On Pseudoglaridacris laruei (Lamont, 1921) (Cestoda: Caryophyllidae) from Wisconsin, USA: A Review and Perspectives
Specimens of Pseudoglaridacris laruei (Lamont, 1921) Oros, Uhrovič, Scholz, 2018 (Caryophyllidae) were first described as Caryophyllaeus laruei Lamont, 1921, redescribed as Glaridacris laruei Hunter, 1927, and subsequently designated as Pseudoglaridacris laruei Oros, Uhrovič, Scholz, 2018, were collected and examined from the white sucker Catostomus commersoni Lacépède and the lake chubsucker Erimyzon sucetta Lacépède in Silver and Tichigan lakes, SE Wisconsin. We update our first and only 1986 redescription of this Wisconsin population, with new observations and measurements, and compare with those in the original and other descriptions. The morphology of P. laruei is revisited using light microscopy revealing new features. Emphasis is placed on the morphological variability of its scolex and reproductive structures throughout development using many microscopic images. Measurements of 100 specimens and the re-examination of 25 more specimens from both lakes provided an opportunity to review the taxonomy and description of the species, distinguish locality differences, and correct and update other statements and concepts by other observers.
Introduction
Mackiewicz JS [1] reviewed the morphology, anatomy, development, zoogeography, and systematics of the Caryophyllidea van Beneden (Cestoidea) and indicated the existence of 89 species and 37 genera in these monozoic cestodes. Mackiewicz JS [2] provided perspectives on the evolution, biology and zoogeography of 128 species in 45 genera of caryophyllids from the Nearctic, Neotropical, Palearctic, Ethiopian, Oriental, and Australian regions. Mackiewicz JS [3] later recognized about 150 species in 41 genera worldwide (except for the Neotropical Region). Of these, 14 genera and 90 species belonging to 3 families were described from the Indo-Malayan region from Bagridae, Heteropneustidae, Schilbeidae and Siluridae, cypriniformes, and cobitid fishes [3]. Ash A, et al. [4, 5] invalidated a large number of inadequately described species from clariid and catfishes in India and neighboring countries. Most recently, however, Scholz, et al. [6] and Scholz, et al. [7] recognized only 117 species and 46 genera in order Caryophyllidea Carus, 1863 parasitizing teleost fish (Cypriniformes, Siluriformes, and some catfish). Chubb JC [8] reviewed the seasonal ecology of the Caryophyllidea in the various climate zones of the world.
Amin OM [9, 10, 11, 12] recognized 15 species in 8 families of caryophyllid cestodes mostly from suckers (Catostomidae) in lake and river systems in Arizona and Wisconsin.
These included Pseudoglaridacris laruei (Lamont, 1921) Oros, Uhrovič, Scholz, 2018 for Caryophyllaeus laruei Lamont, 1921 and Glaridacris laruei Hunter, 1927. Oros, Uhrovič, Scholz, 2018 redescribed the genus Glaridacris and created a new genus, Pseudoglaridacris with 3 species: Pseudoglaridacris laruei (Lamont, 1921) Oros, Uhrovič, Scholz, 2018, Pseudoglaridacris oligorchis (Haderlie, 1953) Oros, Uhrovič, Scholz, 2018, and Pseudoglaridacris confusa (corrected to confusus [13]) Oros, Uhrovič, Scholz, 2018. See collections, following, for details. It is important to note here that the order Caryophyllidea includes 4 families (Balanotaeniidae, Capingentidae, Caryophyllaeidae, Lytocestidae) discussed by Mackiewicz JS [1, 3] and reviewed in Amin OM, et al. [13].
In Wisconsin, Amin OM [9, 12] gave an account of P. laruei in 2 lakes, Silver and Tichigan lakes, and distinguished between it and other accounts elsewhere [14, 15, 16]. In his Ph.D. dissertation, Mackiewicz [15] noted that P. laruei was the most common species found in C. commersoni in New York and reviewed reports that were based on its erroneous identification from various hosts confusing it with Glaridacris confusus.
Materials, Collections and Methods
Collections
Seasonal collections of 403 specimens of P. laruei from the white sucker C. commersoni and the lake chubsucker E. sucetta in Silver and Tichigan lakes, SE Wisconsin were made in April, 1976, April and November, 1977 and April, 1984. A total of 376 specimens of P. laruei were collected from 6 of 10 (60%) examined white suckers in Silver Lake (mean 37.6, maximum 263), and 21 specimens from 7 of 105 (7%) examined white suckers in Tichigan Lake (mean 0.2, maximum 8). In E. sucetta, a total of 5 cestodes were collected from 3 of 116 (3%) examined lake chubsuckers (mean 0.04, maximum 3) in Silver Lake and 1 cestode from 1 of 2 (50%) examined chubsuckers (mean 0.5, maximum 1) in Tichigan Lake.
The lakes
Silver Lake is a land-locked eutrophic natural lake (Kenosha County) (42°32′58″N,88°10′9″W). Tichigan Lake is a relatively large eutrophic impoundment on the Fox River which is a tributary of the Mississippi River in Racine County (42°48′20″N,88°13′00″W).
Methods
After capture and transfer to the laboratory in coolers on ice, the fish were dissected within a few hours after capture. Upon recovery, worms were placed at once in warm 70% ethanol then processed and stained with Semichon’s carmine. Worms were then processed in ascending concentrations of ethanol for dehydration reaching 100%. Worms were ultimately cleared in oil of winter green before whole mounting in Canada balsam. Measurements are in mm or micrometers as noted in Table 1; the range is followed by the mean values between parentheses. Width measurements represent maximum width. Microscope images were created using 10X and 40X objective lenses of a BH2 light Olympus microscope (Olympus Optical Co., Osachi- shibamiya, Okaya, Nagano, Japan) attached to an AmScope 1000 video camera (United Scope LLC, DBA AmScope, Irvine, California), linked to an ASUS lab top equipped with HDMI high-definition multimedia interface system (Taiwan- USA, Fremont, California). Images from the microscope were transferred from the labtop to a USB and stored for subsequent processing on a computer. We found microscope images to be considerably more informative than schematic line drawings as they depict the natural appearance of anatomical structures.
Results
Amin [12] gave morphological notes of P. laruei as Glaridacris laruei, from the Wisconsin population collected from white suckers C. commersoni and the lake chubsuckers E. sucetta captured in Silver and Tichigan lakes, SE Wisconsin between 1976 and 1984. He [12] made initial taxonomic comparisons with specimens from other collections in Wisconsin and Michigan [14, 16, 17, 18]. We have restudied specimens in our personal collection. Measurements of about 100 specimens were made in 1967-1968 [12] and new measurements and observations of 25 additional specimens from the original collection were recently made and added to the earlier measurements creating combined measurements included in Table 1. We provide below a brief revised description of our qualitative morphological findings and a detailed comparative morphometrical data (Table 1) high-lightening extreme variations from other descriptions by Lamont [18], Hunter [14], and Williams [16].
| Locality | Lake Mendota & Green Lake, Wisconsin | Green L. & Lake Mendota, Wisconsin; Douglas L., Michigan | Red Cedar River, NW Wisconsin | Silver Lake, SE Wisconsin | Tichigan Lake, SE Wisconsin |
|---|---|---|---|---|---|
| Hosts (N) | Catostomus commersoni | Catostomus commersoni | Catostomus commersoni, Hypentelium nigricans, Moxostoma macrolepidotum | Catostomus commersoni | Erimyzon sucetta |
| Specimens measured | 24+ collected | 15 | 112, 19, 12 | 119 | 6 |
| Authority | Lamont (1921) | Hunter (1927) | Williams (1980) | Amin (1986) & this paper | |
| Characters | Combined | ||||
| Body length (mm) | 7 | 4.2-9.5 | 3.8-13.2 (6.2-6.9) | 3.8-9.6 (6.2) | 5.9-12.2 (9.5) |
| Width at neck | --- | 0.4-0.55 | 0.35-0.70 (0.54) | 0.8 | |
| Width at gonopore (mm) | 0.8 | 0.5-0.73 | 0.41-0.94 (0.60-0.69) | 0.40-0.84 (0.64) | 0.48-1.04 (0.74) |
| Scolex | Primitive, with terminal disc & 6 loculi (Figure 2) | II-type with 6 loculi & terminal 0.055 mm disc 0.06 mm thick | Disc occasionally absent, With 6 loculi, 75-100% bothrioloculodiscate | Terminally hexagonal disc frequently absent, largely bothrioloculodiscate scolex often with 6 poorly defined loculi | |
| Length of scolex (mm) | --- | 0.3-0.75 | --- | 0.25-0.87 (0.53) | 0.4 |
| Width of Scolex (mm) | --- | 0.5-0.6 | --- | 0.36-0.92 (0.67) | 0.48-0.76 (0.63) |
| Vitellaria | Oval-elongate in 2 lateral rows | Confined in 2 lateral rows | --- | In 2 lateral rows bordering median testes. | |
| Vitellaria length (µm) | 100-200 | 94-229 (154) | 146-208 (182) | ||
| Vitellaria diameter (µm) | 30-50 | 62-135 (92) | 73-83 (71) | ||
| Testes (T) number | 74 *(Figure2) in 2 median rows | 60-85 (70) globular- lobate in 1-3 median rows | 0-109(51-93) | 60-122 (99) + 1 protogynous worm Usually in 2 rows | 70-112 (97) Usually in 2 rows |
| Testis diameter (µm) | --- | 120-180 (146) | --- | 80-208 (133) | 122-208 (150) |
| First T to ant. tip (mm) | --- | --- | --- | 0.64-1.56 (1.03) | 0.84-1.88 (129) |
| First pre-ovarian vitellarium to ant. tip (mm) | --- | --- | --- | 0.52-1.52 (0.94) | 0.68-1.64 (1.19) |
| % of worm length | --- | --- | --- | 12-27 (16) | 7-13 (11) |
| Post-gonopore distance (mm) | --- | --- | --- | 0.60-1.60 (0.92) | 0.84-1.76 (1.41) |
| % of worm length | --- | --- | --- | 11-18 (15) | 12-14 (13) |
| Cirrus sac diameter (µm) | --- | 108-120 | 94-154 (109-115) | 126-168 (137) | 126-224 (171) |
| No. of post- ovarian vitellaria | --- | 0-7 (1.9-4.4) | 0-5 (2.6) | 1-4 (5.4; 1 worm with 15 vitellaria) | |
| Ovary shape | Lobate, H-shaped | Lobate, H-shaped | 0-5% inverted A-shaped | Lobate, 7% inverted A-shaped. Most of these worms lack post-ovarian vitellaria | |
| Ovarian arms’ length (µm) | --- | 500-700 | --- | 350-1,400 (720) | 728-1,162 (933) |
| Distance between ovarian arms (µm) | --- | 500-550 | --- | 300-1,025 (665) | 1,070 |
| Shape of posterior end | 50% pointed or rounded | Usually bluntly pointed | |||
| Eggs | --- | Ovoid, non- operculate | --- | Small, ovoid, thin shelled, non- operculate | |
| Egg length (µm) | --- | 39-42 | --- | 38-58 (45) | 38-48 (43) |
| Egg diameter (µm) | --- | 26-30 | --- | 26-38 (32) | 26-35 (32) |
Table 1: Comparative morphometrics of Pseudoglaridacris laruei from Catostomus commersoni, Erimyzon sucetta, Hypentellium nigrica
Bolded figures indicate numbers markedly different among relevant populations. Table 1: Comparative morphometrics of Pseudoglaridacris laruei from Catostomus commersoni, Erimyzon sucetta, Hypentellium nigricans and Moxostoma macrolepidotum in various lakes and the Red Cedar River of Wisconsin and in Michigan.
Brief Morphological Description of Adult Cestodes from Wisconsin
Body small-medium with distinct neck (Figs. 9,13), widest at gonopore (Figs. 19-24). It is often dorso-ventrally flattened, covered with thin cuticle enclosing prominent cortical parenchyma and inner and weaker outer longitudinal muscles. Measurements and counts in Table 1. Scolex somewhat bothrioloculodiscate, wider than long, widest near base with fairly parallel sides (II-shaped), tapering anteriorly to a frequently absent terminally hexagonal disc cut by 6 often poorly-demarcated loculi separated by frequently indefinite ribs (Figures 9,11-14). Younger specimens with anteriorly cone-shaped proboscis, no terminal disc and undefined loculi (Figures 1 & 6) as well as with incipient testes and vitellaria (Figure 3) and underdeveloped ovary and uterus (Figures 2,4,5). Scolexes of younger adults with poorly defined terminal disc and loculi (Figures 7,8,10). Testes spheroidal, large, in 2 medium parallel rows occasionally appearing with 3 rows where partially crowded. Testes developing first before vitelline glands (Figures 3,15- 18) and mature adults with testicular field occupying less space than vitellaria anteriorly and posteriorly. Cirrus sac small, round, near ventral surface (Figure 19). Vitelline glands smaller than testes, ovoid-elongate, multi-layered in 2 lateral fields (Figures 3,15-18), extending posteriorly to anterior end of ovarian arms. Vitellaria extend dorsally and ventrally about the edge of testicular field (Figure 18). Post-ovarian vitellaria few, often 1-3 (Figures 19- 22,24,26-29) or absent (Figures 23,25,30). Male and female reproductive systems do not open into common atrium. Vas deference detectable about middle of testicular field ventrally passing into muscular ejaculatorius at seminal vesicle then reaching cirrus sac dorsally and anteriorly at about center of medullary parenchyma. Distinctly lobate ovary H-shaped with long slender converging posterior arms appearing to join when tips of posterior ovarian arms link with few post-ovarian vitellaria often confused with ovarian lobules creating appearance of an inverted A-shaped ovary (Figure 19-29). Rarely, however, true inverted A-shaped ovary distinguishable especially with occasional absence of post-ovarian vitellaria (Figure 30). Ovarian commissure fanning at juncture with ovarian arms and tapering towards center (Figures 19,21). Vagina running posteriorly from female cloaca into receptaculum seminis then posteriorly to anterior and dorsal aspects of ovarian commissure. Uterine coils not extending anterior to cirrus sac but posteriorly reaching to posterior tip of posterior ovarian arms [19, 20, 21, 22, 23, 24, 25]. Posterior end usually conical (Figures 21,23,25,27), rarely rounded [20, 24] with slight terminal middle invagination where excretory vesicle and pore open tremino-ventrally. Egg small, ovoid, thin-shelled, non-operculate.
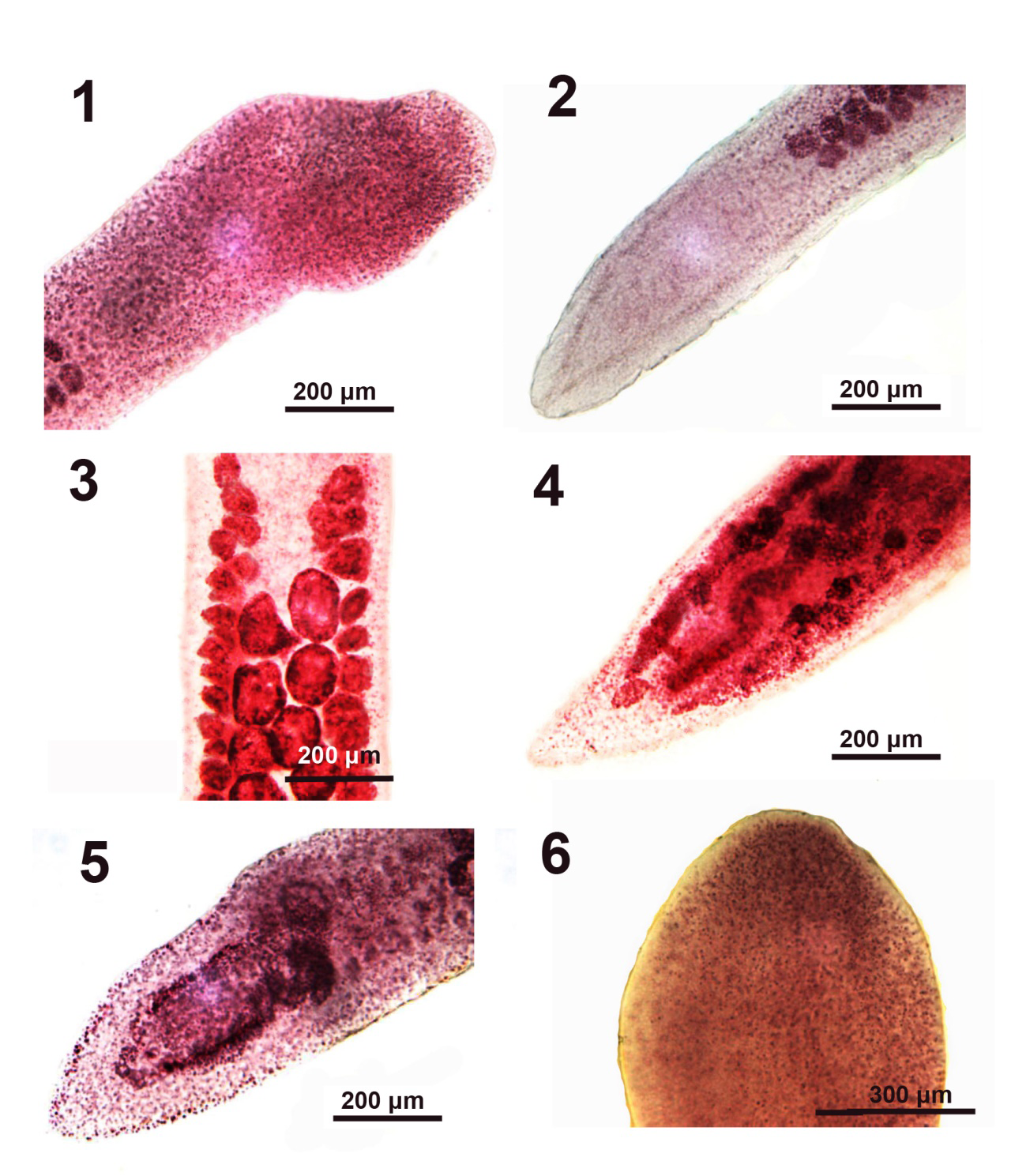
Figures 1-6: Microscope images of developing Pseudoglaridacris laruei from Catostomus commersoni in Tichigan Lake, Wisconsin. 1, 2. The anterior and posterior portions of one small 3.1 mm long juvenile with underdeveloped scolex and reproductive system but with testes. 3, 4. The mid-section and posterior portion of one young maturing 5.6 mm long specimen showing underdeveloped vitellaria and posterior displacement of more developed testes (Figure 3) and incipient female reproductive system with barely developing ovarian arm and uterus (Fig. 4). 5. The posterior portion of another specimen with barely developing ovary comparable to that in Fig. 4. 6. A developing scolex of an older juvenile showing undefined loculi and no terminal disc.
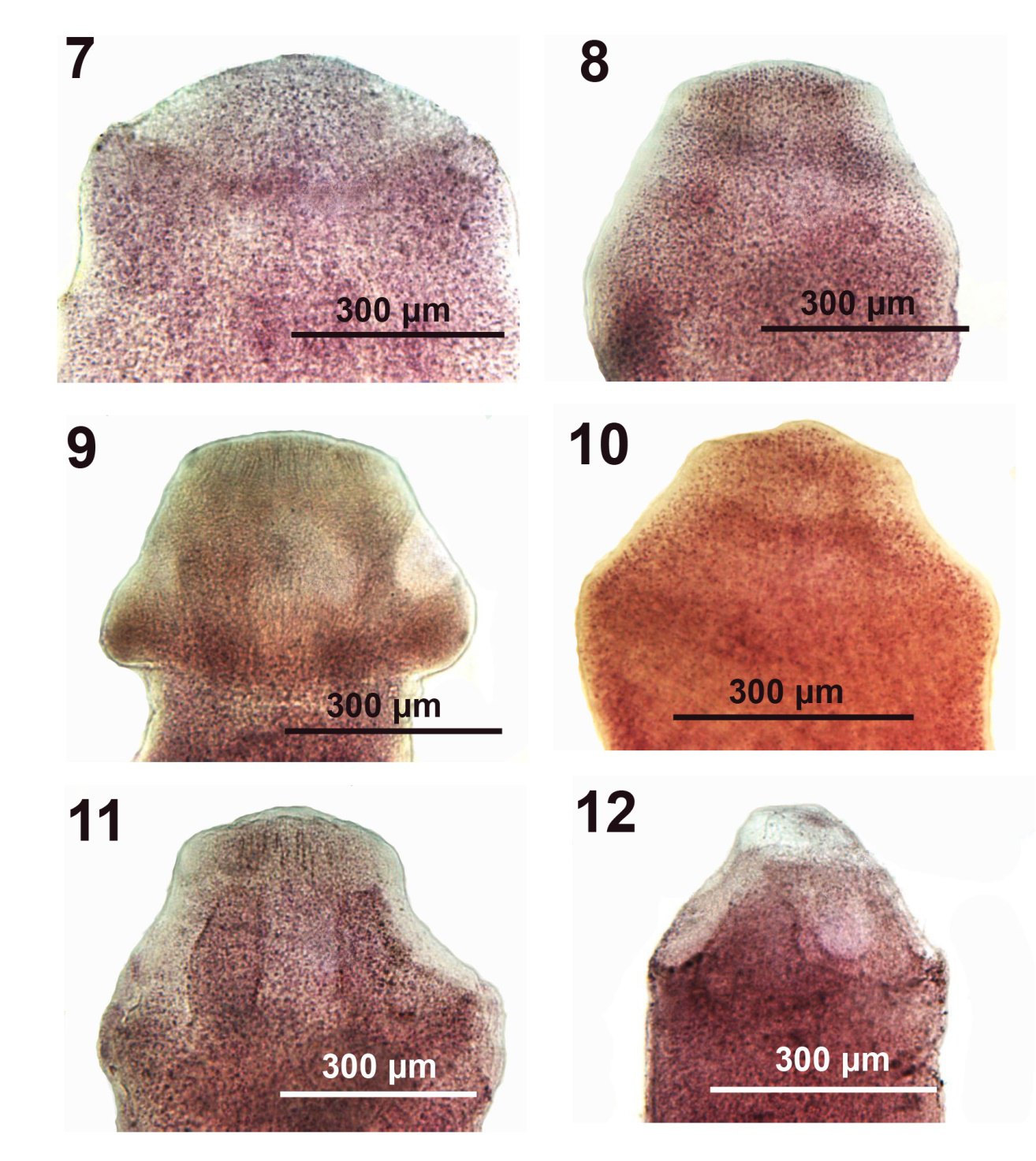
Figures 7-12: Microscope images of the scolexes of adult Pseudoglaridacris laruei from Erimyzon sucetta (Figure 7) and Catostomus commersoni (Figures 8-12) in Tichigan Lake, Wisconsin. Figures show variations in the demarcation of the scolex from anterior trunk (Figure 9) and the expression of loculi (Figures 9,11,12). The terminal disc is marginally developing only in Figures 9,11 and possibly Figure12. Magnification is to the same scale in all figures.
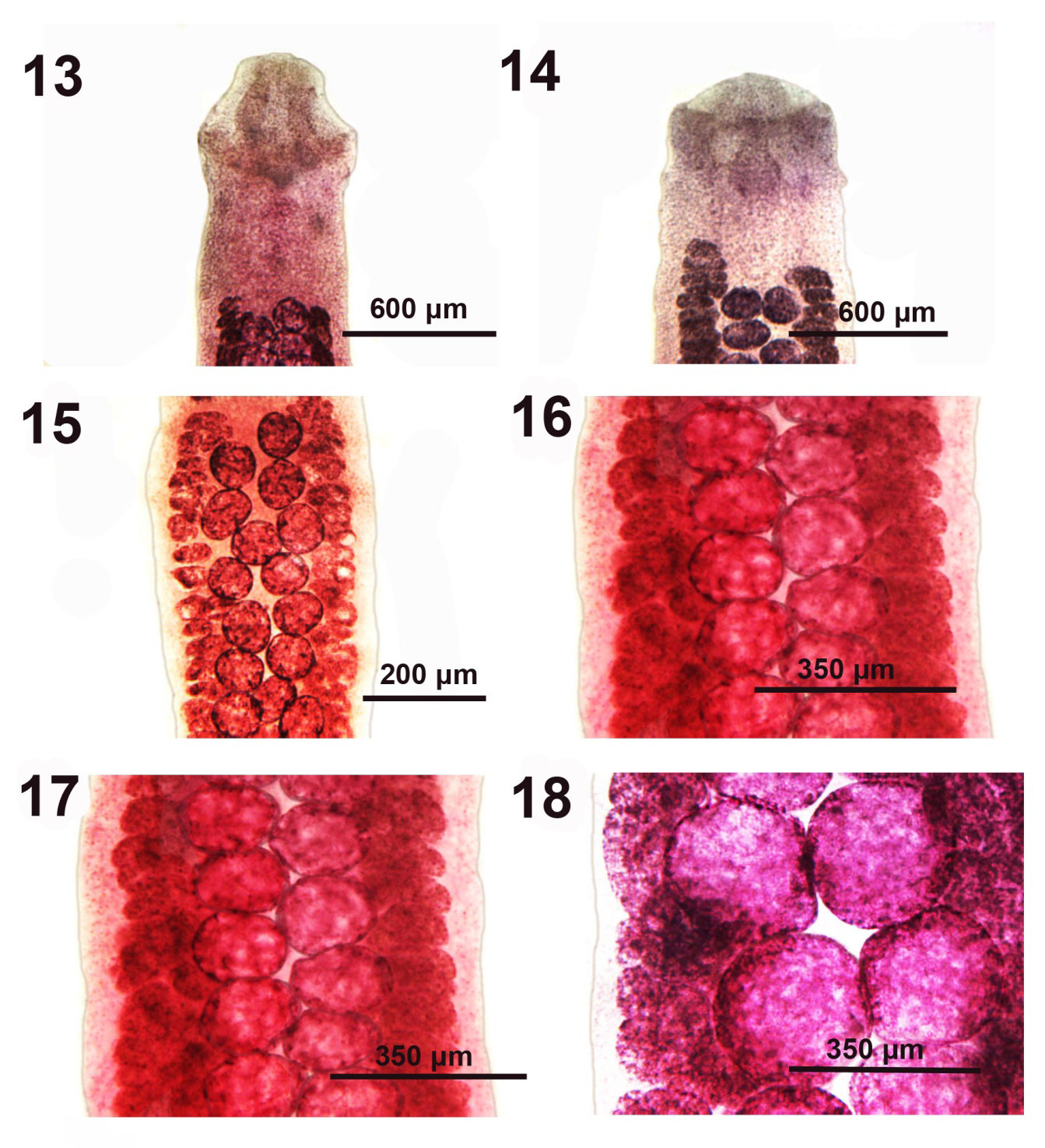
Figures 13-18: Microscope images of the anterior portion (Figures 13 & 14) and the mid-section (Figures 15-18) of adult Pseudoglaridacris laruei from Catostomus commersoni in Tichigan Lake, Wisconsin. 13, 14. The anterior portions of 2 younger adults showing some delineation of scolex loculi with a suggestion of developing terminal disc. Note the anterior extension of vitellaria compared to testes especially in Figures 14. 15. A normal mid-section of a young adult specimen showing the relative size and shape of the vitellaria compared to the testes in 10X magnification. 16-18. Mid-sections of 3 fully developed adult worms similar to that in Figure15 all at the same magnification at 40X. Note the variation in size of testes and the extreme large size and detail of testes in Figure 18.
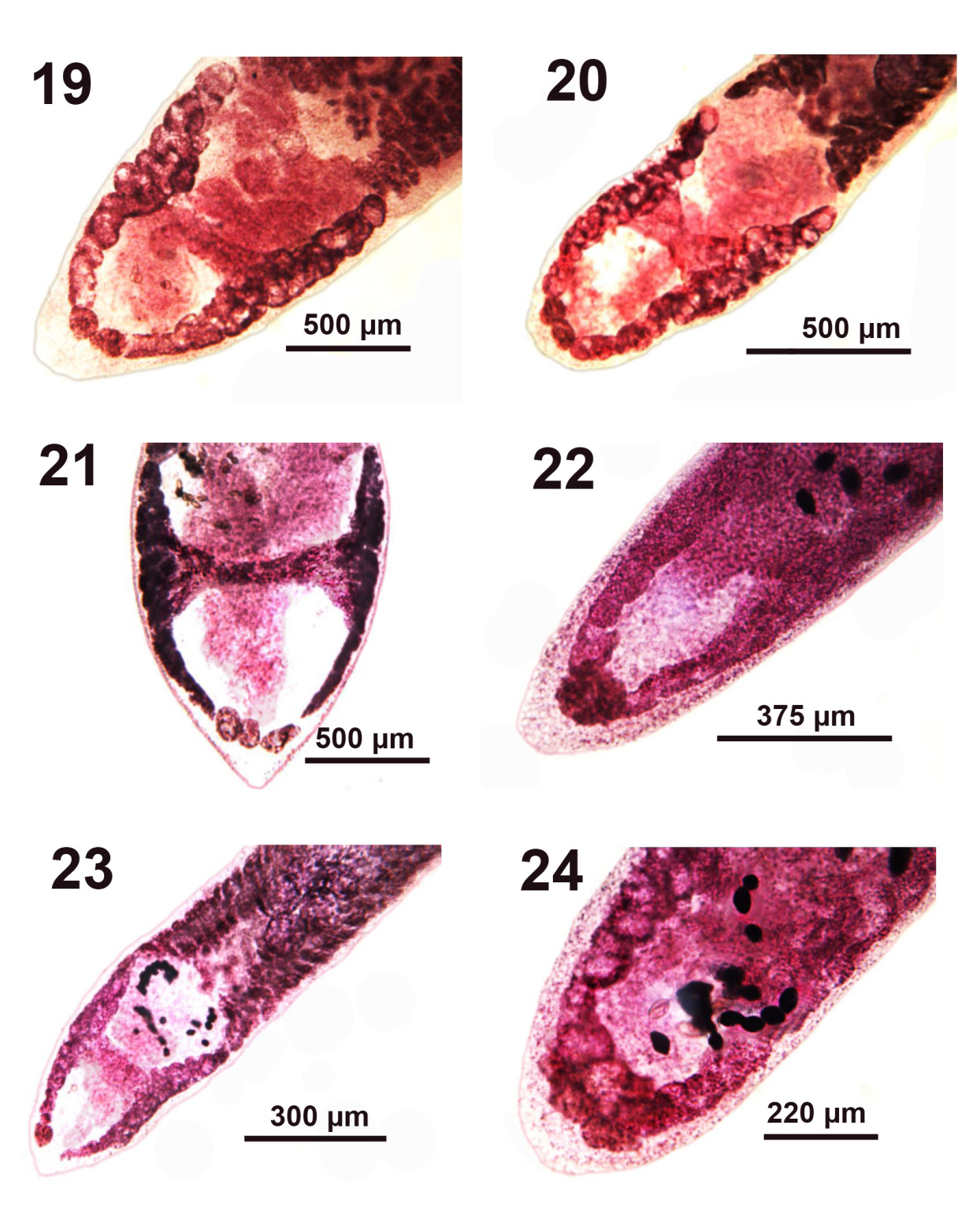
Figures 19-24: Microscope images of the posterior portion of adult Pseudoglaridacris laruei from Catostomus commersoni in Tichigan Lake, Wisconsin. Variations in the shape and organization of the H-shaped ovary in 6 different adult specimens. Note that the long posterior ovarian arms of P. laruei converge to produce what appears to be an inverted A-shaped ovary when posteriorly conjoined with a few post-ovarian vitelline glands resembling ovarian lobules (Figures 19,20,24).
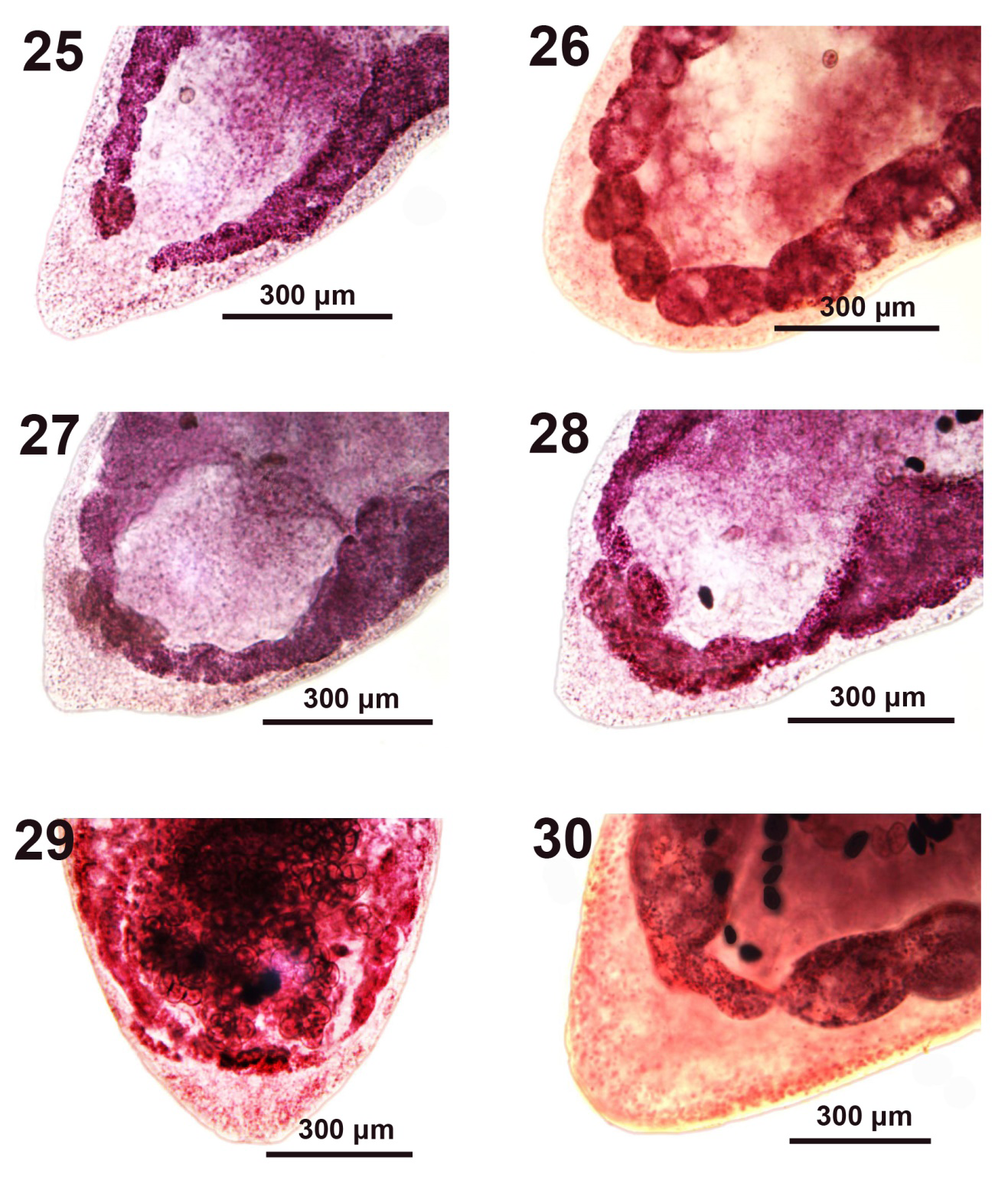
Figures 25-30: Microscope images of the posterior portion of adult Pseudoglaridacris laruei from Catostomus commersoni in Tichigan Lake, Wisconsin. More variations in the shape and organization of the H-shaped (Figures 25-29) and inverted A-shaped (Figure 30) ovaries in adult specimens. Note the curving inward of the long posterior limb of ovarian arms and the position of the very few post-ovarian vitellaria creating the impression of an inverted A-shaped ovary (Figures 26-28).
Taxonomic Summary
- Type Host: White sucker Catostomus commersoni Lacépède
- Other Hosts: Lake chubsucker Erimyzon sucetta Lacépède, northern hogsucker Hypentelium nigricans (Lesueur), shorthead redhorse Moxostoma macrolepidotum (Lesueur) (Table 1)
- Type Locality: Lake Mendota, Wisconsin
- Other Localities: Green, Silver, and Tichigan lakes, Wisconsin; Douglas Lake, Michigan; Red Cedar River, NW Wisconsin (Table 1)
- Deposited specimens: US National Museum Helminthological collections nos. 78655, 78656 & Milwaukee Public Museum coll. no. IZ985-01B.
Remarks
The original description [17] was limited in scope but Lamont’s Figure 2 of a whole worm made up for much lost detail. It (Figure 2) showed a well-developed scolex with a terminal disc and 6 loculi, 74 large almost globular testes receding short of the field occupied by the compact vitelline glands anteriorly and posteriorly, H-shaped ovarian arms converging and connected posteriorly by a few post-ovarian vitellaria, and uterine coils not reaching past the cirrus sac anteriorly but extending to near posterior end of the worm. Lamont [17], however, over simplified her treatment, missed much detail and made a few errors such as stating that “This primitive scolex has four poorly defined suckers with an obscure terminal disk at the anterior end” (p. 2). True, in our specimens the scolex has poorly defined loculi and an obscure terminal disc but, however, it has 6 and not 4 loculi. Hunter [14] emphasized the deficiencies in Lamont’s description and provided novel observations and interpretations of unusual features. For example, he also acknowledged the “irregularly marked loculi … beneath the disc” (p. 70). Hunter [14] further defined the fibers of the outer longitudinal muscles as “infrequently found in the neck just under the basement membrane” (p. 71). Hunter [14] identified 8 pairs of ascending and descending excretory canals and reported 2 rows of testes but occasionally observed 1 or 3. Hunter [14] characterized the II-shaped scolex as belonging to Glaridacris different from that of Isoglaridacris hexacotyle in not being pointed distally and from G. catostomi in not being wedge-shaped. The scolex of both I. hexacotyle and G. catostomi shared 6 loculi with P. laruei but lacked its terminal disc. Hunter GW [14] also noted than P. laruei has the smallest cirrus sac and eggs, and that “the male and female reproductive systems do not open into a common genital atrium” (p. 75).
In our specimens, the scolexes may assume diverse shapes with the terminal disc protruding as Hunter [14]
previously observed. Hunter [14] speculated that in fully contracted scolexes, the terminal disc alone shows and that “the remainder of the scolex appears swollen laterally when the parallel longitudinal muscles within the scolex (are) drawn down inside by the action of the strong inner longitudinal muscles.” Hunter GW [14] also observed that the terminal disc may be absent, as we did, as it can be “pulled in between the bands or due to degeneration of the scolex before fixation” (p. 75). The typical II-shaped scolex characteristic of P. laruei was attributed to the action of the specialized muscles flattening the distal end [14].
Our specimens exhibited the usual variations in the shape of scolex, the number of testes and post-ovarian vitellaria as often observed by Mackiewicz JS [15] and Williams DD [16]. Specimens from C. commersoni, however, had smaller cirrus sac, and ovarian arms, shorter post-gonopore distance, and fewer post-ovarian vitellaria compared to those from E. sucetta. Our specimens from C. commersoni varied from those of Hunter GW [14] and Williams [16] who reported fewer testes and smaller cirrus sac (Table 1). In New York, Mackiewicz JS [15] concluded that P. laruei was the most common species of caryophyllids found in C. commersoni. In the Red Cedar River, Wisconsin, Williams DD [16] reported P. laruei from 2 additional species of suckers, Hypentelium nigricans (Lesueur) and Moxostoma macrolepidotum (Lesueur) and compared its morphometrics (Table 1) with those of Pseudoglaridacris oligorchis (Haderlie, 1953) Oros, Uhrovič, Scholz, 2018. Williams [16] presented 17 informative line drawings of immature, nongravid and gravid specimens from all hosts covering a considerable range of morphological variations.
Non-taxonomic issues from our local Wisconsin material included the persistent localization of worms in posterior intestinal sites, late summer early autumn recruitment and spring maturation, and localization in concurrent infections [12]. Other issues related to host and geographical distribution, misidentifications, size allometry, studies of scolex and neck, scolex glands, histopathology, and calcareous corpuscles were summarized by Hoffman GL [17].
Discussion
The taxonomic status of Caryophyllaeus laruei Lamont, 1921 [18] has not been challenged until Hunter [14] redescribed it as Glaridacris laruei and Oros M, et al. [19] erected the genus Pseudoglaridacris Oros, Uhrovič, Scholz, 2018 and included 3 former members of Glaridacris in it: Pseudoglaridacris laruei (Lamont, 1921) Oros, Uhrovič, Scholz, 2018, Pseudoglaridacris confusa Oros, Uhrovič, Scholz (later corrected by Amin OM and Rubtsova N [20] to P. confusus), and Pseudoglaridacris oligorchis (Haderlie, 1953) Oros, Uhrovič, Scholz [19]. Oros M, et al. [19] diagnosed the new genus, differentiated it from Glaridacris, provided systematic summaries and remarks but did not redescribe or measure these 3 species. Pseudoglaridacris was distinguished from Glaridacris in having compact rather than follicular ovary, bothrioloculodiscate rather than cuneiform scolex, genital atrium rather than well separated gonopores, lateral position of vitelline follicles rather than with median follicles, and a smaller size body [19]. We accept the designation of Pseudoglaridacris with one qualification based on our specimens of P. laruei from Wisconsin. In our specimens: the male and female gonopores ARE separate even though they may appear to be in a genital atrium depending on the state of contraction of the specimen.
The taxonomic implications of allometric growth at different stages of development of a number of helminth parasites especially caryophyllid cestodes have been reported. Fischthal JH, et al. [21] showed growth of the post-vitelline region of P. laruei from the C. commersoni in New York comparable to that of other helminths like P. confusus. In P. confusus, allometric growth was observed to proceed at a considerably greater rate in regions containing reproductive structures posterior to first vitellaria compared to pre-vitelline region of scolex and neck. Hunter GW [14] demonstrated this phenomenon from a study of 106 specimens of P. confusus in 7 length classes reaching 20 mm long worms. Hunter’s [14] pioneering concept applied also to other caryophyllid as well as pseudophyllid examples. Amin OM [10] also studied growth patterns in Isoglaridacris hexacotyle (Linton, 1897) Mackiewicz, 1968 from Catostomus insignis Baird & Girard in the lower Salt River, Tempe, Arizona, and demonstrated the acceleration of growth of the post -vitelline region, the region of reproductive activity, compared to the pre-vitelline region including the scolex and neck, as well as growth in the scolex, cirrus sac, and in width at the common gonopore region. We have additionally observed the same phenomenon in non-tapeworm helminths also enhancing the increased reproductive potential of worms. Amin OM [10] also studied a total of 136 metacercariae of Clinostomum marginatum Rudolphi, 1819 collected from 17 C. insignis and 4 C. clarkii Baird & Girard in the Upper Salt River, Arizona, placed in 6 size classes ranging from 2.49 to 6.04 mm in length. The hind body posterior to the constriction at the level of the ventral sucker including the developing reproductive structures grew at a considerably faster rate than the fore body anterior to the ventral sucker.
Conclusion
We conclude our treatment of P. laruei from Wisconsin fishes with our evaluation of the morphological variability of 5 Arizona and Wisconsin caryophyllid cestodes originally collected and reported by Amin. Other species in this series include Isoglaridacris hexacotyle (Linton, 1897) Mackiewicz, 1968 from Arizona [22] and Biacetabulum biloculoides Mackiewicz [23], Glaridacris catostomi Cooper, 1920 [24], and Pseudoglaridacris confusus (Hunter, 1929) Oros, Uhrovič, Scholz [13] from Wisconsin. In all cases, we have reported the expanded range of qualitative variations in the morphology of the scolex and reproductive structures that varied with host species and developmental stages as has been well documented for P. laruei. The greater relative growth of worm parts housing the reproductive structures compared to other parts of the developing worm body was shown to have taxonomic implications in I. hexacotyle and G. laruei. Scolexes in developing juveniles assume immature anteriorly tapering morphology lacking defined terminal disc or loculi different from the scolexes of adults reported in original descriptions. The degree of variations in the size or number of testes by host species or geographical locations varied in many species but was especially greatest in P. laruei the adults of which differed from those of all other species by their II-shaped scolex with 6 well defined loculi and a classical H-shaped ovary. The long posterior ovarian arms of P. laruei converge to produce what appears to be an inverted A-shaped ovary when posteriorly conjoined with a few post-ovarian vitelline glands resembling ovarian lobules. Variabilities in other anatomical structures of all five cestode species were also presented by Amin, et al. [13, 22, 23, 24, 25] accounting for much of the full range of measurements and shapes of attachment and reproductive structures that have not been previously published, and introduced new ones that may have not been previously accounted for. In addition, we included among the pivotal features of taxonomic importance in our descriptions of all species such characters as the distance between the male and female gonopores, the earlier development of the testes compared to vitellaria as in P. laruei, the lateral vs. the medullary fields of distribution of vitellaria, size of cirrus sac and eggs and whether the latter are operculated, the degree of development of inner vs. outer longitudinal muscles, the number of post-ovarian vitellaria, the distance between anterior-most vitellaria and scolex tip, the makeup of the ovary (whether lobate or granular), and the anterior and posterior extensions of uterine coils. Some of these characters are of taxonomic importance at the generic level such as the makeup of ovarian tissue, and the distance between the male and female gonopores and whether they are in a common genital atrium. Measurements of some of these structures were not reported in the original descriptions and other descriptive accounts but are all included for our reported Arizona and Wisconsin specimens.
Acknowledgments
This project was supported by an Institutional Grant from the Parasitology Center, Inc. (PCI), Scottsdale, Arizona.
Declarations
Compliance with ethical standards
Conflict of Interest
The authors declare no conflicts of interest or competing interests.
Ethical Approval
The authors declare that they have observed all applicable ethical standards.
Availability of Data
All presented and related data are available by contacting the senior author.
References
-
Mackiewicz JS (1972) Caryophyllidea (Cestoidea): a review. Exper Parasitol 31(3): 417-512.
-
Mackiewicz JS (1982) Caryophyllidea (Cestoidea): Perspect Parasitol 84: 397-417.
-
Mackiewicz JS (1994) Order Caryophyllidea van Beneden in Carus, 1863. In: Khalil LF, Jones A, et al. (Eds.), Keys to the cestode parasites of vertebrates. CAB Intern, Wallingford, UK, pp: 21-43.
-
Ash A, Scholz T, Oros M, Kar PK (2011) Tapeworms (Cestoda: Caryophyllidea), parasites of Clarias batrachus (Pisces: Siluriformes) in the Indomalayan region. J Parasitol 97(3): 435-459.
-
Ash A, Scholz T, Oros M, Levron C, Kar PK (2011b) Cestodes (Caryophyllidea) of the stinging catfish Heteropneustes fossilis (Siluriformes: Heteropneustidae) from Asia. J Parasitol 97(5): 899-907.
-
Scholz T, Kuchta R, Oros M (2021) Tapeworms as pathogens of fish: A review. J Fish Dis 44(12): 1883- 1900.
-
Scholz T, Kuchta R (2017) A digest of fish tapeworms. Vie et milieu - Life and environment 67(2): 43-58.
-
Chubb JC (1982) Seasonal occurrence of helminths in freshwater fishes Part IV. Adult Cestoda, Nematoda and Acanthocephala. Adv parasitol 20: 1-292.
-
Amin OM (1968) Helminth fauna of Suckers (Catostomidae) of the Gila River System, Arizona. PhD Thes Ariz St Univ (No 68-1644) Univ Microfilms. Ann Arbor, Michigan; Dissert Abstr 28: 3521.
-
Amin OM (1969) Helminth fauna of suckers (Catostomidae) of the Gila River System, Arizona. II. Five parasites from _Catostomus_ spp. Am Midland Nat 82(2): 429-443.
-
Amin OM (1974) Intestinal helminthes of the white sucker, _Catostomus commersoni_ (Lacépède), in SE Wisconsin. Proc Helminthol Soc Wash 41(1): 81-88.
-
Amin OM (1986) Caryophyllaiedae (Cestoda) from lake fishes in Wisconsin with a description of _Isoglaridacris_ _multivitellaria_ sp. n. from Erimyzon sucetta (Catostomidae). Pro Helminthol Soc Wash 53: 48-58.
-
Amin OM and Rubtsova NYu (2022) Reexamining _Pseudoglaridacris confusus_ (Hunter, 1929) nom. nov. (Cestoda: Caryophyllidae) from buffalo fish, Ictiobus spp. in Arizona, USA. Int J Zoo Animal Biol 5(6): 000420.
-
Hunter III GW (1930) Studies on the Caryophyllaeidae of North America Illinois Biological Monographs 11 (1927) pp: 556.
-
Mackiewicz JS (1960) Studies on the Caryophyllaeidae (Cestoidea) of _Catostomus commersoni_ (Lacépède), with emphasis on those fish near Ithaca, New York (Tompkins County), USA PhD thesis, Cornell Univ, Ithaca, New York, USA, pp: 391.
-
Williams DD (1980) Caryophyllidian tapeworms. Report on fauna and flora of Wisconsin No. 17. Museum of Natural History, Stevens Point, Wisconsin, USA, pp: 24.
-
Hoffman GL (1999) Parasite of North American freshwater fishes. Comstock Publishing Assoc, Cornell Univ Press, Ithaca, New York, USA, pp: 539.
-
Lamont ME (2017) Two new parasitic flatworms. Occasional papers of the Museum of Zoology, Univ. Michigan 3: 1-6.
-
Oros M, Uhrovič D, Scholz T (2018) A new classification of _Glaridacris_ Cooper, 1920 (Cestoda: Caryophyllidea), parasites of suckers (Catostomidae) in North America, including erection of _Pseudoglaridacris_ n. gen. J Parasitol 104(1): 60-69.
-
Amin OM and Rubtsova NYu (2022) Reexamining _Pseudoglaridacris confusus_ (Hunter, 1929) nom. nov. (Cestoda: Caryophyllidae) from buffalo fish, Ictiobus spp. in Arizona, USA. Int J Zoo Animal Biol 5(6): 000420.
-
Fischthal JH, Carson DO, Vaught RS (1982) Size allometry of the caryophyllidean cestode _Glaridacris laruei_ from the white sucker. J Parasitol 68(6): 1175-1177.
-
Amin OA, Rubtsova NY (2022) Revisiting _Isoglaridacris_ _hexacotyle_ (Linton, 1897) Mackiewicz, 1968 (Cestoda: Caryophyllidae) from Catostomid Fishes in Rivers of Arizona. Int J Zoo Animal Biol 5(4): 000392.
-
Amin OA, Rubtsova NY(2022) A Morphological Study and Description of _Biacetabulum biloculoides_ Mackiewicz and Mccrae, 1965 (Cestoda: Caryophyllidae) From Catostomid Fishes in Lakes and Rivers of Wisconsin, With Special Notes on Concurrent Infections. Int J Zoo Animal Biol 5(4): 000395.
-
Amin OA and Rubtsova NY (2022) The Morphology and Description of _Glaridacris catostomi_ Cooper, 1920 (Cestode: Caryophyllidae) from Catostomid Fishes in Lakes and Rivers of Wisconsin, with Morphological Observations. Int J Zoo Animal Biol 5(5): 000402.
-
Amin OM and Rubtsova N Yu (2022) Reexamining _Pseudoglaridacris confusus_ (Hunter, 1929) nom. nov. (Cestoda: Caryophyllidae) from buffalo fish, Ictiobus spp. in Arizona, USA. Int J Zoo Animal Biol 5(6): 000420.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework