Morphological and Molecular Evidence of a Trematode Parasite Infected the Liver of Garfish Xenentodon cancila (Beloniformes: Belonidae) in India
Morphological traits to describe trematode parasites are sometimes difficult to identify and validate a species, especially in cases where many species are described from a single host. The current work uses molecular data to describe diplostomoid metacercariae supplemented with morphology found in freshwater garfish, Xenentodon cancila (Hamilton) collected from the River Ganga in district Bijnour, Meerut (Uttar Pradesh), India. The metacercariae were identified as Posthodiplostomum species, collected from the liver showed a high mass of cysts embedded in and surrounded by partially damaged liver tissue. Partial DNA sequences of the 18S, internal transcribed spacers (ITS1–5.8S–ITS2), and 28S of nuclear ribosomal DNA were generated and compared with available sequences of other congeners on the Genbank database. The phylogenetic analysis of 18S, the ITS cluster (ITS1–5.8S–ITS2), and 28S rDNA of Posthodiplostomum sp. from India fell within the superfamily Diplostomoidea, along with other members of Posthodiplostomum, which confirms its distinct status and places it close to other Indian species. In the Indian region, with morphology alone, many species are described as Neascus-type metacercariae that are awaiting their validation to be supplemented with molecular data. Furthermore, the validity of a few species of the genus Posthodiplostomum is also discussed in the present study.
Introduction
Transmission of infectious parasites by the increasing trade of fish poses a risk to human health [1]. One of the important trades of species introduction globally is aquaculture, which causes the transmission of infectious diseases like bacterial and parasitic diseases to wild and aquaculture-raised fish [2, 3, 4]. Piscivorous migratory birds as definitive hosts are also a major factor related to parasite transmission between the continents [5]. Posthodiplostomum Dubois, 1936, is a trematode genus rich in diversity and comprises parasites that are widespread worldwide [6, 7, 8, 9, 10, 11, 12]. Initially, for the identification of strigeid larva types, they were categorized into different types, i.e., Tetracotyle Filippi, 1854; Codonocephalus Diesing, 1850; Neascus Hughes, 1927; and Diplostomulum Brandes, 1892. Hughes (1927) proposed a group of larval trematodes, i.e., Neascus, to contain metacercariae, and Dubois (1936) described the genus Posthodiplostomum, which is in the family Diplostomidae. Parasites of this genus have a three-host life cycle, with snails as first intermediate hosts, fish acting as second intermediate hosts, and the fish-eating birds as definitive hosts [11, 13]. According to research, during the initial phase of infection, metacercaria penetration and migration cause mechanical damage, hemorrhage, and other secondary bacterial infections in fishes [15]. Infection caused by these trematodes is called black-spot disease and may have its origins in fish muscles due to the deposition of melanocytes around metacercariae [14]. A few studies have mentioned Posthodiplostomum cuticola (Nordmann, 1832) as the causative agent of black-spot disease, and it is one of the common diseases that affect many fish species [15, 16, 17, 18, 19, 20]. Fish infected with black-spot diseases have had reduced growth along with other deformities and mortalities [12, 19, 21, 22]. Sometimes, the presence of metacercaria in the liver of infected fish can be associated with changes in hepatic tissues like parenchymal atrophy, necrosis, and fibrosis that lead to dysfunction in digestion and malnutrition [23]. Additionally, Posthodiplostomum species infection in fishes may cause weight loss, slow developmental processes, and mortality, making them less able to avoid predation by definitive hosts, i.e., fish-eating birds [15, 24, 25, 26].
Many species of Posthodiplostomum have been described worldwide, while more than twenty-five species are reported from India, described as Neascus-type metacercariae and as Posthodiplostomum species [27]. Their life cycle is complex, and it is very difficult to study or link different stages in the laboratory. In India, most of the Posthodiplostomum species are characterised based on morphology alone, which cannot be considered reliable due to a lack of descriptions or deposited specimens. Difficulties in identification can be overcome by the generation of molecular data to link different developmental stages, as suggested in various studies [28, 29, 30]. Molecular identification would eventually improve our understanding of the correct diversity of Posthodiplostomum species that are scarce and limited, especially in India and help in linking the adult and metacercariae stages.
As part of an ongoing survey of diplostomid metacercaria (Neascus type) from freshwater fishes in India, Posthodiplostomum species was collected in the liver of the garfish Xenentodon cancila with severe liver infection. A combined morphological, histological, and molecular analysis using the 18S, ITS cluster (ITS1–5.8S–ITS2), and 28S rDNA gene sequences was provided to evaluate the phylogenetic position of our specimens in relation to other closely related species.
Materials and Methods
Parasite Collection and Morphology
During a helminthological survey in Bijnor (29° 23′ N, 79° 11′ E), Uttar Pradesh, India, between November 2020 and May 2021, a total of 57 garfish, Xenentodon cancila (Hamilton, 1822) (Beloniformes: Belonidae), were collected. After that, fish internal organs were examined with the aid of a stereomicroscope (Motic SMZ-168 series, Xiamen, People’s Republic of China) to detect the infection of parasites. Metacercariae were found to infect the liver, removed from the cysts with the help of a fine needle, and placed in a 0.9% saline solution for approximately 10–15 minutes. The morphology of metacercariae was investigated using both live and preserved specimens. For a live study of morphology, metacercariae were placed on a slide using a pipette in physiological saline (0.9%), coverslipped, and photographed by a Nikon Eclipse Ts2 microscope. Some of them were fixed in 10% formalin for permanent slide preparation, and some were immediately fixed in 95% ethanol for molecular analysis. For permanent slide preparation, they were stained with aceto-alum carmine, dehydrated in ascending grades of ethanol, cleared in xylene, and mounted in Canada balsam. Measurements were taken with the help of the digital images and NIS Elements 5.10 image analysis software. All measurements are given in millimeters. Abbreviations of the measurements are provided in the Table 2. Voucher specimens were deposited at the museum collection of the Department of Zoology, Chaudhary Charan Singh University, Meerut (U.P.), India, and at the Museum d’Histoire Naturelle, Geneva, Switzerland.
The infected fish liver sections were preserved immediately in 4% formalin and processed for histopathology. The samples were washed in 80% ethanol numerous times, implanted in paraffin wax, cut into 5- to 8-m-thick sections, and stained with hematoxylin and eosin. The histological sections were photographed by a Nikon Eclipse T_s_2 microscope equipped with NIS Elements 5.10 image analysis software.
DNA Isolation and Amplification
DNA extraction from ethanol-fixed metacercariae (n= 02) was performed using the QIAGEN DNeasyTM tissue kit (Qiagen, Hilden, Germany) as per manufacturer’s instructions. Sequences of the 18S, ITS cluster (ITS1–5.8S– ITS2), and 28S rDNA were obtained by PCR amplification according the primers suggested by Shinad et al. [31]. For all primers, PCR conditions were also performed according to Shinad et al. [31]. PCR products were then purified using Purelink™ Quick Gel Extraction and PCR Purification Combo Kit (Invitrogen, Löhne, Germany) according to the manufacturer’s instructions. Sequencing reactions were performed in both directions using the ABI Big Dye Terminator Cycle Sequencing Reaction Kit (Applied Biosystems, Foster City, California, USA) from the above mentioned primers and using the primers BD1 and BD2 [32] for the ITS cluster (ITS1-5.8S-ITS2) to acquire a long stretch of sequences.
Phylogenetic Analysis
The generated sequences were visualized, assembled, and edited using BioEdit software [33]. Sequences of other diplostomids available at GenBank were retrieved after a BLASTn search. The sequences used in the phylogenetic analyses are presented in Table 1. The obtained sequences were aligned separately using ClustalW, which was implemented in MEGA vr. 11 [48]. Phylogenetic analyses were conducted using maximum likelihood (ML) and Bayesian inference (BI) algorithms. The best nucleotide substitution models for 18S, ITS cluster (ITS1-5.8S-ITS2), and 28S rDNA data sets were invariant sites and gamma-distributed among site variation (GTR + I + G). Pairwise comparisons for each region were estimated using MEGA v. 11. The ML trees were computed in MEGA vr. 11 and nodal support values were estimated from 1000 bootstrap pseudo-replicates. The BI analyses were carried out in TOPALi [49] using Markov chain Monte Carlo (MCMC) searches with two concurrent runs of four chains for 1,000,000 generations, with every 100th tree saved. The first 25% of the sampled trees were discarded as “burn-in.” Tylodelphys species was selected as an out group for the 18S, ITS1–5.8S–ITS2 region, and 28S rDNA gene analyses.
Results
During the study, a total of 57 garfish specimens were collected, of which 54 were found infected with metacercariae. Metacercariae were found in the liver tissue of the infected fish in the form of numerous small cysts; the cyst wall was thin and delicate (Figure 1). Each fish was parasitized by 50–190 metacercariae. The excysted specimens of the present study clearly showed the morphological features of the genus Posthodiplosomum Dubois, 1936 and of species Posthodiplostomum pandei (=Neascus pandei) Rai and Pande, 1964 [50].
Morphological Description of Posthodiplostomum pandei (=Neascus pandei) Rai and Pande, 1964 (Figures 1-10)
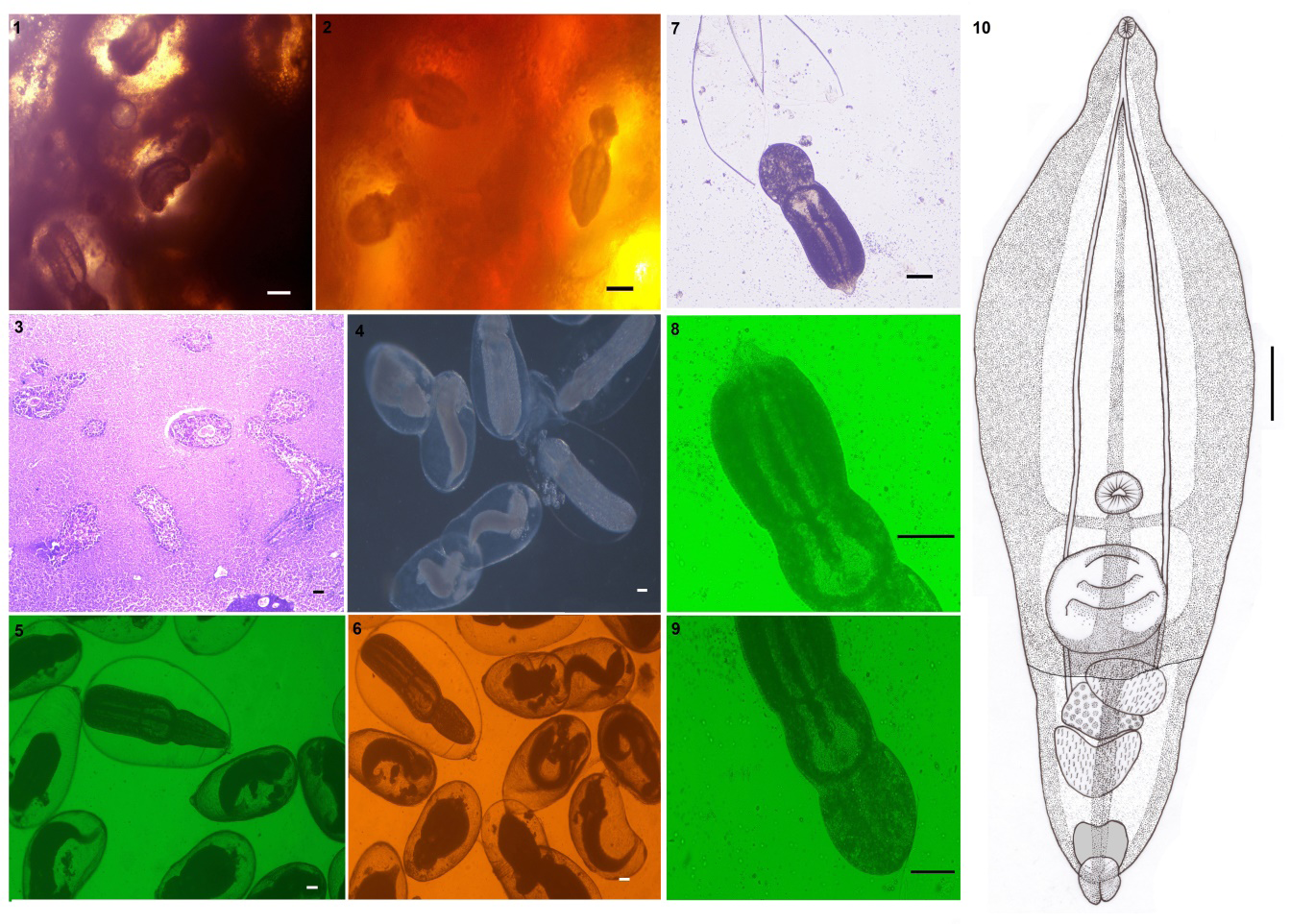
Figures 1-10: Posthodiplostomum pandei (1,2) Encysted metacercariae embedded in liver tissue; (3) Histology of the liver shows encysted metacercaria within the liver tissue layer (arrow head); (4-6) Encysted metacercaria collected from X. cancila’s liver tissue; (7) Metacercaria exit from the cyst; (8, 9) Excysted metacercaria (10) Line drawing. Scale bars (1, 2) 100 µm; (3-6) 50 µm; (7-10) 100 µm.
Metacercariae (n= 14). Encysted metacercariae with a thin, single layer of cystic wall. Spatulate, divided into two parts, or bipartite body of excysted metacercaria. Leaf-shaped forebody, larger than the hindbody. Hindbody are typically bulb-shaped. Pseudosuckers absent. At the anterior end, sub- terminal, there is a spherical-shaped oral sucker. The ventral sucker is spherical, larger than the oral sucker, and situated in the body’s center, anterior to the holdfast organ. The holdfast organ is located near the forebody’s posterior margin. The distinction between genital primordials is poor. The testes are divided into two parts: the anterior and posterior testes. Ovary intra-testicular. The excretory bladder is oval and terminates with a genital pore.
All the measurements of Posthodiplostomum species collected in the present study were compared in their morphology with those of other congeneric species in Table 2. Based on the morphology, the present species is recognised and differentiated as Neascus pandei Rai and Pande, 1964, and molecular biology confirms it as a Posthodiplostomum species. In histological sections, the cysts of P. pandei (=N. pandei) Rai and Pande, 1964 were found inside the liver and had a heavy infection (Figure 3).
- Host: Garfish, Xenentodon cancila Hamilton, 1822 (Beloniformes: Belonidae).
- Locality: Bijnor (29° 23′ N, 79° 11′ E), Uttar Pradesh, India.
- Site of infection: Liver (cysts embedded in the liver tissue).
- Prevalence in intermediate hosts: 95% (54 infected out of 57 hosts).
- Specimens deposited: Voucher specimens were submitted to the collection of the Museum, Department of Zoology, Chaudhary Charan Singh University, Meerut (U.P.), India (HS-TR/2022/01) and to the Museum d’Histoire Naturelle, Geneva, Switzerland (MHNG- PLAT-0144332).
- Representative DNA sequence: The newly generated sequences were deposited in GenBank under the following accession numbers: 18S rDNA gene: OP324624 (1068 bp), OP324628 (1079 bp); ITS1-5.8S-ITS2 region: OP324627 (1190 bp), OP324629 (1202 bp); 28S rDNA gene: OP324625 (1019 bp), OP324626 (1001 bp).
Remarks
Based on the shape of body parts and morphometric measurements, we identified our specimens as Posthodiplostomum pandei (=Neascus pandei) Rai and Pande, 1964. Many descriptions of Posthodiplostomum or Neascus-like metacercaria from X. cancila liver show inadequate data that results in taxonomic confusion in the identification of species, which leads to the addition of new species. Many previous studies from India were based solely on morphology and described as Neascus-type metacercaria species. In India, about twenty-six species of Neascus– type metacercariae and five of Posthodiplostomum were described (Table 3). Eleven of these have been reported solely from X. cancila as mentioned in Table 2, although they show minor variations in their morphometry. N. mesentriformis, N. baughi, N. nanaksagrensis, N. simhai, N. moghei, N. vedi, and N. khurramnagarensis have larger body size and width compared to our specimens, which clearly differentiate them from P. pandei (Table 2). There are no substantial morphometrical differences found in the body parts and morphology of N. xenentodoni, N. hepatica, and N. srivastavi with P. pandei, showing they are most closely related in nearly all metrics, which predicts they might be similar/closest species to P. pandei (Table 2). Although N. xenentodoni is close in morphology to P. pandei, as the molecular data of N. xenentodoni becomes available, it will be better to discuss it further. As we are not able to find out the deposited type specimens of N. hepatica and N. srivastavi for further clarification, it is too early to suggest them as synonyms with P. pandei. Thus, we consider P. pandei to be a valid species, and anyhow, these species (N. hepatica and N. srivastavi) should be taken into account in future studies in order to uncover their validity or synonymies with P. pandei.
Phylogenetic Analysis
The amplification and sequencing of the 18S gene of rDNA were successfully performed from two isolates of Posthodiplostomum pandei (=N. pandei) in the present study. The generated sequences were aligned with other diplostomids available in GenBank, as shown in the table (Table 1). The 18S sequences generated in this study show no intraspecific variations (Figure 11). The 18S sequence of P. pandei differed from other Posthodiplostomum species from different geographical regions by 1.5–3.9%. The phylogenetic tree obtained by ML and BI analyses of 18S sequences showed similar topologies. For comparison of the 18S gene with Indian species, only one sequence is available to date from C. punctata (KF738455), which is far away from our specimens. The analysis of 18s gene shows our specimens formed a well-supported clade found close to the Mexican isolate of P. minimum (PMU88074) and showing interspecific divergence of 1.5% (Figure 11).
| Species | Host | Location | GenBank accession no. | References | ||
|---|---|---|---|---|---|---|
| 18S | ITS region | 28S | ||||
| Ornithodiplostomum scardinii | Scardinius erythrophthalmus | Czech Republic | KX931443 | - | KX931427 | Stoyanov, et al. [11] |
| O. scardinii | Ampullaceana balthica | Denmark | - | MW001049, MW001051 | - | Duan, et al. [8] |
| Ornithodiplostomum sp. | HNA | Canada | - | KY951727 | - | Blasco-costa, et al. [29] |
| Mesoophorodiplostomum pricei | Morone americana | Canada | HM064959 | HM064960 | - | Locke, et al. [34] |
| Posthodiplostomum pricei | L. delawarensis | USA | - | - | MZ710972 | Achatz, et al. [35] |
| Posthodiplostomum centrarchi | L. gibbosus | Slovakia | KX931442 | - | - | Stoyanov, et al. [11] |
| P. centrarchi | L. gibbosus | Bulgaria | KX931441 | - | - | Stoyanov, et al. [11] |
| Posthodiplostomum centrarchi | L. gibbosus | USA | - | - | OM638425 | Koxlien, et al. 2022 |
| P. centrarchi | L. gibbosus | Hungary | - | MN080277, MN080281 | - | Cech, et al. [7] |
| P. centrarchi | Ardea herodias | Canada | - | MH521251 | - | Locke, et al. [36] |
| Posthodiplostomum sp. | L. gibbosus | Canada | HM064958 | HM064955 | - | Locke, et al. [34] |
| Posthodiplostomum sp. | Micropterus salmoides | Canada | HM064962 | - | - | Locke, et al. [34] |
| Posthodiplostomum sp. | L. gibbosus | Slovakia | - | KX931442 | - | Stoyanov, et al. [11] |
| Posthodiplostomum brevicaudatum | Gasterosteus aculeatus | Bulgaria | KX931431, KX931432 | - | - | Stoyanov, et al. [11] |
| P. brevicaudatum | Perca fluviatilis | Czech Republic | - | - | KX931426 | Stoyanov et al. 2017 [11] |
| Posthodiplostomum minimum | HNA | Mexico | U88074 | - | - | Campos, et al. [37] |
| Posthodiplostomum pandei† | Xenentodon cancila | India | OP324624, OP324628 | OP324627, OP324629 | OP324625, OP324626 | Present study |
| Posthodiplostomum sp. | L. gibbosus | Canada | FJ469590 | - | - | Moszczynska, et al. [38] |
| Posthodiplostomum cuticola | Ardea cinerea | Czech Republic | MK089352 | - | - | Heneberg, et al. [39] |
| P. cuticola | Nycticorax nycticorax | Ukraine | - | - | MZ710955 | Achatz, et al. [35] |
| P. cuticola | Anisus vortex | Denmark | - | MW001124, MW001121 | - | Duan, et al. [8] |
| P. cuticola | Abramis brama | Hungary | - | MN080266, MN080287 | - | Cech, et al. [7] |
| Posthodiplostomum sp. | L. gibbosus | Canada | - | HM064949 | - | Locke, et al. [34] |
| Posthodiplostomum sp. | Tilapia sparrmanii | South Africa | - | MK604881 | - | Hoogendoorn, et al. [19] |
| Posthodiplostomum sp. | Channa argus | Japan | AB693170 | AB693170 | - | Nguyen et al. 2012 [40] |
| Posthodiplostomum sp. | Channa punctata | India | KF738455 | KF738447 | KF738450 | Athokpam, et al. [41] |
| P. minimum | Lepomis macrochirus | USA | KY809062 | - | - | Lovy, et al. [42] |
| P. minimum | N. nycticorax | USA | - | - | MZ710962 | Achatz, et al. [35] |
| P. centrarchi | A. herodias | Canada | MH521251 | - | - | Locke, et al. [36] |
| P. minimum | HNA | USA | AY245767 | - | - | Flowers, et al. 2003 |
| Posthodiplostomum sp. | Nannopterum brasilianus | Mexico | MF398354 | - | MF398331 | Hernandez- mena, et al. [43] |
| Posthodiplostomum macrocotyle | Busarellus nigricollis | Brazil | - | - | MZ710958 | Achatz, et al. [35] |
| Posthodiplostomum microsicya | Tigrisoma lineatum | Brazil | - | - | MZ710960 | Achatz, et al. [35] |
| Posthodiplostomum erickgreenei | Pandion haliaetus | USA | - | - | MZ710956 | Achatz, et al. [35] |
| Posthodiplostomum eurypygae | Eurypyga helias | Brazil | - | - | MZ710957 | Achatz et al. 2021 [35] |
| Posthodiplostomum nanum | Ardea alba | USA | - | - | MZ710963 | Achatz, et al. [35] |
| P. nanum | Gundlachia ticaga | Brazil | - | MH358392 | - | Lopez- Hernandez, et al. [23] |
| P. nanum | Poecilia reticulata | Brazil | - | MH358393 | - | Lopez- Hernandez, 2018 [23] |
| Posthodiplostomum sp. | A. herodias | USA | - | - | MZ710994 | Achatz, et al. [35] |
| Posthodiplostomum xenentodoni (=Neascus xenentodoni) | X. cancila | India | - | KY234201 | KY234203 | Choudhary, et al. [44] |
| Posthodiplostomum sp. | Channa striata | Vietnam | - | - | MT394045 | Sokolov and Gordeev, 2020 [45] |
| Posthodiplostomum sp. | Trichopodus trichopterus | Vietnam | - | - | MT394051 | Sokolov, et al. [45] |
| Posthodiplostomum hanumanthai (=Neascus hanumanthai) | C. punctata | India | - | KY234199 | KY042122 | Choudhary, et al. [44] |
| Posthodiplostomum gussevi (=Neascus gussevi) | Colisa fasciata | India | - | KY234200 | KY234202 | Choudhary, et al. [44] |
| Posthodiplostomum pacificus | Larus californicus | USA | - | - | MZ710967 | Achatz, et al. [35] |
| Posthodiplostomum cf. anterovarium | L. gibbosus | USA | - | - | OM688205 | Koxlien, et al. 2022 |
| P. cf. anterovarium | L. gibbosus | USA | - | - | MZ710942 | Achatz, et al. [35] |
| Posthodiplostomum cf. podicipitis | Lophodytes cucullatus | USA | - | - | MZ710969 | Achatz, et al. [35] |
| Posthodiplostomum sp. | Physa sp. | USA | - | - | MZ710982 | Achatz, et al. [35] |
| Posthodiplostomum recurvirostrae | Recurvirostra americana | USA | - | - | MZ710975 | Achatz, et al. [35] |
| Posthodiplostomum ptychocheilus | Mergus merganser | USA | - | - | MZ710974 | Achatz, et al. [35] |
| Posthodiplostomum sp. | Trichopodus pectoralis | Thailand | ON614094 | - | - | Nguyen, 2022 |
| Posthodiplostomum sp. | Skiffia lermae | Mexico | - | OK315760, OK315761 | - | Perez-Ponce de Leon, et al. [46] |
| Posthodiplostomum sp. | Allotoca dugesii | Mexico | - | OK315771 | - | Perez-Ponce de Leon, et al. [46] |
| Posthodiplostomum sp. | Gambusia sp. | Mexico | - | OK315772 | - | Perez-Ponce de Leon, et al. [46] |
| Posthodiplostomum sp. | Pimephales promelas | Mexico | - | OK315775 | - | Perez-Ponce de Leon, et al. [46] |
| Outgroups | ||||||
| Tylodelphys immer | Gavia immer | Canada | MH521252 | - | - | Locke, et al. [36] |
| Tylodelphys aztecae | Podilymbus podiceps | Mexico | - | - | MF398337 | Hernandez- mena, et al. [43] |
| Tylodelphys sp. | Aechmophorus occidentalis | Mexico | - | MK177831 | - | Sereno-Uribe, et al. [47] |
Table 1: Trematode species are included in the phylogenetic analysis with information on the host, locality and GenBank accession
| Body parts | P. pandei (=N. pandei) | P. pandei (=N. pandei) | N. mesentri formis | N. hepatica | P. xenen todoni (=N. xenen todoni) | N. baughi | N. nanaks agrensis | N. simhai | N. moghei | N. vedi | N. sriva stavi | N. khur ram naga rensis |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refe rence | Present study | Rai and Pande, 1964 [50] | Rai and Pande, 1964 [50] | Chakrabarti, 1970 [51] | Pandey, 1971 [52] | Baugh and Chakrabarti, 1977 [53] | Baugh and Chakra barti, 1977 [53] | Agrawal and Khan, 1982 [54] | Agrawal and Khan, 1982 [24] | Pandey and Tiwari, 1986 [55] | Pandey and Pandey, 2000 [56] | Gupta, et al. [57] |
| Infected Organ | Cysts in Liver | Liver | Mesentry | Cysts in Liver | Liver and cranium | Body cavity | Cysts in liver and gonads | Body muscles | Liver | Body muscles | Liver | Liver |
| CSTL | 0.61 (0.49-0.76) | 0.51-0.7 | 1.4-1.8 | 0.72-0.87 | 0.52-0.67 | 1.32-1.50 | 1.05-1.03 | 1.17- 1.19 | 0.99- 1.00 | 1.13- 1.33 | 0.58- 0.69 | 1.13 - 1.33 |
| CSTW | 0.37 (0.28-0.46) | 0.31-0.6 | 0.8-0.9 | 0.44-0.52 | - | 0.65-0.76 | 0.50-0.68 | 0.76- 0.78 | 0.62- 0.64 | 0.90- 1.20 | 0.41- 0.43- | 0.90 - 1.20 |
| WBL | 0.89 (0.51-1.2) | 0.78-0.85 | 0.64-0.72 | - | - | - | - | - | - | - | - | - |
| WBW | 0.30 (0.11z-0.42) | 0.25-0.34 | - | - | - | - | - | - | - | - | - | - |
| FBL | 0.62 (0.38-0.86) | - | - | 0.46-0.75 | 0.67-1.05 | 1.28-1.65 | 0.78-1.05 | 1.07- 1.20 | 0.97- 0.98 | 0.47- 1.03 | 0.54- 0.64 | 0.47 - 1.03 |
| FBW | 0.30 (0.11-0.42) | - | - | 0.23-0.39 | 0.24-0.33 | 0.90-1.08 | 0.61-0.76 | 0.54- 0.60 | 0.37- 0.38 | 0.19- 0.21 | 0.28- 0.44 | 0.19 - 0.21 |
| HBL | 0.23 (0.12-0.33) | - | - | 0.18-0.26 | 0.18-0.30 | 0.62-0.82 | 0.41-0.65 | 0.37- 0.39 | 0.26- 0.27 | 0.19- 0.22 | 0.20- 0.25 | 0.19 - 0.22 |
| HBW | 0.16 (0.07-0.20) | - | - | 0.15-0.24 | 0.14-0.27 | 0.56-0.71 | 0.53-0.70 | 0.27- 0.22 | 0.28- 0.29 | 0.13- 0.19 | 0.18- 0.27 | 0.13 - 0.19 |
| OSL | 0.02 (0.01-0.03) | 0.024-0.04 | 0.04-0.05 | 0.04-0.07 | - | 0.05-0.08 | 0.04-0.06 | 0.05- 0.06 | 0.07- 0.75 | 0.03- 0.04 | 0.02- 0.04 | 0.03 - 0.04 |
| OSW | 0.02 (0.01-0.02) | 0.024-0.04 | 0.04-0.05 | 0.03-0.05 | - | 0.03-0.05 | - | 0.03- 0.04 | - | 0.02- 0.03 | 0.02- 0.03 | 0.02 - 0.03 |
| VSL | 0.04 (0.01-0.05) | 0.04-0.06 | 0.04-0.05 | 0.05-0.09 | 0.04-0.07 | 0.06-0.09 | 0.06-0.09 | 0.09- 0.10 | - | 0.03- 0.04 | 0.04- 0.06 | 0.03 - 0.04 |
| VSW | 0.04 (0.02-0.05) | 0.04-0.05 | 0.05-0.09 | - | 0.06-0.09 | - | 0.07- 0.08 | - | 0.02- 0.03 | 0.05- 0.08 | 0.02 -0.03 | |
| HOL | 0.12 (0.06-0.18) | 0.12-0.14 | 0.14-0.16 | - | - | - | - | 0.14- 0.16 | - | - | 0.12 - 0.17 | |
| HOW | 0.14(0.06- 0.19) | 0.12-0.14 | - | - | - | - | - | - | - | - | 0.10 - 0.16 | |
| ATL | 0.05 (0.02-0.07) | 0.06-0.07 | - | 0.10-0.15 | - | 0.20-0.26 | 0.17-0.20 | 0.10- 0.12 | 0.13- 0.14 | 0.02- 0.07 | 0.03- 0.05 | 0.02- 0.07 |
| ATW | 0.07 (0.03-0.10) | 0.02-0.03 | - | 0.06-0.10 | - | 0.25-0.32 | 0.16-0.23 | 0.13- 0.14 | 0.04- 0.045 | 0.10- 0.16 | 0.06- 0.09 | 0.06 - 0.12 |
| PTL | 0.06 (0.03-0.07) | 0.08-0.1 | - | 0.16-0.22 | - | 0.41-0.54 | 0.35-0.40 | 0.23- 0.25 | 0.13- 0.14 | 0.01- 0.03 | 0.03- 0.05 | 0.01 - 0.03 |
| PTW | 0.11 (0.04-0.15) | 0.03-0.05 | - | 0.02-0.07 | - | 0.07-0.13 | 0.07-0.10 | 0.10- 0.13 | 0.04- 0.045 | 0.05 -0.09 | 0.06- 0.09 | 0.05 - 0.09 |
| OL | 0.06 (0.02-0.09) | - | - | 0.08-0.12 | - | - | 0.05-0.07 | 0.13- 0.14 | 0.11- 0.12 | 0.01- 0.03 | 0.04- 0.05 | - |
| OW | 0.08 (0.01-0.11) | - | - | 0.0-0.08 | - | - | 0.06-0.09 | 0.07- 0.08 | 0.01- 0.015 | 0.05- 0.09 | 0.05- 0.06 | - |
| EBL | 0.07 (0.03-0.09) | - | - | - | - | - | - | - | - | - | - | - |
| EBW | 0.05 (0.02-0.07) | - | - | - | - | - | - | - | - | - | - | - |
| OSVS | 0.42 (0.24-0.63) | - | - | - | - | - | - | - | - | - | - | - |
| VSHO | 0.04 (0.02-0.07) | - | - | - | - | - | - | - | - | - | - | - |
Table 2: Comparative measurements shown as mean (range) of _Neascus_-type and _Posthodiplostomum_ species, all infected the sa
CSTL= Cyst length, CSTW= Cyst width, WBL= Whole body length, WBW= Whole body width, FBL= Fore body length, FBW= Fore body width, HBL= Hind body length, HBW= Hind body width, OSL= Oral sucker length, OSW= Oral sucker width, VSL= Ventral sucker length, VSW= Ventral sucker width, HOL= Holdfast organ length, HOW= Holdfast organ width, ATL= Anterior testis length, ATW= Anterior testis width, PTL= Posterior testis length, PTW= Posterior testis width, OL= Ovary length, OW= Ovary width, EBL= Excretory bladder length, EBW= Excretory bladder width, OSVS= Distance between oral sucker and ventral sucker, VSHO= Distance between ventral sucker and holdfast organ. Table 2: Comparative measurements shown as mean (range) of Neascus-type and Posthodiplostomum species, all infected the same host (Xenentodon cancila) from India.
| Species | Host | Site of infection | Species status/ developmental stage | Identification evidence | References |
|---|---|---|---|---|---|
| P. botauri | NA | Intestine | Valid/A | Morphology | Vidyarthi [58] |
| N. vetastai | Schizothorax esocinus, S. micropogon, S. niger | Cyst on skin and viscera | Incertae sedis/M | Morphology | Kaw [59] |
| N. chelai | Chela clupeoides | Encysted in the integument and muscles | Incertae sedis/M | Morphology | Khera [60] |
| P. pandei | Xenentodon cancila | Liver | Valid/M | Morphology/ Molecular* | Rai, et al. [50] |
| N. mesentriformis | X. cancila | Mesentery | Incertae sedis/M | Morphology | Rai, et al. [50] |
| P. milvi | NA | Intestine | Valid/A | Morphology | Fotedar, et al. [61] |
| N. indicus | Nuria dursica, Catla catla | Cyst in muscles below scales | Incertae sedis/M | Morphology | Thapar [62] |
| N. cirrhinus | Cirrhinus mrigala | Cyst in muscles | Incertae sedis/M | Morphology | Thapar [62] |
| N. muscularis | Channa punctata | Muscles | Incertae sedis/M | Morphology | Rai , et al. [50] |
| N. elongatus | Colisa fasciata | Mesenteries inside body cavity | Incertae sedis/M | Morphology | Singh [63]; Pandey 64] |
| N. hepatica | X. cancila | Cyst in liver | Incertae sedis/M | Morphology | Chakrabarti [51]; Vankara, et al. [65] |
| P. mehtai | Milvus migrans | Intestine | Valid/A | Morphology | Gupta, et al. 1974 [66] |
| N. channi | C. punctata | Cranium | Incertae sedis/M | Morphology | Pandey [52] |
| P. xenentodoni | X. cancila | Liver and cranium | Valid/M | Morphology/ Molecular | Pandey [52]; Choudhary, et al. [44] |
| N. komiyai | Glossgobius giuris | Cyst attached to stomach | Incertae sedis/M | Morphology | Pandey [67] |
| N. hoffmani | Nandus nandus | Stomach | Incertae sedis/M | Morphology | Pandey [67] |
| N. gussevi | C. punctata | Cyst attached to visceral organs | Valid/M | Morphology/ Molecular | Chakrabarti [68]; Choudhary, et al. [44] |
| N. baughi | X. cancila | Body cavity | Incertae sedis/M | Morphology | Baugh, et al [53] |
| N. nanaksagrensis | X. cancila | Cysts attached to liver, gonads and in liver | Incertae sedis/M | Morphology | Baugh, et al. [53] |
| N. chauhani | Heteropneustus fossilis | Free in body muscles | Incertae sedis/M | Morphology | Agrawal and Khan, 1982 [54] |
| N. hanumanthai | C. punctata | Cyst in muscles | Valid/M | Morphology/ Molecular | Agrawal, et al. [54]; Choudhary, et al. [44] |
| N. simhai | X. cancila | Cyst in body muscles | Incertae sedis/M | Morphology | Agrawal, et al. [54] |
| N. moghei | X. cancila | Liver | Incertae sedis/M | Morphology | Agrawal, et al. [54] |
| N. shahjahanpurensis | Clarius batrachus | Muscles | Incertae sedis/M | Morphology | Pandey, et al. [55] |
| N. ramalingami | Labeo rohita | Body muscles | Incertae sedis/M | Morphology | Pandey and Tiwari, 1986 [55] |
| N. vedi | X. cancila | Body muscles | Incertae sedis/M | Morphology | Pandey, et al. [55] |
| N. punctatusi | C. punctata | Cranial cavity | Incertae sedis/M | Morphology | Dhanukumari [69] |
| P. grayii | Apocheilus panchax | Liver | Valid/A | Morphology | Verma, [70]; Madhavi, et al. [71] |
| N. srivastavi | X. cancila | Liver | Incertae sedis/M | Morphology | Pandey, et al. [56] |
| Posthodiplostomum sp. | C. punctata | Body muscles | Valid/M | Morphology/ Molecular | Athokpam, et al. [41] |
| N. khurramnagarensis | X. cancila | Liver | Incertae sedis/M | Morphology | Gupta, et al. [57] |
Table 3: Species of Neascus-type or Posthodiplostomum described from India on the basis of metacercariae specimens, infected host
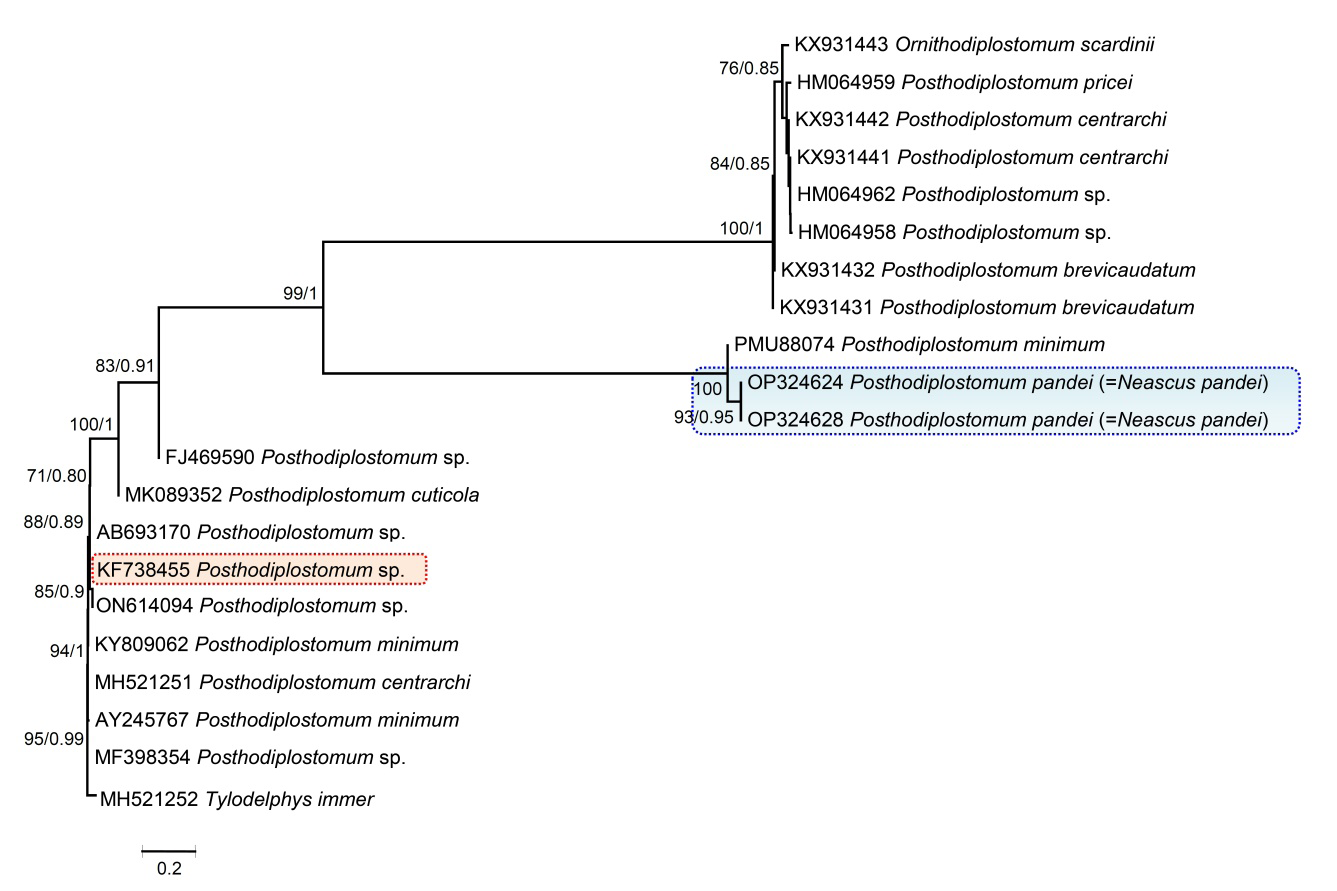
Figure 11: Phylogenetic interrelationships among Posthodiplostomum species based on Maximum Likelihood (ML) and Bayesian Inference (BI) analyses of partial 18S rDNA gene sequences. The scale-bar indicates the number of substitutions per site. Species names are provided after GenBank accession numbers. The numbers represent bootstrap values greater than 70%. The present studied species, Posthodiplostomum pandei, is indicated by the blue line box, while the red line box shows the other Indian species.
In this study, the ITS (ITS1–5.8S–ITS2) sequences of two individuals of P. pandei were generated and aligned with other sequences of the diplostomids (Figure 12). The genetic divergence between the different species of Posthodiplostomum ranged from 0.92 to 4.1%. Moreover, ITS (ITS1–5.8S–ITS2) sequences were available for the following Indian species: N. hanumanthai (KY042122), N. gussevi (KY234202), N. xenentodoni (KY234203), and Posthodiplostomum sp. (KF738447), whose genetic divergence ranged from 0.92 to 2.6%. Maximum likelihood (ML) and Bayesian inference (BI) analyses produced a similar tree in topology that shows N. xenentodoni to be the closest to our specimens; both belong to the same host (Figure 12).
The 28S sequences in the current phylogenetic analysis clearly demonstrated the status of P. pandei within Posthodiplostomum (Figure 13). The Indian species N. hanumanthai (KY042122), N. gussevi (KY234202), N. xenentodoni (KY234203), and an unknown Posthodiplostomum sp. (KF738450) were positioned with Posthodiplostomum members, validated their status, and formed a well-supported clade. Choudhary, et al. [44] were not mentioned the status of N. xenentodoni regarding this is a species of Posthodiplostomum as the molecular data predicts_, the present study analysis confirmed that it is a species of _Posthodiplostomum with good bootstrap values (90% ML and 0.92 BI) and sister to the present specimens (Figure 13). In comparison, the genetic divergence among Posthodiplostomum and P. pandei ranged from 1.2 to 4.5%. P. pandei and other Indian Posthodiplostomum species had 1.2– 2.2% genetic variation. Analysis shows all the 28S sequences of Indian species and the data generated in this study are nested within a clade with strong bootstrap support and Bayesian posterior probability values of 83 and 0.9 (Figure 13).
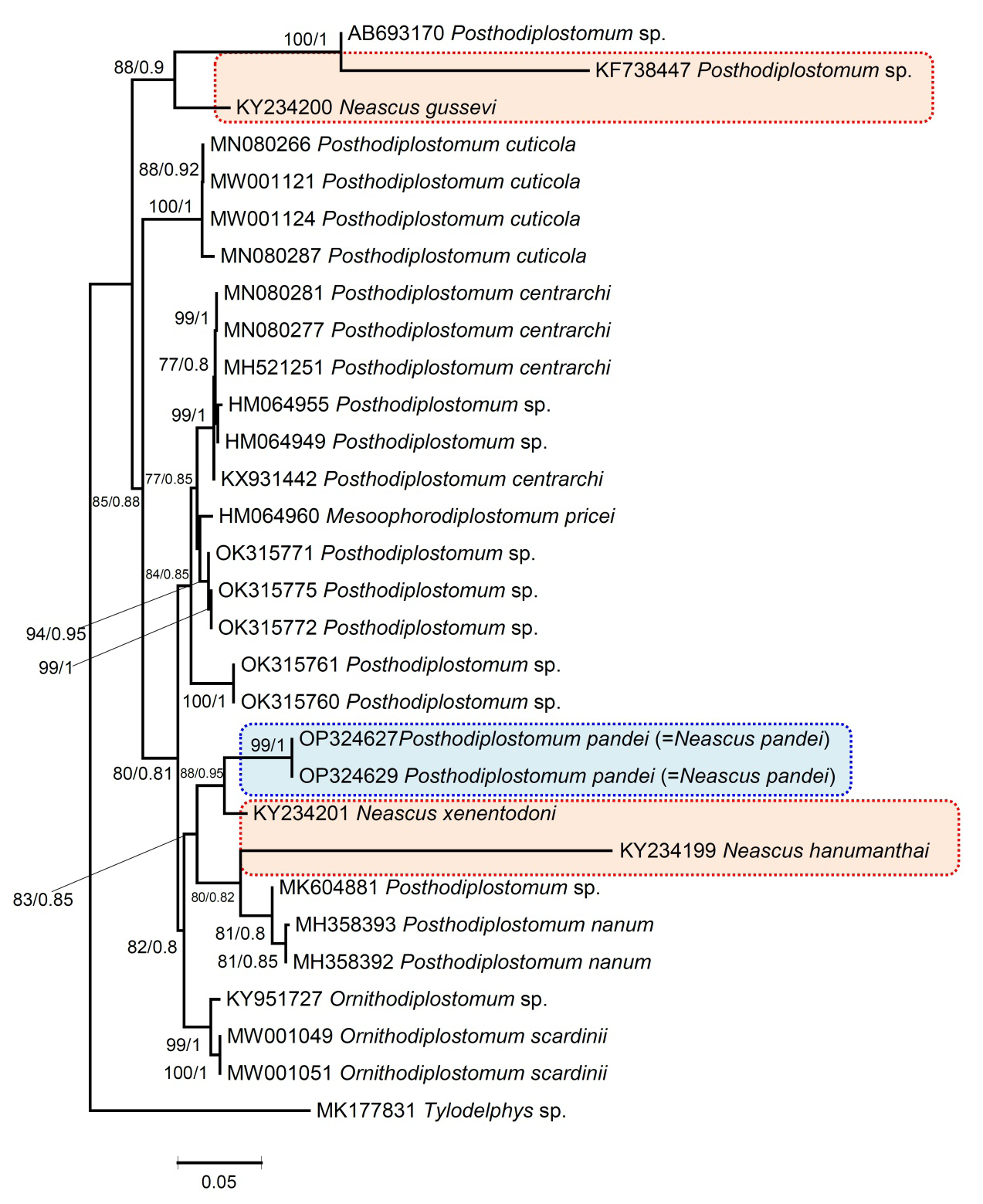
Figure 12: Phylogenetic reconstruction using ITS region (ITS1-5.8S-ITS2) sequences of Posthodiplostomum species (shown in blue box). The numbers represent bootstrap values greater than 70%. Numbers above branches indicate nodal support as maximum likelihood (ML) and posterior probabilities from BI. The scale bar indicates the expected number of substitutions per site. Species names are provided after Gen Bank accession numbers. The blue line box represents the currently studied species P. pandei, while the red line box represents the other Indian species.
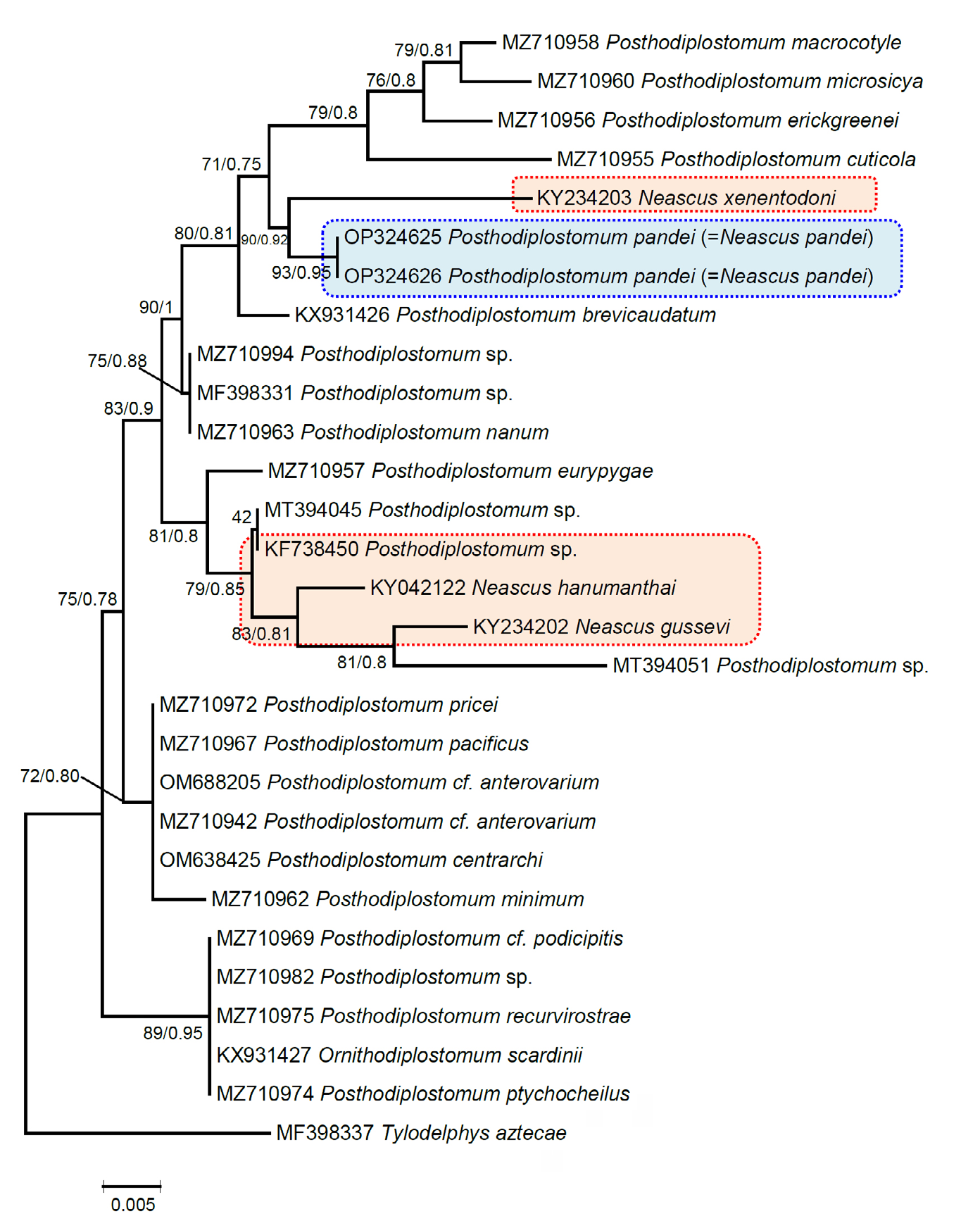
Figure 13: Phylogenetic relationship between Posthodiplostomum species inferred from sequences of the 28S rDNA gene based on maximum likelihood (ML) and Bayesian inference (BI) analyses. Nodal support is indicated as ML/BI. The numbers represent bootstrap values greater than 70%. The scale bar indicates the expected number of substitutions per site. Species names are provided after GenBank accession numbers. Posthodiplostomum pandei is indicated by the blue line box, while the red line box shows the other Indian species.
We also want to mention that in a molecular phylogenetic study by Achatz, et al. [35], the synonymy of Posthodiplostomum, Ornithodiplostomum Dubois, 1936, and Mesoophorodiplostomum Dubois, 1936, was suggested. To the best of our knowledge, the Genbank database only contains molecular data for the following Neascus-_type species from the Indian region to date: _N. hanumanthai, N. gussevi, N. xenentodoni, and one unknown Posthodiplostomum sp. The comparison of the genetic data of our specimens with the above-mentioned species molecular level and in variations of measurements of the body parts also easily differentiates them with P. pandei.
Discussion
Previous studies show that it is difficult to evaluate the host specificity with regard to larval helminthes because of their poor morphological identification [72, 73]. But molecular methods are the best tools to deal with this problem and achieve accurate identifications of larval trematodes in fish hosts [72, 74, 75]. Larval trematodes often harbour multiple sites in their hosts, but most of their infection site preferences or specificities are likely mistaken [76, 77, 78, 79]. In a study by Locke, et al. [34], different Posthodiplostomum species were recorded from viscera and muscles. Therefore, thorough examinations of infection sites supplemented with molecular data are presently required to decide if infection sites are the same or differ between Posthodiplostomum species. In the present study, P. pandei was molecularly characterised for the first time. The placement of our specimens in the phylogenetic trees produced in this study supports their settlement within Diplostomidae and separates them from other representatives of different geographical regions and India too. Our species appeared as a sister taxon to the Posthodiplostomum species isolate that belongs to Mexico in the 18S rRNA tree, though it was far from the Indian species (KF738455), which is the only sequence available for the Channa punctata. However, in the ITS analysis, P. pandei was grouped of N. xenentodoni, both species infected the same host, X. cancila; and were fairly distinct from each other. In the distribution of species by 28S rRNA tree, it was congruent with the 18S and ITS rRNA trees in many respects, with Posthodiplostomum species of C. punctata (KF738450) appearing along with two other species, N. hanumanthai and N. gussevi from India. N. xenentodoni from the same host appeared as a sister taxon to P. pandei in the 28S rRNA tree. The phylogenetic trees generated with all molecular markers in the present study revealed that P. pandei belongs to a clade containing Posthodiplostomum species Further work is a prerequisite to establishing and understanding the Indian Posthodiplostomum species infections in fish and avian hosts that need to be molecularly characterised to determine their diversity, which might be different from what we know today.
About 26 species of Neascus-type metacercariae have been described in India, mostly based on morphology (Table 3). For many species, at this point, it is hard to establish how many species are under the genus Posthodiplostomum, which shows their unstable taxonomic history in India. The reason behind the unknown status of diplostomid parasites in India is that they have not been well studied, leaving a big gap with regard to their true diversity. However, in recent years, a few studies have pointed it out, contributed to the understanding of diplostomid parasites, and tried to fill the gap [80, 81, 82, 83, 84, 85]. Based on morphological variation between specimens, previous authors [27] recommended the description of eleven Neascus-type metacercariae species from X. cancila (Tables 1 & 3). In our study, nearly similar morphological measurements were found in the overall body measurements for P. pandei, N. hepatica, and N. srivastavi. In all eleven species reported from the host X. cancila, only P. xenentodoni (= N. xenentodoni) has molecular sequences. With the exception of P. xenentodoni, no genetic sequences for the other Indian species were available for comparison from X. cancila. Another reason that also hampers the taxonomic studies and diversity of this group of parasites in India is the restrictions of ethical guidelines that are related to definitive hosts (birds), which make it challenging to study the relationship of adults in birds along with metacercariae and the completion of their life cycles.
In the case of diplostomid parasites, García-Varela, et al. [86] recently proposed that all South American species described on the basis of metacercaria morphology be considered incertae sedis. We are agreeing with that because studies based only on the morphology of metacercariae will make it difficult to reveal the relationship with their adult forms in future studies. Therefore, we also propose that, from India, those metacercariae for which molecular data is available have been considered valid species, as their phylogenetic relationships can be easily evaluated with their adults in future studies (Table 3). Under the incertae sedis status, we believe that those Posthodiplostomum/Neascus- type metacercariae species for which only morphological data is available should be taken into account, leaving the possibility for future studies to perform morphological and molecular comparisons that help to uncover their life cycle and clarify their status.
Conclusion
Overall, our identification of P. pandei is supported by its morphology as well as at the molecular level. Furthermore, molecular data generated here will be useful in future studies to determine whether Neascus-type metacercariae species belong to Posthodiplostomum or another genus. Though we robustly acclaim that no new species can be further described without molecular data under the genus Posthodiplostomum, We also proposes that some previously published Neasus-type species that had been misidentified should be reassigned on the basis of morphological and molecular data in future studies.
Conflicts of Interest
The authors declare no competing interests.
Acknowledgements
We are grateful to the Head, Department of Zoology, Chaudhary Charan Singh University, Meerut (Uttar Pradesh), 250004, India, for providing laboratory facilities.
Funding
This research was supported by grants from the DST (Department of Science and Technology), Government of India, New Delhi, India, under the WOS-A Scheme (SR/ WOS-A/LS-382/2018) in Delhi to AC.
References
-
Ababouch L (2006) Assuring fish safety and quality in international fish trade. Mar Pollut Bull 53(10-12): 561- 568.
-
Bauer ON, Hoffman GL (1976) Helminth range extension by translocation of fish. In: Page LA (Editor), Wildlife Diseases. Springer, Boston, MA, pp: 163-172.
-
Naylor R, Burke M (2005) Aquaculture and ocean resources: Raising tigers of the sea. Annu Rev Environ Resour 30: 185-218.
-
Houston RD, Haley CS, Hamilton A, Guy DR, Mota-Velasco JC, Gheyas AA, et al. (2010) The susceptibility of Atlantic salmon fry to freshwater infectious pancreatic necrosis is largely explained by a major QTL. Heredity 105: 318- 327.
-
Hoogendoorn C, Smit NJ, Kudlai O (2019) Resolution of the identity of three species of _Diplostomum_ (Digenea: Diplostomidae) parasitising freshwater fishes in South Africa, combining molecular and morphological evidence, Int J Parasitol Parasites Wildl 11: 50-61.
-
Niewiadomska K (2002) Superfamily Diplostomoidea Poirier. In: Gibson DI, Jones A, et al. (Eds.), Keys to the Trematoda, CABI Publishing and The Natural History Museum, Wallingford-London, UK 1: 159-242.
-
Cech G, Sándor D, Molnár K, Paulus P, Papp M, et al. (2020) New record of metacercariae of the North American _Posthodiplostomum centrarchi_ (Digenea, Diplostomidae) in pumpkinseed (_Lepomis gibbosus_) in Hungary. Acta Vet Hung 68(1): 20-29.
-
Duan Y, Al-Jubury A, Kania PW, Buchmann K (2021) Trematode diversity reflecting the community structure of Danish freshwater systems: molecular clues. Parasit Vectors 14: 43.
-
Ritossa L, Flores V, Viozzi G (2013) Life-cycle stages of a _Posthodiplostomum_ species (Digenea: Diplostomidae) from Patagonia, Argentina. J Parasitol 99: 777-780.
-
Hamouda AH, Bazh E (2019) First report of _Posthodiplostomum_ _minimum_ metacercariae and resulting histopathological changes in _Bagrus bajad_ from Lake Nasser, Egypt. J Parasit Dis 43: 14-21.
-
Stoyanov B, Georgieva S, Pankov P, Kudlai O, Kostadinova A, (2017) Morphology and molecules reveal the alien _Posthodiplostomum centrarchi_ Hoffman, 1958 as the third species of _Posthodiplostomum_ Dubois, 1936 (Digenea: Diplostomidae) in Europe. Syst Parasitol 94: 1-20.
-
Lopez-Hernandez D, Locke SA, Lane de Melo A, Rabelo LME, Pinto SH (2018) Molecular, morphological and experimental assessment of the life cycle of _Posthodiplostomum nanum_ Dubois, 1937 (Trematoda: Diplostomatidae) from Brazil, with phylogenetic evidence of the paraphyly of the genus _Posthodiplostomum_ Dubois, 1936. Infect Genet Evol 63: 95-103.
-
Niewiadomska K (2003) MonografieParazytologiczne. 15. PasozytyrybPolski (klucze do oznaczania): Przywry- Digenea,PolskieTowarzystwoParazytologiczne, Warszawa, Poland, pp: 169.
-
Horak P, Kolarova L, Mikeš L (2014) Schistosomatoidea and Diplostomoidea. Adv Exp Med Biol 1154: 217-254.
-
Ondrackova M, Bartosova S, Valova Z, Jurajda P, Gelnar M (2004) Occurrence of black-spot disease caused by metacercariae of _Posthodiplostomum cuticola_ among juvenile fishes in water bodies in the Morava River basin. Acta Parasitol 49: 222-227.
-
Quist MC , Bower MR, Hubert WA, (2007) Infection by a black spot-causing species of Uvulifer and associated opercular alterations in fishes from a high-desert stream in Wyoming. Dis Aquat Org 78(2): 129-136.
-
Otachi EO (2009) Studies on Occurrence of Protozoan and Helminth Parasites in Nile Tilapia (_Oreochromis_ _niloticus_ L.) from Central and Eastern Provinces, Kenya. MSc. Thesis, Egerton University.
-
McAllister C, Tumlison R, Robison H, Trauth S (2013) Initial survey on black-spot disease (Digenea: Strigeoidea: Diplostomidae) in select Arkansas fishes. J Ark Acad Sci 67: 200-203.
-
Hoogendoorn C, Smit NJ, Kudlai O (2019) Molecular and morphological characterisation of four diplostomid metacercariae infecting _Tilapia sparrmanii_ (Perciformes: Cichlidae**)** in the north West Province, South Africa. Parasitol Res 118: 1-14.
-
Achatz TJ, Curran SS, Patitucci KF, Fecchio A, Tkach VV (2019) Phylogenetic affinities of Uvulifer Spp. (Digenea: Diplostomidae) in the Americas with description of two new species from Peruvian Amazon. J Parasitol 105: 704-717.
-
Steedman RJ (1991) Occurrence and environmental correlates of black spot disease in stream fishes near Toronto, Ontario. Trans Am Fish Soc 120: 494-499.
-
Kirankaya ŞG, Ekmekçi FG (2011) Frequency of black spot disease in _cobitis cf. turcica_ from pınarbaşı springs (Haymana, Turkey). Folia Zool 60: 350-354.
-
López-Hernández D, Locke SA, de Melo AL, Rabelo ÉML, Pinto HA (2018) Molecular, morphological and experimental assessment of the life cycle of _Posthodiplostomum nanum_ Dubois, 1937 (Trematoda: Diplostomidae) from Brazil, with phylogenetic evidence of the paraphyly of the genus _Posthodiplostomum_ Dubois, 1936. Infect Genet Evol 63: 95-103.
-
Osorio-Sarabia D, Pérez-Ponce de León G, García- Marquez LJ (1986) Helmintos de Pecesen Pátzcuaro, Michoacán II: Estudio histopatológico de la lesión causada por metacercarias de _Posthodiplostomum_ _minimum_ (Trematoda: Diplostomatidae) en el hígado de _Chirostoma estor_. Anales Inst Biol Univ Nac Autón México, Zool 57: 247-260.
-
Lane R, Morris J (2000) Biology, prevention and effects of common grubs (digenetic trematodes) in freshwater fish. Tech Bull 115: 1-6.
-
Ondračková M, Dávidová M, Gelnar M, Jurajda P (2006) Susceptibility of Prussian carp infected by metacercariae of _Posthodiplostomum cuticola_ (v. Nordmann, 1832) to fish predation. Ecol Res 21: 526-529.
-
Pandey KC, Agrawal N (2013) Metacercarial Fauna of India. Rec Zool Serv India 349: 1-310.
-
Blasco-Costa I, Poulin R, Presswell B (2017) Morphological description and molecular analyses of _Tylodelphys_ sp. (Trematoda: Diplostomidae) newly recorded from the freshwater fish _Gobiomorphus_ _cotidianus_ (common bully) in New Zealand. J Helminthol 91(3): 332-345.
-
Sereno-Uribe AL, Andrade-Gómez L, de León G, García- Varela M (2019) Exploring the genetic diversity of _Tylodelphys_ (Diesing, 1850) metacercariae in the cranial and body cavities of Mexican freshwater fishes using nuclear and mitochondrial DNA sequences, with the description of a new species. Parasitol Res 118(1): 203- 217.
-
Sitko J, Heneberg P (2021) Long-term dynamics of trematode infections in common birds that use farmlands as their feeding habitats. Parasit Vectors 14: 383.
-
Shinad K, Chaudhary A, Prasadan PK, Singh HS (2021) Phylogenetic position of _Tremiorchis ranarum_ Mehra and Negi, 1926 (Trematoda: Plagiorchiidae) with remark on this monotypic genus. Parasitol Int 84: 102398.
-
Luton K, Walker D, Blair D (1992) Comparison of ribosomal internal transcribed spacers from two congeneric species of flukes (Platyhelminthes: trematoda: Digenea). Mol Biochem Parasitol 56(2): 323- 328.
-
Hall TA (1999) BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp Ser 41: 95-98.
-
Locke SA, McLaughlin JD, Dayanandan S, Marcogliese DJ (2010) Diversity and specificity in _Diplostomum_ spp. metacercariae in freshwater fishes revealed b cytochrome c oxidase I and internal transcribed spacer sequences. Int J Parasitol 40(3): 333-343.
-
Achatz TJ, Chermak TP, Martens JR, Pulis EE, Fecchio A, et al. (2021) Unravelling the diversity of the Crassiphialinae (Digenea: Diplostomidae) with molecular phylogeny and descriptions of five new species. Curr Res Parasitol Vector Borne Dis 1: 100051.
-
Locke SA, Van Dam A, Caffara M, Pinto HA, López- Hernández D, et al. (2018) Validity of the Diplostomoidea and Diplostomida (Digenea, Platyhelminthes) upheld in phylogenomic analysis. Int J Parasitol 48(13): 1043- 1059.
-
Campos A, Cummings MP, Reyes JL, Laclette JP (1998) Phylogenetic relationships of platyhelminthes based on 18S ribosomal gene sequences. Mol Phylogenet Evol 10(1): 1-10.
-
Moszczynska A, Locke S, McLaughlin J, Marcogliese D, Crease T (2009) Development of primers for the mitochondrial cytochrome c oxidase I gene in digenetic trematodes (Platyhelminthes) illustrates the challenge of barcoding parasitic helminths. Mol Ecol Res 9: 75-82.
-
Heneberg P, Sitko J, Těšínský M (2020) Paraphyly of _Conodiplostomum_ Dubois, 1937. Parasitol Int 76: 102033.
-
Nguyen TC, Li YC, Makouloutou P, Jimenez LA, Sato H (2012) _Posthodiplostomum_ sp. metacercariae in the trunk muscle of northern snakeheads (_Channa argus_) from the Fushinogawa River, Yamaguchi, Japan. J Vet Med Sci 74: 1367-1372.
-
Athokpam VD, Tandon V (2014) Morphological and molecular characterization of _Posthodiplostomum_ sp. (Digenea: Diplostomidae) metacercaria in the muscles of snakeheads (_Channa punctata_) from Manipur, India. Helminthologia 51: 141-152.
-
Lovy J, Friend SE (2017) Phylogeny and morphology of _Ovipleistophora diplostomuri_ n. sp. (Microsporidia) with a unique dual-host tropism for bluegill sunfish and the digenean parasite _Posthodiplostomum minimum_ (Strigeatida). Parasitology 144(14): 1898-1911.
-
Hernandez-Mena DI, Garcia-Varela M, Perez-Ponce de Leon G (2017) Filling the gaps in the classification of the Digenea Carus, 1863: systematic position of the Proterodiplostomidae Dubois, 1936 within the superfamily Diplostomoidea Poirier, 1886, inferred from nuclear and mitochondrial DNA sequences. Syst Parasitol 94(8): 833-848.
-
Choudhary K, Tripathi A, Pandey KC, Agrawal N (2017) Morphological and molecular characterisation of three Indian Neascus-type metacercariae (Digenea: Diplostomidae), Parasitol Int 66(5): 652-659.
-
Sokolov SG, Gordeev II (2020) Molecular and morphological characterisation of flatworm larvae parasitizing on fish in Cat Tien National Park, Vietnam. Nat Conserv Res 5: 19-30.
-
Perez-Ponce de Leon G, Sereno-Uribe AL, Pinacho- Pinacho CD, Garcia-Varela M (2021) Assessing the genetic diversity of the metacercariae of _Posthodiplostomum_ _minimum_ (Trematoda: Diplostomidae) in Middle American freshwater fishes: one species or more? Parasitology 149: 239-252.
-
Sereno-Uribe AL, Andrade-Gómez L, Pérez Ponce de León G, García-Varela M (2019) Exploring the genetic diversity of _Tylodelphys_ (Diesing, 1850) metacercariae in the cranial and body cavities of Mexican freshwater fishes using nuclear and mitochondrial DNA sequences, with the description of a new species. Parasitol Res 118(1): 203-217.
-
Tamura K, Stecher G, Kumar S (2021) MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol 38: 3022-3027.
-
Milne I, Lindner D, Bayer M, Husmeier D, McGuire G, (2009) TOPALi v2: a rich graphical interface for evolutionary analyses of multiple alignments on HPC clusters and multi-core desktops, Bioinformatics 25(1): 126-127.
-
Rai P, Pande BP (1964) On the Metacercarial fauna of the Indian fresh water scombresocid fish _Belone cancila._ Indian J Fish 11: 315-320.
-
Chakrabarti KK (1970) Two new species of Strigeid metacercariae from an Indian fresh water fish, _Xenentodon_ _cancila_ (Ham.). Proc Helminth Soc Washington 37: 5-10.
-
Pandey KC (1971) Studies on metacercaria of freshwater fishes of India. VIII. On two new species of _Neascus_ Hughes, 1927. Proc Indian Acad Sci 43: 78-83.
-
Baugh SC, Chakrabarti KK (1977) On a triple infection of strigeid metacercaria in an Indian freshwater fish _Xenentodon cancila._ Angew Parasitol 18(2): 84-94.
-
Agrawal N, Khan N (1982) Studies on some strigeid _(Neascus)_ metacercariae from fishes of Lucknow. Uttar Pradesh J Zool 2: 32-40.
-
Pandey KC, Tewari SK (1986) Studies on few strigied metacercariae, Part I. Ad Bio 5: 21-31.
-
Pandey KC, Pandey N (2000) Studies on two new strigeid metacercariae from _Xenentodon cancila_ (Ham.). Indian J Helminth (N.S.) 17: 29-32.
-
Gupta BK, Shukla SK (2019) Studies on larval digenetic trematode structure of some new species of Strigeids from food fishes of U.P. and its adjacent state. Indian J Pure Appl Biosci 4(4): 129-135.
-
Vidyarthi RD (1937) Studies on the family Diplostomidae Poirier, Part I. Two new parasites of the genus _Diplostomulum_ from Indian carnivorous birds. Proc Nat Acad Sci India 7: 22-28.
-
Kaw BL (1950) Studies in Helminthology: Helminth parasites of Kashmir. Part I. Trematoda. Indian J Helminth 2: 67-126.
-
Khera S (1958) _Neascus chelai_ nov. sp. (Trematoda) from the fish _Chela clupeoides._ Anales de la escuela Nat Gen Biol 9: 1-4.
-
Fotedar DN, Raina MK (1965) On a new species of the trematode genus _Posthodiplostomum_ Dubois, 1936, from _Milvus migrans lineatus_ (Gray), common Kite in Kashmir. Kashmir Sc 1: 64-69.
-
Thapar GS (1967) Some new larval strigeids causing black spot disease in fishes from Tungabhadra dam. Indian J Helminth 19: 173-182.
-
Singh RN (1956) Studies on a new strigeid metacercaria parasitic in cyprinoid fishes. Proc Nat Acad Sci India 26: 255-263.
-
Pandey KC (1970) Studies on metacercaria of freshwater fishes of India. III On two new species of _Diplostomulum_ Brandes, 1892. Proc Ind Acad Sci India 42: 162-170.
-
Vankara AP, Mani G, Vijayalakshmi C (2011) A report on various digenetic metacercariae from the freshwater fishes of River Godavari, Rajahmundry. J Parasit Dis 35: 177-185.
-
Gupta NK, Mishra PN (1974) On a new trematode _Posthodiplostomum_ _mehtai_ new species family diplostomatidae from _Milvus migrans_ at Shimla, Himachal. Indian J Zool 2: 23-25.
-
Pandey KC (1973) Studies on metacercariae of freshwater fishes of India. II. Description of a known and five unknown stregeid metacercariae from Lucknow. Indian J Zool 14: 155-166.
-
Chakrabarti KK (1974) Studies on some metacercariae of the Indian fresh water fishes _Channa punctatus_ (Bloch) and C. _striatus_ (Bloch). Rev Iber Parasitol 34: 57-80.
-
Dhanukumari CH (1994) A new species of strigeid metacercaria, _Neascus_ _punctatusi_ (Trematoda: Diplostomatidae) from an Indian freshwater fish, _Channa_ _punctatus_ (Bloch). Riv Di Parasitol 11: 245-249.
-
Verma SC (1936) Studies on the family Bucephalidae (Gasterostomata). Part I. Descriptions new forms from Indian freshwater fishes. Proc Nat Acad Sci 6: 66-89.
-
Madhavi R, Rukmini C (1997) Observations on the life cycle of _Posthodiplostomum grayii_ (Verma, 1936) (Trematoda: Diplostomatidae). J Parasit Appl Anim Biol 6: 93-102.
-
De León GP, Nadler SA (2010) What we don’t recognize can hurt us: a plea for awareness about cryptic species. J Parasitol 96: 453-464.
-
Perkins SL, Martinsen ES, Falk BG (2011) Do molecules matter more than morphology? Promises and pitfalls in parasites. Parasitology 138: 1664-1674.
-
Pérez-Ponce De León G, García-Varela M, Pinacho- Pinacho CD, Sereno-Uribe AL, Poulin R (2016) Species delimitation in trematodes using DNA sequences: Middle- American _Clinostomum_ as a case study. Parasitology 143: 1773-1789.
-
Soldánová M, Georgieva S, Roháčová J, Knudsen R, Kuhn JA, et al. (2017) Molecular analyses reveal high species diversity of trematodes in a sub-Arctic lake. Int J Parasitol 47(6): 327-345.
-
Hoffman GL (1999) Parasites of North American Freshwater Fishes. Cornell University Press, Ithaca, New York, USA, pp: 539.
-
Poulin R (2011) Uneven distribution of cryptic diversity among higher taxa of parasitic worms. Biol Lett 7(2): 241-244.
-
Boone EC, Laursen JR, Colombo RE, Meiners SJ, Romani MF, et al. (2018) Infection patterns and molecular data reveal host and tissue specificity of _Posthodiplostomum_ species in centrarchid hosts. Parasitology 145: 1458- 1468.
-
Díaz Pernett SC, Brant SV, Locke SA (2022) First integrative study of the diversity and specificity of metacercariae of _Posthodiplostomum_ Dubois, 1936 from native and introduced fishes in the Caribbean. Parasitology 149: 1894-1909.
-
Choudhary K, Tripathi A, Pandey KC, Agrawal N (2017) Morphological and molecular characterisation of three Indian Neascus-type metacercariae (Digenea: Diplostomidae). Parasitol Int 66(5): 652-659.
-
Chaudhary A, Gupta S, Tripathi R, Singh HS (2017) Morphological and molecular analyses of _Tylodelphys_ spp. metacercaria (Trematoda: Diplostomidae) from the vitreous humour of two freshwater fish species, _Channa_ _gachua_ (Ham.) and _Puntius sophore_ (Ham.). Vet Parasitol 244: 64-70.
-
Rajvanshi S, Choudhary K, Agrawal N (2016) Threading: A novel insilico indagation method for genetic characterization of some diplostomoid metacercariae (Digenea: Diplostomidae Poirier, 1886). Exp Parasitol 171: 71-76.
-
Chaudhary A, Gupta S, Verma C, Tripathi R, Singh HS (2017) Morphological and Molecular Characterization of Metacercaria of _Tylodelphys_ (Digenea: Diplostomidae) from the Piscine Host, _Mystus tengara_ from India. J Parasitol 103: 565-573.
-
Chaudhary A, Tripathi R, Gupta S, Singh HS (2017) First report on molecular evidence of _Tylodelphys cerebralis_ (_Diplostomulum cerebralis_) Chakrabarti, 1968 (Digenea: Diplostomidae) from snakehead fish _Channa punctate_. Acta Parasit 62: 386-392.
-
Chaudhary A, Singh K, Sharma B, Singh HS (2021) Description of the Metacercaria of _Cardiocephaloides_ sp. (Digenea, Diplostomoidea), newly recorded from the brain of Gangetic Leaffish (_Nandus nandus_) and its genetic characterization in India. Acta Parasit 66(4): 1597-1604.
-
García-Varela M, Sereno-Uribe A, Pinacho-Pinacho C, Hernández-Cruz E, Pérez-Ponce de León G (2016) An Integrative taxonomic study reveals a new species of _Tylodelphys_ Diesing, 1950 (Digenea: Diplostomidae) in central and northern Mexico. J Helminthol 90(6): 668- 679.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework