8, 14, 15, 16].
As previously mentioned, bacteria are the main microorganisms involved in cases of nosocomial infections, which may come from the patient himself (endogenous contamination) after an immunological decline, or come from the environment or cross-contamination (other animals or via health professionals). Generally, the bacteria involved in nosocomial infections are multi-resistant to antimicrobials, especially those most used in hospitals, which makes treatment and patient improvement even more difficult [3].
As there is not much knowledge about the main microorganisms that cause nosocomial infections in animals, the great concern is with those that affect humans, since they have a great zoonotic potential. Another major concern is that they are commonly found in healthy animals/humans (Staphylococcus, Enterococcus and Enterobacteriaceae) as well as in the environment (Pseudomonas aeruginosa and Acinetobacter baumanii) [17].
The main multidrug-resistant microorganisms involved in nosocomial infections in humans are: Methicillin-resistant Staphylococcus (MRS), Vancomycin-resistant Enterococcus (VRE), Gram-negative bacteria (Enterobacteriaceae family, Pseudomonas aeruginosa and Acinetobacter baumanii) that produce extended-spectrum beta-lactamase (ESBL) and those producing carbapenemases [18].
Vancomycin Resistant Enterococcus (VRE)
The microorganisms belonging to the genus Enterococcus are Gram positive cocci, catalase negative, facultative anaerobes and they are being considered part of the normal intestinal microbiota of humans and animals [19]. More
than 40 different species have already been described, with E. faecalis, E. faecium, E. cecorum, E. hirae and E. gallinarum being the most prevalent ones found in animals.
Enterococci are considered commensal microorganisms with worldwide spread [20], and for a long time they were considered harmless microorganisms, however, recently, they have been considered emerging and of great importance in the health area, mainly with the increasing incidence of nosocomial infections [20, 21, 22, 23, 24].
Currently, Enterococci are considered important opportunistic pathogens capable of causing infections in animals and humans [20, 25]. They are the third most prevalent pathogen in nosocomial infections [26].
Vancomycin is one of the main antimicrobials used in the treatment of Enterococcus infections, however, at the end of the 1980s, in Europe, the indiscriminate use of the glycopeptide avoparcin (analogous to vancomycin) as a growth promoter for production animals, led to the emergence of resistant strains, called vancomycin-resistant Enterococcus (VRE) [27, 28]. Since then, VRE has already been isolated in the environment, in domestic and production animals, food of animal origin, fish, free-living animals, water, hospitals, among others [29, 30, 31, 32, 33, 34, 35, 36].
The first VRE isolates from clinical samples were identified in England and France in 1988, being an isolate of E. faecium [37]. With the appearance of VRE strains in the hospital environment, the treatment of these microorganisms was limited, making them one of the most important multidrug-resistant bacteria in the world [26]. The only antimicrobials used in some situations for the treatment of VRE are: quinupristin/dalfopristin, linezolid, tigecycline and daptomycin. Meantime, resistance to these antimicrobials has already been described [38, 39].
In veterinary medicine, especially in small animals, there is not much data regarding VRE. Sfaciotte, et al. [14] divided a veterinary hospital in Brazil into 39 locations, and in 10 of them, there was the presence of VRE identified as Enterococcus faecalis and with the presence of the vanA gene. So far, nine different variations in the vancomycin resistance gene of Enterococci have been found, with the vanA, vanB and vanC genes being the most commonly found [40, 41].
The impact on public health caused by the transfer of the plasmid vanA gene between strains of Enterococcus isolated from humans and those isolated from animals is still unknown [42]. In addiction, it is already known that the transfer of this gene happens to microorganisms of different genera, since the vanA gene has been identified in strains of methicillin-resistant Staphylococcus aureus (MRSA) [43].
Methicillin Resistant Staphylococcus (MRS)
Staphylococci are Gram positive, catalase positive bacteria and comprise more than 40 different species. They are divided into two main groups: positive coagulase (PCS), mainly S. aureus and S. pseudintermedius; and negative coagulase (SCN), S. epidermidis and S. [44]. They are considered part of the normal microbiota of the skin and mucous membranes of animals and humans, and are commonly associated with opportunistic infections [45].
Staphylococcus pseudintermedius belongs to the “Staphylococcus intermedius group” (SIG group) and is an opportunistic pathogen, being considered the main pathogen isolated from dogs and cats [46, 47]. The SIG group (S. pseudintermedius, S. intermedius and S. delphini) can only be differentiated by molecular techniques, however, when it is not possible to use these techniques, all dog and cat isolates can be classified as S. pseudintermedius [46, 48].
The two main mechanisms of antimicrobial resistance in Staphylococcus are associated with resistance to the beta-lactam class, the first mechanism being an enzymatic inactivation encoded by the blaZ gene, usually plasmid, which confers resistance to penicillins [49]; and the second by the production of additional penicillin-binding protein (PBP2a or PBP2’), a low-affinity penicillin-binding protein encoded by the mecA and mecC genes [50], conferring resistance to all antimicrobials of the beta-lactam class [51], with the exception of fifth-generation cephalosporins (ceftaroline and ceftobiprole).
The blaZ gene has already been identified in Staphylococcus isolates of canine and feline origin [52], including strains of S. pseudintermedius methicillin resistant (MRSP) and methicillin sensitive strains (MRSS) [53, 54], leading to an increase in resistance to penicillin and ampicillin mainly [55].
The mecC gene was recently identified, and, so far, there are few positive isolates for this gene. However, as it has already been isolated in different animal species and in humans in contact with animals, it is believed that mecC is of animal origin [56, 57, 58]. Phenotypically, the mecC gene is characterized by being resistant to cefoxitin, but sensitive to oxacillin, while the mecA gene is resistant to both antimicrobials [57, 59].
Methicillin resistance is the most important antimicrobial resistance mechanism identified in Staphylococcus, which is characterized by the presence of mecA and mecC genes carried in a mobile “genetic island” called “Staphylococcal Cassette Chromosome mec” (SCCmec) [60]. The SCCmec elements are capable of harboring insertion sequences, plasmids and transposons, conferring resistance to other classes of antimicrobials [61].
In total, 13 different types of SCCmec and their variations have already been identified in S. aureus and S. pseudintermedius [62]. Several studies have already identified the mecA gene as part of the SCCmec elements of animal origin [63, 64, 65].
The first phenotypic strain of MRSP was isolated from healthy dogs and dogs with pyoderma in France in the late 1980s [66]. The first identification of the mecA gene in dogs occurred in the United States in 1999 [67] and, later, in 2005, in Europe [68]. Since then, MRSP isolates have been reported worldwide in veterinary clinics and hospitals, being recognized as a pathogen of great importance due to multidrug resistance and the difficulty in treating infections [43,48,], in particular in cases of nosocomial infections [53, 55].
Initially, Staphylococcus pseudintermedius was susceptible to most antimicrobials. However, from the year 2006, MRSP emerged as a pathogen resistant to almost all classes of antimicrobials used in dogs, making antibiotic therapy difficult in small animals [69, 70].
In addition to being isolated in animals with some type of infection, MRSP have already been isolated in healthy dogs and cats, mainly in isolates from the nostrils, oral mucosa and skin [71, 72], being found a prevalence of 1.5% to 10% [73, 74].
The study conducted by Abusleme, et al. in a school veterinary hospital in Chile, a total of 45 strains of coagulase positive Staphylococcus (CoPS) were obtained, eight from veterinary professionals, three from hospital surfaces and eight from owners and 26 from dogs. Nine of the strains (20%) were methicillin resistant and all of them carried the mecA gene with high genetic diversity. The study also points out that veterinarians have a high risk of harboring methicillin-resistant CoPS (25% versus 2.5% of owners). In a study carried out in a school veterinary hospital in Brazil, MRS was isolated throughout the hospital environment, from those with large circulation of animals to those where there was no animals circulation, such as in bathrooms and restaurant [14].
These data are important because, despite being rare, human infections by MRSP have already been reported with suspected transmission from dogs [75]. Nasal colonizations by MRSP have also been described in veterinarians and dog owners who have already had MRSP infections [76, 77, 78], in addition to environmental contamination [79].
The treatment of MRSP isolates is a challenge in veterinary medicine, since the main antimicrobials used to treat these multiresistant microorganisms are vancomycin and linezolid, but their use is questionable due to the use of these antimicrobials in humans and the fact that they can lead to strains resistant to them [53].
Methicillin-resistant Staphylococcus aureus (MRSA) is the main problem of nosocomial infection in humans [63], but in dogs its prevalence is lower than MRSP, while in cats there are still doubts about which of the two is more prevalent [80]. Both healthy dogs and cats can be colonized by MRSA [81, 82], but it is transient and can range from 0% to 3% in dogs and up to 4% in cats [83].
Other species of Staphylococcus, such as S. schleiferi and SCN, have also been described with the presence of the mecA gene, but it seems not to be relevant because they are less common in causing some type of infection. In a study carried out by Griffeth, et al. [84], a 2% prevalence of methicillin- resistant S. schleiferi (MRSS) was found in healthy dogs with skin inflammation, while Abraham, et al. [85] did not isolate MRSS in cats.
Extended Spectrum Beta-Lactamase (ESBL)
One of the main resistance mechanisms found in bacteria of the Enterobacterales order is the hydrolysis of the beta-lactam ring through enzymes, conferring resistance to antimicrobials of the beta-lactam class, including penicillins, cephalosporins and monobactams [86, 87], but they do not hydrolyze cephamycins and carbapenems, in addition to being inactivated by beta-lactamase inhibitors (clavulanic acid, sulbactam and tazobactam) [88]. These enzymes are called extended-spectrum beta-lactamase (ESBL), being divided into three main families, TEM, SHV and CTX-M [89, 90].
The genes that encode ESBL are located on conjugative plasmids or on integrons and can be transferred to other species of enterobacteria, further facilitating their propagation [91]. In recent decades, resistance to cephalosporins in much of the world has been considered a major public health threat [10, 92, 93], often being responsible for cases of nosocomial infections and even community infections [94].
The first enzyme conferring resistance to beta-lactams was of the TEM type, isolated in 1965 in Greece in a human patient called Temoneira, whose name gave rise to the TEM enzyme. This first enzyme, TEM-1, conferred resistance to penicillins and the first generation of cephalosporins, until in the 1980s, extended-spectrum variants of TEM-1 were identified [95].
The second enzyme identified was SHV (variable sulfhydryl), in 1979 [96], which, like TEM, other variants had already been discovered in the mid-80s, both having prominence in Enterobacterales resistant to the third generation of cephalosporins [97].
Currently, the most common ESBL is the CTX-M enzyme found mainly in E. coli strains [98]. It was identified for the first time by Matsumoto, et al. [99], in Japan, in the feces of a dog of laboratory. CTX-M are divided into five groups (1, 2, 8, 9 and 25), with CTX-M-15 (group 1) being one of the most frequently found in humans and animals [100]. It is being also associated with resistance to the class of fluoroquinolones and aminoglycosides [89]. However, in Brazil, a study showed a higher prevalence of CTX-M from groups 8 and 25 isolated from dogs and cats, showing that individual epidemiological studies are important to highlight the reality of each region [101].
Companion animals, mainly dogs and cats, are important sources of transmission of antimicrobial resistance genes due to their direct contact with humans. This fact represents a great risk to public health [102, 103], even more after identifying several types of ESBL-producing microorganisms [93, 100].
In veterinary medicine, the first clinical sample producing ESBL in companion animals was an E. coli, type SHV-12, isolated from a dog with urinary tract infection in Spain, in 1998 [104], followed by samples from dogs in Italy and Portugal [105, 106]. Different variants of CTX-M have already been isolated in clinical samples from dogs, as well as the enzymes TEM [107] and SHV [108]. In Brazil, a study showed the presence of these three gene families within a school veterinary hospital [14] and in healthy and sick animals admitted to the same hospital.
One of the main studies in the detection of ESBL in small animals was carried out in Germany, where 2700 samples were analyzed. CTX-M (1, 2, 14 and 15), were detected in isolates of E. coli, Salmonella enterica, Proteus mirabilis and Enterobacter cloaceae from wounds, urinary, respiratory, abdominal and bone infections [107].
Most studies carried out with small animals to identify ESBL-producing microorganisms target E. coli, however, these enzymes have already been described in other Enterobacterales, such as: Pantoea agglomerans, Morganella morganii, Providencia sp . [109], Citrobacter sp., Enterobacter sp., Klebsiella sp., Serratia marcescens, as well as non- Enterobacterales bacteria, such as Pseudomonas aeruginosa, Acinetobacter baumanii and Stenotrophomonas maltophilia in animals.
Although several ESBL isolates have already been identified in small animals, the main studies are focused on production animals, since the transfer of resistance genes between food of animal origin and humans has already been widely described [110, 111], but the prevalence, especially of CTX-M in small animals, is limited and requires further studies [112].
About nosocomial infection, ESBL are one of the main causes of this type of infection in humans, but the prevalence of this type of microorganism is still little known in veterinary medicine and the cases of nosocomial infections caused by them are even scarcer. In Brazil, Sfaciotte, et al. [101] collected rectal swabs from 106 animals (dogs and cats) admitted to a school veterinary hospital. 44.34% of which were colonized by ESBL-producing bacteria, showing the importance of disseminating these multiresistant microorganisms
Gram Negative Bacteria Producing Carbapenemases
Resistance to beta-lactams in Gram-negative bacilli has become an emerging problem worldwide in recent years, especially in cases of nosocomial infections, leading to increased patient mortality and increased hospitalization costs [113, 114].
Carbapenems are antimicrobials of the beta-lactam class and are considered the last choice for treatments. They are used even in ESBL-producing microorganisms, and are often used to treat human nosocomial infections [115, 116]. However, isolates resistant to carbapenems have already been reported, mainly microorganisms from the Enterobacterales order [117]. The mechanisms of resistance to these antimicrobials are associated with decreased membrane permeability, overexpression of beta-lactamase or expression of carbapenemases [118].
Several types of carbapenemases have already been described in Enterobacterales the most clinically important are: Klebsiella pneumoniae carbapenemases (KPC), zinc- dependent carbapenemases, also known as metallo-beta- lactamase (Verona integron – VIM, imipenemase – IMP, New Delhi – NDM), oxacilinases – 48 (OXA-48) [117, 119] and a cephalosporinase associated with porin loss also leading to resistance to beta-lactams (AmpC) [120].
Beta-lactamases are classified according to several criteria, two of them are the most widespread criteria (1) according to Ambler RP, et al. [121] who classifies beta- lactamase according to their enzymatic molecular structure into 4 groups, where classes A, C and D are called serine-β- lactamase, which have the amino acid serine in the active center of the enzyme; and class B, which are zinc-dependent and therefore called metallo-β-lactamase. (2) While the second criterion is according to the inhibition profile of the β-lactamases, known as the Bush group [122, 123]. Bush, et Group Bush Jacob1 Class Melecular2 Representative Enzymes Feature3 Substratos4 Inhibition by
1; 1e C CMY-2; CMY-37 AmpC SCfs Nao Nao
2be A TE<-3, SHV-2, CTX-M-2, 14,15 ESBL Oxiamino-Cfs e monobactans Sim Nao
2bre A TEM-50 IRT-ESBL Oxiamino-Cfs e monobactans Nao Nao
2de; 2df D OXA-11, 115, OXA- 23, 48 ESBL; Carbapenemases Oxiamino-Cfs e carbapenemicos Variable Nao
2f S GES-2, KPC-2,3 Carbapenemases Oxiamino-Cfs cefamicinas, monobactans e carbapenemicos Variable Nao
3a B(MBL) SPM-1, IMP-1, VIM-1, NDM-1 Carbapenemases Oxiamino-Cfs cefamicinas, e carbapenemicos, e monobactans Nao Sim al. [90] updated the classification scheme according to the two described criteria, which are represented in Table 1 & Figure 1.
Clavulanic acid EDTA
- 1Bush and Jacob (2010); 2Ambler (1980); 3ESBL (extended Spectrum β-Lactamase); extended spectrum β-Lactamase; IRT; TEM inhibitor resistance; MBL (Metallo-β-Lactomase); 4CFs: Cephalosporins Omiamino-Cfs; Broad-spectrum cephalosporins; 5EDTA
- (from English, Ethylenediamine tereaacetic acid): Ethylenediamine tetraacetic acid.
Table 1: Classification of the main beta-lactamases in Gram negative bacteria.
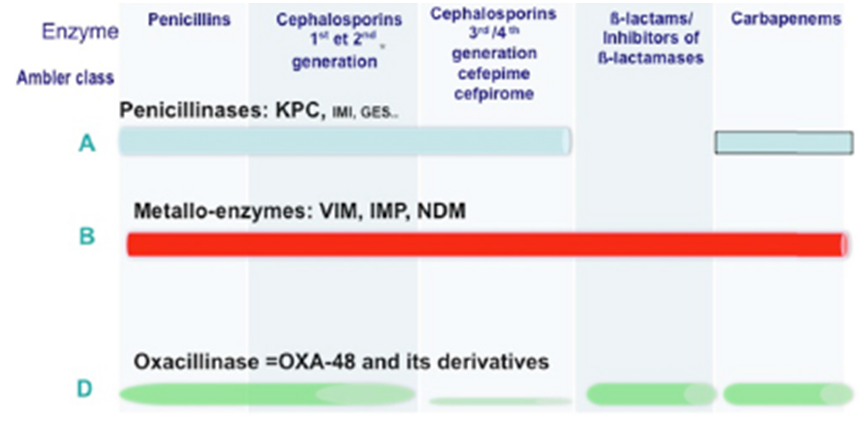
Despite name, the KPC enzyme is not restricted to Klebsiella pneumoniae, but to all enterobacteria, Pseudomonas aeruginosa and Acinetobacter baumanii, as it is normally found in plasmids and transposons, which facilitates its dissemination [122, 124].
In 1996, the first KPC (KPC-1) located in a plasmid was identified, in North Carolina, USA [125]. After that, plasmid KPC began to be isolated in several countries around the world, such as: China and Taiwan [126], Brazil [127] and different countries in Europe [128, 129, 130].
The first NDM (metallo-β-lactamase) described was in a Swedish patient hospitalized with a urinary tract infection after a trip to New Delhi, India, in 2009. The microorganism was a Klebsiella. pneumoniae resistant to almost all antimicrobials, except for tigecycline and colistin [131]. British authors determined that the large reservoir of these carbapenemases is located in India, Pakistan, Bangladesh and Sri-Lanka, through a study where they isolated NDM from bacteria found in water consumed by the population and in rainwater [132].
In addition to being isolated in cases of nosocomial infections, NDM have already been identified in community samples, mainly in E. coli, in cases of urinary infection and diarrhea, which makes it increasingly difficult to control this multidrug resistance mechanism, since controlling these microorganisms in the hospital is possible, but in the community, it is almost impossible [133].
Carbapenemase OXA is derived from ESBL genes and is located in integrons, transposons and/or insertion sequences housed in plasmids [134]. The identification of OXA-48 is relatively recent, and like other carbapenemases, it has already been described in different bacterial genera of the Enterobacterales and responsible for several cases of nosocomial infection [135].
Although not considered a carbapenemase, microorganisms with the presence of the ampC enzyme are considered beta-lactamase hyperproducers, however, the presence of this enzyme confers resistance to carbapenems when associated with an efflux system or alteration in the permeability of the outer membrane [136]. The main ampC with resistance to carbapenems are CTM-Y and FOX [137].
The number of already identified enzymes responsible for conferring resistance to carbapenems is very large and there are several studies isolating these enzymes in cases of infections in humans. In the other hand, in veterinary medicine, these data are little known, with few reports in production animals, in small animals [138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154], and more recently in the veterinary hospital environment in Brazil [14].
Abraham, et al. [85], isolated specimens of Salmonella Typhimurium from four domestic cats, one sick and three healthy that were in an animal shelter [155, 156, 157, 158, 159, 160, 161, 162]. The isolates harbored the blaIMP-4 gene. The genome sequencing revealed the acquisition of a multidrug-resistant plasmid that encoded resistance to nine antimicrobial classes, including carbapenems [163]. In Spain, Torralba-Gonzáles, et al. isolated a carbapenemase-producing Klebsiella pneumoniae by the blaVIM-1 gene in a dog rectal swab. As well as Franandes, et al. in Brazil, isolated a Pseudomonas aeruginosa ST233 with the blaVIM- gene 2 in a dog with otitis traced back to the dog’s owner who had been hospitalized about a month before the animal’s clinical signs manifested (this ST233 strain is internationally reported to be restricted to hospital settings) [164, 165, 166, 167, 168, 169, 170].
In a study carried out by Galarde López, et al. analyzing wastewater and treated wastewater from two hospitals in Mexico, 30 isolates were obtained, 26 (86.7%) of Klebsiella pneumoniae and 4 (13.3%) of of Klebsiella oxytoca, with 13 isolates carrying the blaKPC gene while one isolate was detected harboring both the blaKPC gene and the blaOXA-48 gene [171,s172].
Conclusion
The reality of nosocomial infections worldwide is a fact, with the costs associated with them and the number of deaths growing each year, mainly due to the indiscriminate and often wrong use of antimicrobials, a fact seen in the COVID-19 pandemic. However, in veterinary medicine these data are scarce, since research related to antimicrobial resistance is still punctual and often restricted to a few research groups around the world.
Despite research related to antimicrobial resistance, the reality of the clinical routine of animals does not seem to be in line with the world reality. There are few studies in veterinary medicine regarding nosocomial infections (some extremely punctual), In the other hand, these infections are already a reality in veterinary hospitals, mainly of small animals, and, unfortunately, they are most of the time neglected due to the difficulty of their traceability or even by the lack of knowledge.
References
-
Magill SS, Edwards JR, Bamberg W (2014) Multistate point-prevalence survey of health care-associated infections. N Engl J Med 370: 1198-1208.
-
Breathnach A (2013) Nosocomial infections and infection control**.** Medicine 41(11): 649-653.
-
Stull JW, Weese JS (2015) Hospital-Associated Infections in Small Animal Practice. Vet Clin Small Anim 45(2): 217- 233.
-
Ruple Czerniak A, Aceto HW, Bender JB (2013) Using syndromic surveillance to estimate baseline rates for healthcare-associated infections in critical care units of small animal referral hospitals. J Vet Intern Med 27(6): 392-399.
-
Benedict KM, Morley PS, Van Metre DC (2008) Characteristics of biosecurity and infection control programs at veterinary teaching hospitals. J Am Vet Med Assoc 233(5): 767-773.
-
Oliveira AC, Cardoso CS, Mascarenhas D (2010) Precauções de contato em Unidade de Terapia Intensiva: fatores facilitadores e dificultadores para adesão dos profissionais. Rev Esc Enferm USP 44(1): 161-165.
-
Scott RD (2009) The direct medical costs of healthcare- associated infections in U.S. hospitals and the benefits of prevention. Atlanta (GA): Centers for Disease Control and Prevention; Division of Healthcare Quality Promotion.
-
Weese JS (2012) Monitoring for surgical infection. In: Tobias KM, Johnston SA (Eds.), Veterinary Surgery, Small Animals, Elsevier Saunders, Missouri, USA, pp: 170-179.
-
Dallap SBL, Aceto H, Rankin SC (2010) Outbreak of salmonellosis caused by Salmonella enterica serovar Newport MDR-AmpC in a large animal veterinary teaching hospital. J Vet Intern Med 24(5): 1138-1146.
-
EFSA, ECDC (2015) EU Summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2013. EFSA J 13: 1-178.
-
Pelligand L, Hammond R, Rycroft A (2007) An investigation of the bacterial contamination of small animal breathing systems during routine use. Vet Anaesthes Analg 34(3): 190-199.
-
Ogeer Gyles JS, Mathews KA, Boerlin P (2006) Nosocomial infections and antimicrobial resistence in critical care medicine. J Vet Crit Care 16(1): 1-18.
-
Scheftel JM, Elchos BL, Cherry B (2010) Compendium of veterinary standard precautions for zoonotic disease prevention in veterinary personnel: National Association of State Public Health Veterinarians Veterinary Infection Control Committee. J Am Vet Med Assoc 237(12): 1403- 1422.
-
Sfaciotte RAP, Ricardo AP (2021) Detection of the main multiresistant microorganisms in the environment of a teaching veterinary hospital in Brazil. Pesquisa Veterinária Brasileira 41: e06706.
-
Ghosh A, Kukanich KS, Brown CE (2012) Resident cats in small animal veterinary hospitals carry multidrug resistant enterococci and are likely involved in crosscontamination of the hospital environment. Frontiers in Microbiol 3: 1-14.
-
Girling SJ, Fraser MA (2009) Bacterial carriage of computer keyboards in veterinary practices in Scotland. Vet Record 165(1): 26-27.
-
Sievert DM, Ricks P, Edwards JR, Schneider A, Patel J, et al. (2013) Antimicrobial-resistant pathogens associated with healthcare-associated infections: summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009- 2010. Infect Control Hosp Epidemiol 34(1): 1-14.
-
CDC (2013) Vital signs: carbapenemresistant Enterobacteriaceae. Morb Mortal Wkly Rep 62(9): 165- 170.
-
Shepard BD, Gilmor MS (2002) Differential expression of virulence-related genes in Enterococcus faecalis in response to biological cues in serum and urine. Infection and Immunity 70(8): 4344-4352.
-
Espinosa Gongora C, Shah SQA, Jessen LR, Bortolaia V, Langebæk R, et al. (2015) Quantitative assessment of faecal shedding of b-lactam-resistant Escherichia coli and enterococci in dogs. Veterinary Microbiology 181(3- 4): 298-302.
-
Khan HA, Ahma A, Mehboob R (2015) Nosocomial infections and their control strategies. Asian Pacific Journal of Tropical Biomedicine 5(7): 509-514.
-
Gilmore MS, Clewell DB, Ike Y, Shankar N (2014) Enterococci from commensals to leading causes of drug resistant infection. Eye and Ear Infirmary, Boston, Massachusetts.
-
Marcade G, Micol JB, Jacquier H, Raskine L, Donay JL, et al. (2014) Outbreak in a haematology unit involving an unusual strain of glycopeptideresistant Enterococcus faecium carrying both vanA and vanB genes**.** J Antimicrob Chemother 69(2): 500-505.
-
Arias CA, Murray BE (2012) The rise of the Enterococcus: beyond vancomycin resistance. Nature Reviews Microbiology 10(4): 266-278.
-
Leavis HL, Bonten MJ, Willems RJ (2006) Identification of high-risk enterococcal clonal complexes: global dispersion and antibiotic resistance. Current Opinion in Microbiology 9(5): 454-460.
-
ECDC (2018) Antimicrobial Resistance Surveillance in Europe 2017. Annual report of the European Antimicrobial resistance Surveillance Network (EARS- net) 2017, Stockholm, Sweden, pp: 1-97.
-
Bates J (1997) Epidemiology of vancomycin-resistant enterococci in the community and the relevance of farm animals to human infection. J Hosp Infect 37(2): 89-101.
-
Rice LB (2013) The complex dynamics of antimicrobial activity in the human gastrointestinal tract. Trans Am Clin Climatol Assoc 124: 123-132.
-
Torres C, Tenorio C, Portillo A, García M, Martínez C, et al. (2003) Intestinal colonization by vanA- or vanB2- containing enterococcal isolates of healthy animals in Spain. Microb. Drug Resist 9(S1): 47-52.
-
Poeta P, Costa D, Rodrigues J, Torres C (2005) Study of faecal colonization by vanA-containing Enterococcus strains in healthy humans, pets, poultry and wild animals in Portugal. J Antimicrob Chemother 55(2): 278-280.
-
Roberts MC, Soge OO, Giardino MA, Mazengia E, Ma G, et al. (2009) Vancomycin-resistant Enterococcus spp. in environments from the West Coast of the USA. J Appl Microbiol 107(1): 300-307.
-
Araujo C, Torres C, Goncalves A, Carneiro C, López M, et al. (2011) Genetic detection and multilocus sequence typing of vanA-containing Enterococcus strains from mullets fish (Liza ramada). Microb Drug Resist 17(3): 357-361.
-
Iversen A, Kuhn I, Franklin A, Möllby R (2012) High prevalence of vancomycin-resistant enterococci in Swedish sewage. Appl Environ Microbiol 68(6): 2838- 2842.
-
Getachew Y, Hassan L, Zakaria Z, Abdul Aziz S (2013) Genetic variability of vancomycin- resistant Enterococcus faecium and Enterococcus faecalis isolated from humans chickens, and pigs in Malaysia. Appl Environ Microbiol 79(15): 4528-4533.
-
Oravcova V, Ghosh A, Zurek L, Bardon J, Guenther S, et al. (2013) Vancomycin-resistant enterococci in rook (Corvus frugilegus) wintering throughout Europe. Environ Microbiol 15(2): 548-556.
-
Lozano C, Gonzalez-Barrio D, Camacho MC, Lima-Barbero JF, de la Puente J, et al. (2016) Characterization of faecal vancomycin-resistant enterococci with acquired and intrinsic resistance mechanisms in wild animals, Spain. Microb Ecol 72(4): 813-820.
-
O’driscoll T, Crank CW (2015) Vancomycin-resistant enterococcal infections: epidemiology clinical manifestations, and optimal management. Infect Drug Resist 8: 217-230.
-
Werner G, Gfrörer S, Fleige C, Witte W, Klare I (2008) Tigecycline-resistant Enterococcus faecalis strain isolated from a German ICU patient. Journal of Antimicrobial Chemotherapy 6(5): 1182-1183.
-
Arias CA, Panesso D, Mcgrath DM, Qin X, Mojica MF, et al. (2011) Genetic basis for in vivo daptomycin resistance in enterococci. New England Journal of Medicine 365(10): 892-900.
-
Fisher K, Phillips C (2009) The ecology, epidemiology and virulence of Enterococcus. Microbiology Washington, USA, 155(Pt6): 1749-1757.
-
Lebreton F, Depardieu F, Bourdon N, Fines-Guyon M, Berger P, et al. (2011) D-Alad- Ser VanN-type transferable vancomycin resistance in Enterococcus faecium. Antimicrobial Agents and Chemotherapy 55(10): 4606- 4612.
-
Sletvold H, Johnsen PJ, Wikmark OG, Simonsen GS, Sundsfjord A, et al. (2010) Tn1546 is part of a larger plasmid-encoded genetic unit horizontally disseminated among clonal Enterococcus faecium lineages**.** J Antimicrob Chemother 65(9): 1894-1906.
-
Sfaciotte RAP, Coronel LG, Osaki SC, Rezler WS (2015) Gram-positive bacterial resistant strains of interest in animal and public health. Brazilian Journals in Veterinary Medicine and Animal Science 36(4): 2693-2712.
-
Bond R, Loeffler A (2012) What’s happened to Staphylococcus intermedius? Taxonomic revision and emergence of multi-drug resistance. J Small Anim Pract 53(3): 147-154.
-
Malachowa N, Deleo FR (2010) Mobile genetic elements of Staphylococcus aureus. Cell Mol Life Sci 67 (18): 3057- 3071.
-
Devriese LA, Vancanneyt M, Baele M, Vaneechoutte M, De Graef E, et al. (2005) Staphylococcus pseudintermedius sp. nov., a coagulase-positive species from animals. International Journal of Systematic and Evolutionary Microbiology 55(Pt 4): 1569-1573.
-
Gronthal T, Eklund M, Thomson K, Piiparinen H, Sironen T, et al. (2017) Antimicrobial resistance in Staphylococcus pseudintermedius and the molecular epidemiology of methicillin-resistant S. pseudintermedius in small animals in Finland. J Antimicrob Chemother 72(4): 1021-1030.
-
Bannoehr J, Guardabassi L (2012) Staphylococcus pseudintermedius in the dog: taxonomy, diagnostics, ecology, epidemiology and pathogenicity. Vet Dermatol 23(4): 253-266.
-
Lowy FD (2003) Antimicrobial resistance: the example of Staphylococcus aureus. J Clin Invest 111(9): 1265- 1273.
-
Castellano-Gonzalez MJ, Perozo-Mena AJ, Vivas-Veja RL, Ginestre-Pérez MM, Rincón-Villalobos GC (2009) Molecular and phenotypical typification of methicillin resistant Staphylococcus aureus (MRSA) strains in a university hospital. Rev Chilena Infectol 26(1): 39-48.
-
Clinical And Laboratory Standards Institute-Clsi (2008) Performance Standards For Antimicrobial Disk And Dilution Susceptibility Test For Bacteria Isolated From Animals Approved Standard. In: 3rd (Edn.), M31-A3. Wayne: Clinical and Laboratory Standards Institute 28(8): 103.
-
Malik S, Christensen H, Peng H, Barton MD (2007) Presence and Diversity Of The Beta-Lactamase Gene In Cat And Dog Staphylococci. Vet Microbiol 123(1-3): 162- 168.
-
Perreten V, Kadlec K, Schwarz S, Grönlund Andersson U, Finn M, et al. (2010) Clonal Spread of Methicillin- Resistant Staphylococcus Pseudintermedius in Europe and North America: An International Multicentre Study. J Antimicrob Chemother 65(6): 1145-1154.
-
Gómez Sanz E, Torres C, Lozano C, Sáenz Y, Zarazaga M, et al. (2011) Detection And Characterization Of Methicillin- Resistant Staphylococcus Pseudintermedius In Healthy Dogs In La Rioja, Spain. Comp Immunol Microbiol Infect 34(5): 447-453.
-
Kadlec K, Schwarz S (2012) Antimicrobial Resistance of Staphylococcus Pseudintermedius. Vet Dermatol 23(4): 276-282.
-
Garcia Alvarez L, Webb CR, Holmes MA (2011) A Novel Field-Based Approach To Validate The Use Of Network Models For Disease Spread Between Dairy Herds. Epidemiology And Infection, Cambridge, 139(12): 1863- 1874.
-
Paterson GK, Larsen AR, Robb A, Edwards GE, Pennycott TW, et al. (2012) The Newly Described Meca Homologue, Meca(Lga251), Is Present In Methicillin-Resistant Staphylococcus Aureus Isolates From A Diverse Range Of Host Species. Journal Of Antimicrobial Chemotherapy, London, UK 67(12): 2809-2813.
-
Petersen A, Stegger M, Heltberg O, Christensen J, Zeuthen A, et al. (2013) Epidemiology Of Methicillin-Resistant Staphylococcus Aureus Carrying The Novel Mecc Gene In Denmark Corroborates A Zoonotic Reservoir With Transmission To Humans. Clin Microbiol Infect 19(1): 16-22.
-
Cartwright EJP, Paterson GK, Raven KE, Ewan M Harrison, et al. (2013) Use Of Vitek 2 Antimicrobial Susceptibility Profile To Identify Mecc In Methicillin-Resistant Staphylococcus Aureus. Journal Of Clinical Microbiology 51(8): 2732-2734.
-
Kania SA, Williamson NL, Frank LA, Wilkes RP, Jones RD, et al. (2004) Methicillin Resistance Of Staphylococci Isolated From The Skin Of Dogs With Pyoderma. Am J Vet Res 65(9): 1265-1268.
-
Shore AC, Coleman DC (2013) Staphylococcal Cassette Chromosome Mec: Recent Advances And New Insights. International Journal Of Medical Microbiology 303(6-7): 350-359.
-
International Working Group on The Classification Of Staphylococcal Cassette Chromosome Elements -Iwg- Scc (2009) Classification Of Staphylococcal Cassette Chromosome Mec (Sccmec): Guidelines For Reporting Novel Sccmec Elements. Antimicrobial Agents and Chemotherapy 53(12): 4961-4967.
-
Weese JS, Van Duijkeren E (2010) Methicillinresistant Staphylococcus Aureus and Staphylococcus Pseudintermedius In Veterinary Medicine. Veterinary Microbiology 140(3-4): 418-429.
-
Fitzgerald JR (212) Livestock Associated Staphylococcus Aureus: Origin, Evolution And Public Health Threat. Trends Microbiol 20(4): 192-198.
-
Pantosti A (2012) Methicillin Resistant Staphylococcus Aureus Associated With Animals And Its Relevance To Human Health. Front Microbiol 3: 127.
-
Pellerin JL, Bourdeau P, Sebbag H, Person JM (1988) Epidemiosurveillance Of Antimicrobial Compound Resistance Of Staphylococcus Intermedius Clinical Isolates From Canine Pyodermas. Comp Immunol Microbiol Infect Dis 21(2): 115-133.
-
Gortel K, Campbell KL, Kakoma I, Whittem T, Schaeffer DJ, et al. (1999) Methicillin Resistance Among Staphylococci Isolated From Dogs. Am J Vet Res 60(12): 1526-1530.
-
Loeffler A, Linek M, Moodley A, Guardabassi L, Sung JML, et al. (2007) First Report Of Multiresistant, Mecapositive Staphylococcus Intermedius In Europe: 12 Cases From A Veterinary Dermatology Referral Clinic In Germany. Vet Dermatol 18(6): 412-421.
-
Cain CL (2013) Antimicrobial Resistance In Staphylococci In Small Animals. Vet Clin Small Anim 43(1): 19-40.
-
Van Duijkeren E, Kamphuis M, Van Der Mije IC, Laarhoven LM, Duim B, et al. (2011) Transmission Of Methicillinresistant Staphylococcus Pseudintermedius Between Infected Dogs And Cats And Contact Pets, Humans And The Environment In Households And Veterinary Clinics. Veterinary Microbiology Amsterdan 150(3-4): 338-343.
-
Gingrich EN, Kurt T, Hyatt DR, Lappin MR, Ruch Gallie R (2011) Prevalence of Methicillin-Resistant Staphylococci In Northern Colorado Shelter Animals. J Vet Diagn Invest 23(5): 947-950.
-
Vanderhaeghen W, Van De Velde E, Crombe F, Polis I, Hermans K, et al. (2011) Screening For Methicillinresistant Staphylococci In Dogs Admitted To A Veterinary Teaching Hospital. Res Vet Sci 93(1): 133-
-
Rubin JE, Chirino Trejo M (2011) Prevalence, Sites Of Colonization, And Antimicrobial Resistance Among Staphylococcus Pseudintermedius Isolates From Healthy Dogs In Saskatoon, Canada. J Vet Diagn Invest 23(2): 351-354.
-
Couto N, Pomba C, Moodley A, Guardabassi L (2011) Prevalence Of Methicillin-Resistant Staphylococci Among Dogs And Cats At A Veterinary Teaching Hospital In Portugal. Vet Rec 169(3): 72.
-
Stegmann R, Burnens A, Maranta CA, perrenten V (2010) Human Infection Associated With Methicillin-Resistant Staphylococcus Pseudintermedius St71. J Antimicrob Chemother 65(9): 2047-2048.
-
Laarhoven LM, De Heus P, Van Luijn J, Duim B, Wagenaar JA, et al. (2011) Longitudinal Study On Methicillinresistant Staphylococcus Pseudintermedius In Households. Plos One 6(11): 27788.
-
Paul NC, Moodley A, Ghibaudo G, Guardabassi L (2011) Carriage Of Methicillin-Resistant Staphylococcus Pseudintermedius In Small Animal Veterinarians: Indirect Evidence of Zoonotic Transmission. Zoonoses Public Health 58(8): 533-539.
-
Sasaki T, Kikucki K, Tanaka Y, Takahashi N, Kamata S, et al. (2007) Reclassification Of Phenotypically Identified Staphylococcus Intermedius Strains. J Clin Microbiol 45(9): 2770-2778.
-
Ishihara K, Shimokubo N, Sakagami A, Ueno H, Muramatsu Y, et al. (2010) Occurrence Of Molecular Characteristics of Methicillin-Resistant Staphylococcus Aureus And Methicillin-Resistant Staphylococcus Pseudintermedius In An Academic Veterinary Hospital. Appl Environ Microbiol 76(15): 5165-5174.
-
Fazakerley J, Nuttall T, Sales D, Schmidt V, Carter SD, et al. (2009) Staphylococcal Colonization Of Mucosal And Lesional Skin Sites In Atopic And Healthy Dogs. Vet Dermatol 20(3): 179-184.
-
Morris DO, Lautenbach E, Zaoutis T, Leckerman K, Edelstein PH, et al. (2012) Potential For Pet Animals To Harbour Methicillin-Resistant Staphylococcus Aureus When Residing With Human Mrsa Patients. Zoonoses Public Health 59(4): 286-293.
-
Bender JB, Waters KC, Nerby J, Olsen KE, Jawahir S, et al. (2012) Methicillin-Resistant Staphylococcus Aureus (Mrsa) Isolated From Pets Living In Households With Mrsa-Infected Children. Clin Infect Dis 54(3): 449-450.
-
Bender JB, Schiffman E, Hiber L, Gerads l, Olsen K (2012) Recovery Of Staphylococci From Computer Keyboards In A Veterinary Medical Centre And The Effect Of Routine Cleaning. Vet Rec 170(16): 414-416.
-
Griffeth GC, Morris DO, Abraham JL, Shofer FS, Rankin SC (2008) Screening For Skin Carriage Of Methicillin- Resistant Coagulase Positive Staphylococci And Staphylococcus Schleiferi In Dogs With Healthy And Inflamed Skin. Vet Dermatol 19(3): 142-149.
-
Abraham JL, Morris DO, Griffeth GC, Shofer FS, Rankin SC (2007) Surveillance Of Healthy Cats And Cats With Inflammatory Skin Disease For Colonization Of The Skin By Methicillin-Resistan Coagulase-Positive Staphylococci And Staphylococcus Schleiferi Ssp. Schleiferi. Vet Dermatol 18(4): 252-259.
-
European Centre for Disease Prevention and Control- Ecdc (2011) Annual Epidemiological Report Reporting On Lligence Data. Annual Epidemiological Report On Communicable Diseases In Europe. Stockholm: Ecdc pp: 227.
-
Pfeifer Y, Cullik A, Witte W (2010) Resistance To Cephalosporins And Carbapenems In Gram Negative Bacterial Pathogens. Int J Med Microbiol 300(6): 371- 379.
-
Lee JH, Bae IK, Lee SH (2012) New Definitions Of Extended-Spectrum Beta-Lactamase Conferring Worldwide Emerging Antibiotic Resistance. Med Res Ver 32(1): 216-232.
-
Coque TM, Novais A, Carattoli A (2008) Dissemination of Clonally Related Escherichia Coli Strains Expressing Extended-Spectrum Lactamase Ctx-M-15. Emerg Infect Dis 14(2): 195-200.
-
Bush K Jacob GA (2010) Updated Functional Classification Of Β-Lactamases. Antimicrobial Agents And Chemotherapy 54(3): 969-976.
-
Carattoli A (2013) Plasmids and the spread of resistance. Int J Med Microbiol 303(6-7): 298-304.
-
Leverstein-Van Hall MA, Dierikx CM, Cohen Stuart J, Voets GM, Van den Munckhof MP, et al. (2011) Dutch patients, retail chicken meat and poultry share the same ESBL genes, plasmids and strains. Clin Microbiol Infect 17(6): 873-880.
-
Woodford N, Turton JF, Livermore DM (2011) Multiresistant gram-negative bacteria: the role of high- risk clones in the dissemination of antibiotic resistance. FEMS Microbiol Rev 35(5): 736-755.
-
Berglund B (2015) Environmental dissemination of antibiotic resistance genes and correlation to anthropogenic contamination with antibiotics. Infect Ecol Epidemiol 5: 28564.
-
Paterson DL, Bonomo RA (2005) Extended-spectrum β lactamases: a clinical update. Clinical Microbiology Reviews 18(4): 657-686.
-
Matthew M, Hedges RW, Smith JT (1979) Types of beta- lactamase determined by plasmids in gram-negative bacteria. J Bacteriol 138(3): 657-662.
-
Pitout JD (2012) Extraintestinal pathogenic Escherichia coli: an update on antimicrobial resistance, laboratory diagnosis and treatment. Expert Rev Anti Infect Ther 10(10): 1165-1176.
-
Bevan ER, Jones AM, Hawkey PM (2017) Global epidemiology of CTX-M β-lactamases: temporal and geographical shifts in genotype. J Antimicrob Chemother 72(8): 2145-2155.
-
Matsumoto Y, Ikeda F, Kamimura T, (1988) Novel plasmid-mediated -lactamase from Escherichia coli that inactivates oxyiminocephalosporins. Antimicrob Agents Chemother 32: 1243-1246.
-
Ewers C, Bethe A, Wieler LH, Guenther S, Stamm I, et al. (2011) Companion animals: a relevant source of extended-spectrum beta-lactamase-producing fluoroquinolone-resistant Citrobacter freundii. Int J Antimicrob Agents 37(1): 86-87.
-
Sfaciotte RAP, Parussolo L, Melo FD, Wildemann P, Bordignon G, et al. (2021) Identification and characterization of multidrug-resistant extended- spectrum beta-lactamase-producing bacteria from healthy and diseased dogs and cats admitted to a veterinary hospital in Brazil. Microb Drug Resist 27(6): 855-864.
-
De Briyne N, Atkinson J, Pokludová L, Borriello SP (2014) Antibiotics used most commonly to treat animals in Europe. Vet Rec 175(13): 325.
-
Hordijk J, Schoormans A, Kwakernaak M, (2013) High prevalence of fecal carriage of extended spectrum -lactamase/AmpC-producing Enterobacteriaceae in cats and dogs. Front Microbiol 4: 242.
-
Teshager T, Domínguez L, Moreno MA, Saénz Y, Torres C, et al. (2000) Isolation of an SHV-12 beta- lactamase-producing Escherichia coli strain from a dog with recurrent urinary tract infections. Antimicrob Agents Chemother 44(12): 3483-3484.
-
Féria C, Ferreira E, Correia JD, Gonçalves J, Caniça M, et al. (2002) Patterns and mechanisms of resistance to beta-lactams and beta-lactamase inhibitors in uropathogenic Escherichia coli isolated from dogs in Portugal. J Antimicrob Chemother 49(1): 77-85.
-
Carattoli A, Lovari S, Franco A, Cordaro G, Di Matteo P, et al. (2005) Extended-spectrum beta-lactamases in Escherichia coli isolated from dogs and cats in Rome Italy, from 2001 to 2003. Antimicrob Agents Chemother 49(2): 833-835.
-
Dierikx CM, Van Duijkeren E, Schoormans AH, Van Essen-Zandbergen A, Veldman K, et al. (2012) Occurrence and characteristics of extended-spectrum-β- lactamase and AmpC producing clinical isolates derived from companion animals and horses. J Antimicrob Chemother 67(6): 1368-1374.
-
Martins LR, Pina SM, Simões RL, Matos AJ, Rodrigues P, et al. (2013) Common phenotypic and genotypic antimicrobial resistance patterns found in a case study of multiresistant E. coli from cohabitant pets, humans, and household surfaces. J Environ Health 75(6): 74-81.
-
Zhao WH, Hu ZQ (2013) Epidemiology and genetics of CTX-M extended-spectrum -β-lactamases in Gram- negative bacteria. Crit Rev Microbiol 39: 79-101.
-
Seiffert SN, Hilty M, Perreten V, Endimiani A (2013) Extended-spectrum cephalosporin-resistant gram- negative organisms in livestock: an emerging problem for human health?. Drug Resist Update 16(1-2): 22-45.
-
Tamang MD, Nam HM, Kim SR, Chae MH, Jang GC, et al. (2013) Prevalence and molecular characterization of CTX-M beta-lactamase-producing Escherichia coli isolated from healthy swine and cattle. Foodborne Pathog Dis 10(1): 13-20.
-
Rubin JE, Pitout JD (2014) Extended-spectrum b-lactamase, carbapenemase and AmpC producing Enterobacteriaceae in companion animals. Vet Microbiol 170(1-2): 10-18.
-
Genc O, Aksu E, Gulcan A (2016) The identification of carbapenemase types in Enterobacteriaceae by using molecular assay and phenotyping confirmation tests. J Microbiol Methods 125: 8-11.
-
Robledo IE, Aquino EE, Vázquez GJ (2011) Detection of the KPC gene in Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Acinetobacter baumannii during a PCR-based nosocomial surveillance study in Puerto Rico. Antimicrob Agents Chemother 55(6): 2968- 2970.
-
Spellberg B, Blaser M, Guidos RJ, Boucher HW, Bradley JS, et al. (2011) Combating antimicrobial resistance: policy recommendations to save lives. Infectious Diseases Society of America (IDSA) Clin Infect Dis 52(5): 397-428.
-
WHO (2011) Critically Important Antimicrobials for Human Medicine. WHO Library Cataloguing-in- Publication Data 3rd Revision.
-
Nordmann P, Naas T, Poirel L (2011) Global spread of Carbapenemase-producing Enterobacteriaceae. Emerg Infect Dis 17(10): 1791-1798.
-
Thomson KS (2010) Extended-spectrum β-lactamase, AmpC, and carbapenemase issues. J Clin Microbiol 48(4): 1019-1025.
-
Tzouvelekis LS, Markogiannakis A, Psichogiou M, Tassios PT, Daikos GL, et al. (2012) Carbapenemases in Klebsiella pneumoniae and other Enterobacteriaceae: an evolving crisis of global dimensions. Clin Microbiol Rev 25(4): 682-707.
-
Nordmann P, Gniadkowski M, Giske CG, Poirel L, Woodford N, et al. (2012) Identifi cation and screening of carbapenemaseproducing Enterobacteriaceae. Clin Microbiol Infect 18(5): 432-438.
-
Ambler R (1980) The structure of beta-lactamase. Philos Trans R Soc Lond B Biol Sci 289(1036): 321-331.
-
Nordmann P (2014) Carbapenemase-producing Enterobacteriaceae: Overview of a major public health challenge. Med Mal Infect 44(2): 51-56.
-
Bush K, Jacoby GA, Medeiros AA (1995) A functional Classification Scheme for β- Lactamases and Its Correlation with Molecular Structure. Antimicrob Agents Chemother 39(6): 1211-1233.
-
Curiao T, Morosini MI, Ruiz-Garbajosa P, Robustillo A, Baquero F, et al. (2010) Emergence of blaKPC-3- Tn4401a associated with a pKPN3/4-like plasmid within ST384 and ST388 Klebsiella pneumoniae clones in Spain. J Antimicrob Chemother 65(8): 1608-1614.
-
Yigit H, Queenan AM, Anderson GJ, Domenech- Sanchez A, Biddle JW, et al. (2001) Novel carbapenemhydrolyzing β-lactamase, KPC-1, from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob Agents Chemother 45(4): 1151-1161.
-
Chung KP, Tseng SP, Huang YT, Tsai TH, Teng LJ, et al. (2011) Arrival of Klebsiella pneumoniae carbapenemase (KPC)-2 in Taiwan. J Antimicrob Chemother 66(5): 1182- 1184.
-
Chang MR, Biberg CA, Lopes FA, Tetila AF, Pignatari AC, et al. (2013) The first report of infection with Klebsiella pneumoniae carrying the bla kpc gene in State of Mato Grosso do Sul, Brazil. Rev Soc Bras Med Trop 46(1): 114-115.
-
Cantón R, González-Alba JM, Galán JC (2012) CTX-M enzymes: origin and diffusion. Front Microbiol 3: 110.
-
Babouee B, Widmer AF, Dubuis O, Ciardo D, Droz S, et al. (2011) Emergence of four cases of KPC-2 and KPC-3-carrying Klebsiella pneumoniae introduced to Switzerland, 2009-10. Euro Surveill 16(11): 19817.
-
Wendt C, Schütt S, Dalpke AH, Konrad M, Mieth M, et al. (2010) First outbreak of Klebsiella pneumoniae carbapenemase (KPC)- producing K. pneumoniae in Germany. Eur J Clin Microbiol Infect Dis 29(5): 563-570.
-
Yong D, Toleman MA, Giske CG, Cho HS, Sundman K, et al. (2009) Characterization of a new metallo-beta- lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother 53(12): 5046-5054.
-
Walsh TR, Weeks J, Livermore DM, Toleman MA (2011) Dissemination of NDM-1 positive bacteria in the New Delhi environment and its implications for human health: an environmental point prevalence study. Lancet Infect Dis 11(5): 355-362.
-
Poirel L, Benouda A, Hays C, Nordmann P (2011) Emergence of NDM-1- producing Klebsiella pneumoniae in Morocco. J Antimicrob Chemother 66(12): 2781-2783.
-
Walther-Rasmussen J, Hoiby N (2007) Class A carbapenemases. J Antimicrob Chemother 60(3): 470- 482.
-
Poirel L, Bonnin RA, Nordmann P (2011) Analysis of the resistome of a multidrug-resistant NDM-1-producing Escherichia coli strain by high-throughput genome sequencing. Antimicrob Agents Chemother 55(9): 4224- 4229.
-
Livermore DM, Woodford N (2006) The β-lactamase threat in Enterobacteriaceae, Pseudomonas and Acinetobacter. Trends Microbiol 14(9): 413-420.
-
Jacoby GA (2009) AmpC β-Lactamases. Clin Microbiol Rev 22(1): 161-182.
-
EFSA (2013) Scientific Opinion on Carbapenem resistance in food animal ecosystems. EFSA J 11(12): 3501.
-
Fischer J, Rodríguez I, Schmoger S, Friese A, Roesler U, et al. (2013) Salmonella enterica subsp. enterica producing VIM-1 carbapenemase isolated from livestock farms. J Antimicrob Chemother 68(2): 478-480.
-
Stolle I, Prenger-Berninghoff E, Stamm I, Scheufen S, Hassdenteufel E, et al. (2013) Emergence of OXA- 48 carbapenemase-producing E. coli and Klebsiella pneumoniae in dogs. J Antimicrob Chemother 68(12): 2802-2808.
-
Beaches Acht (2000) Beaches Environmental Assessment and Costal Health Act of 2000. United States Statutes at Large 114: 870-877.
-
Bemis DA, Jones RD, Frank LA, Kania SA (2009) Evaluation of susceptibility test breakpoints used to predict mecA-mediated resistance in Staphylococcus pseudintermedius isolated from dogs. J Vet Diagn Invest 21(1): 53-58.
-
Ben Belgacem Z, Abriouel H, Ben Omar N, Lucasb R, Gálvez A, et al. (2010) Antimicrobial activity, safety aspects, and some technological properties of bacteriocinogenic Enterococcus faecium from artesanal Tunisian fermented meat. Journal of Food Protection 21(4): 462-470.
-
Biswas PP, Dey S, Adhikari L, Sen A (2014) Virulence markers of vancomycin resistant enterococci isolated from infected and colonized patients. J Glob Infect Dis 6(4): 157-163.
-
Bortolaia V, Hansen KH, Nielsen CA, Fritsche TR, Guardabassi L, et al. (2014) High diversity of plasmids harbouring blaCMY-2 among clinical Escherichia coli isolates from humans and companion animals in the upper Midwestern USA. J Antimicrob Chemother 69(6): 1492-1496.
-
Cantón R, Akóva M, Carmeli Y, Giske CG, Glupczynski Y, et al. (2012) Rapid evolution and spread of carbapenemases among Enterobacteriaceae in Europe. Clin Microbiol Infect 18(5): 413-431.
-
Eckert C, Gautier V, Saladin-Allard M, Hidri N, Verdet C, et al. (2004) Dissemination of CTX-M-type beta- lactamases among clinical isolates of Enterobacteriaceae in Paris, France. Antimicrob Agents Chemother 48(4): 1249-1255.
-
Ghosh A, Dowd SE, Zurek L (2011) Dogs Leaving the ICU Carry a Very Large Multi-Drug Resistant Enterococcal Population with Capacity for Biofilm Formation and Horizontal Gene Transfer. PLoS One 6(7): e22451.
-
Hidron AI, Edwards JR, Patel J, Horan TC, Sievert DM, et al. (2008) NHSN annual update: antimicrobial- resistant pathogens associated with healthcare- associated infections: annual summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2006-2007. Infect Control Hosp Epidemiol 29(11): 996-1011.
-
Hiramatsu K (2001) Vancomycin-resistant Staphylococcus aureus: a new model of antibiotic resistance. Lancet Infect Dis 1(3): 147-155.
-
Hollenbeck BL, Rice LB (2012) Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 3(5): 421-433.
-
Jacoby G, Bush K (2014) B-lactamase classification and amino acid sequences for TEM, SHV and OXA extended-spectrum and inhibitor resistant enzymes (Lahey Clinic).
-
Jones RD, Kania SA, Rohrbach BW, Frank LA, Bemis DA (2007) Prevalence of oxacillin- and multidrugresistant staphylococci in clinical samples from dogs: 1,772 samples (2001- 2005). J Am Vet Med Assoc 230(2): 221- 227.
-
Kawakami T, Shibata S, Murayama N, Nagata M, Nishifuji K, et al. (2010) Antimicrobial susceptibility and methicillin resistance in Staphylococcus pseudintermedius and Staphylococcus schleiferi subsp. coagulans isolated from dogs with pyoderma in Japan. J Vet Med Sci 72(12): 1615-1619.
-
Kjellman EE, Slettemeås JS, Small H, Sunde M (2015) Methicillin‐resistant Staphylococcus pseudintermedius (MRSP) from healthy dogs in Norway- occurrence, genotypes and comparison to clinical MRSP. Microbiologyopen 4(6): 857-866.
-
Knight GM, Budd EL, Whitney L, Thornley A, Al- Ghusein H, et al. (2012) Shift in dominant hospital- associated methicillin-resistant Staphylococcus aureus (HA-MRSA) clones over time. J Antimicrob Chemother 67(10): 2514-2522.
-
Kramer A, Schwebke I, Kampf G (2006) How long do nosocomial pathogens persist on inanimate surfaces? A systematic review. BMC Infect Dis 16(6): 130.
-
Lee SM, Ender M, Adhikari R, Smith JM, Cook GM, et al. (2007) Fitness cost of staphylococcal cassette chromosome mec in methicillin-resistant Staphylococcus aureus by way of continuous culture. Antimicrob Agents Chemother 51(4): 1497-1499.
-
López M, Sáenz Y, Rojo-Bezares B, Martínez S, del Campo R, et al. (2009) Detection of vanA and vanB2- containing enterococci from food samples in Spain, including Enterococcus faecium strains of CC17 and the new singleton ST425. Int J Food Microbiol 133(1-2): 172-178.
-
Ma J, Zeng Z, Chen Z, Xu X, Wang X, et al. (2009)High prevalence of plasmid-mediated quinolone resistance determinants qnr, aac(6’)-Ib-cr, and qepA among ceftiofur-resistant Enterobacteriaceae isolates from companion and food-producing animals. Antimicrob Agents Chemother 53(2): 519-524.
-
Manageiro V, Jones-Dias D, Ferreira E, Louro D (2012) Antimicrobial Resistance Surveillance Program in Portugal, Genetic diversity and clonal evolution of carbapenem-resistant Acinetobacter baumannii isolates from Portugal and the dissemination of ST118. Int J Antimicrob Agents 40(5): 398-403.
-
MedVetNet (2006) MedVetNet Workpackage 9 Genetic Characterisation for Strains with Beta-lactamase Activity.
-
Mulvey MR, Soule G, Boyd D, Demczuk W, Ahmed R (2003) Characterization of the first extended-spectrum beta-lactamase-producing Salmonella isolate identified in Canada. J Clin Microbiol 41(1): 460-462.
-
Naiemi N, Duim B, Bart A (2006) A CTX-M extended- spectrum -lactamase in Pseudomonas aeruginosa and Stenotrophomonas maltophilia. J Med Microbiol 55(11): 1607-1608.
-
Olesen H, Hasman FM (2005) Aarestrup Prevalence of beta-lactamases among ampicillin-resistant Escherichia coli and Salmonella isolated from food animals in Denmark. Microb Drug Resist 10(4): 334-340.
-
Olsen RH, Schønheyder HC, Christensen H, Bisgaard M (2012) Enterococcus faecalis of human and poultry origin share virulence genes supporting the zoonotic potential of E. faecalis. Zoonoses and Public Health 59(4): 256-263.
-
Ruple-Czerniak A, Aceto HW, Bender JB, Paradis MR, Shaw SP, et al. (2014) Syndromic surveillance for evaluating the occurrence of healthcare-associated infections in equine hospitals. Equine Vet J 46(4): 435- 440.
-
Ruscher C, Lübke-Becker A, Semmler T, Wleklinski CG, Paasch A, et al. (2010) Widespread rapid emergence of a distinct methicillin- and multidrug-resistant Staphylococcus pseudintermedius (MRSP) genetic lineage in Europe. Vet Microbiol 144(3-4): 340-346.
-
Santos WG, Diniz RC, Carvalho IA, Coletto Freitas PM (2012) Infecção hospitalar em medicina veterinária. Revta Vet Zootec Minas 21: 10-15.
-
Stefani S, Chung DR, Lindsay JA, Friedrich AW, Kearns AM, et al. (2012) Meticillin-resistant Staphylococcus aureus (MRSA): global epidemiology and harmonisation of typing methods. Int J Antimicrob Agents 39(4): 273- 282.
-
Toleman MA, Simm AM, Murphy TA, Gales AC, Biedenbach DJ, et al. (2002) Molecular characterization of SPM-1, a novel metallo-β-lactamase isolated in Latin America: report from the SENTRY antimicrobial survillanceprogramme. J Antimicrob Chemother 50(5): 673-679.
-
Yılmaz ES, Aslantaş O, Önen SP, Türkyılmaz S, Kürekci C (2016) Prevalence, antimicrobial resistance and virulence traits in enterococci from food of animal origin in Turkey. LWT-Food Science and Technology 66: 20-26.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework