Lyme Disease: A Zoonosis Tick-Borne Borrelia Bacterium [2/4]
Lyme disease is present worldwide but is most prevalent in the temperate regions of the hemisphere, where it is the primary vector-borne illness. Every year, numerous individuals in Europe and the United States receive a diagnosis and undergo treatment for Lyme disease. The transmission of tick-borne diseases is a complex process, and it is challenging to identify the factors that contribute to their spread
Editorial
Lyme disease is present worldwide but is most prevalent in the temperate regions of the hemisphere, where it is the primary vector-borne illness. Every year, numerous individuals in Europe and the United States receive a diagnosis and undergo treatment for Lyme disease. The transmission of tick-borne diseases is a complex process, and it is challenging to identify the factors that contribute to their spread. Epidemiology provides a means of analyzing the distribution of Lyme disease within a population and identifying risk factors. The epidemiology of infectious Lyme disease is essential for implementing preventive healthcare. The spatiotemporal dynamics of Lyme disease are associated with the spatiotemporal factors of humans, infected vector ticks, and infected hosts. The only environmental variable consistently linked to an increased risk and incidence of Lyme disease is the presence of forests. Therefore, forests are the primary variable that raises the risk of Lyme disease [1, 2].
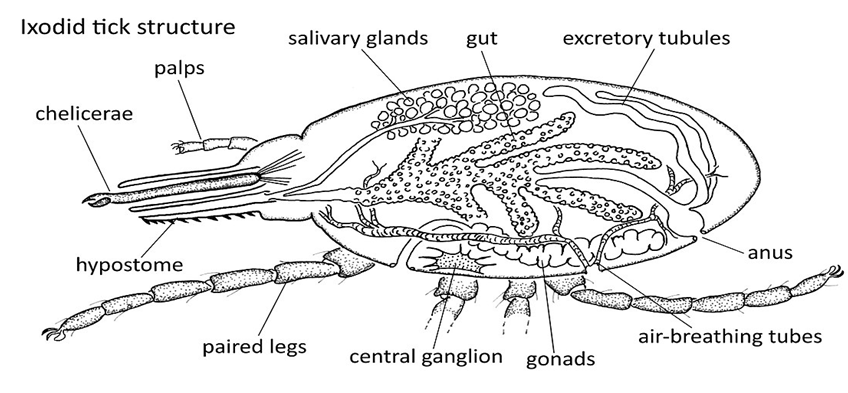
Ticks are ectoparasites of vertebrate animals and are the primary vectors of Lyme disease. Ticks have a unique body structure consisting of a fusion of the opisthosoma with the prosoma. The gnathosoma is the head region and contains mouthparts, two palps, two chelicerae, and a hypostome. The idiosoma, or body, includes the legs, digestive tract, reproductive organs, bears sclerites, and the gonopore. Ticks legs contain Haller’s organ, which detects odors, temperature, moisture, and vibrations in the environment to locate potential hosts [3, 4]. The tick also possesses a cephalic rostrum, a perforating organ composed of the hypostome and chelicerae, which function as macerator organs. Ticks have specialized mouthparts that allow them to insert their hypostome into the host’s skin and extract blood [5, 6]. When feeding on blood, ticks release an anticoagulant and anti- inflammatory substance to bite the host painlessly. Ticks transmit pathogenic infections, including bacteria, viruses, and protozoa [7, 8]. There are three families of ticks: Ixodidae, Argasidae, and Nuttalliellidae. The primary vectors of Lyme disease are ticks from the family Ixodidae, specifically the genus Ixodes (Figure 1).
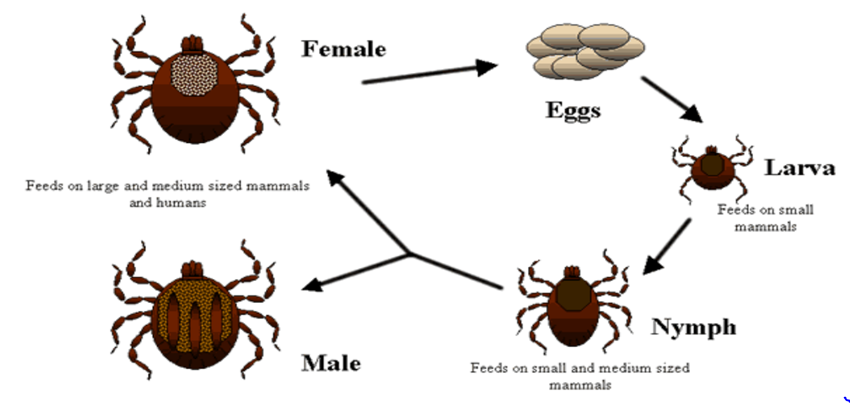
Ticks female lays eggs on the ground, and these eggs hatch into larvae that survive by feeding on the blood of animals. These larvae then mature into nymphs and feed on animal blood before molting and transforming into adult ticks. Once feeding, adult ticks mate, and the female lays eggs in the environment. Depending on the species, Ixodidae ticks may have a one-host, two-host, or three-host lifecycle [9, 10]. The lifecycle of the genus Ixodes is closely related to climatic conditions like temperature and humidity. Tick activity in Europe generally peaks between May and October, while in the Mediterranean region, tick activity is highest between November and January (Figure 2). Ticks are ectoparasites of various animal species. Ixodes ticks infest a variety of hosts, including amphibians, reptiles, birds, ruminants, and carnivores. Germ reservoirs for Lyme disease are generally rodents, birds, and deer. The larvae and nymphs of these ticks feed on rodents, birds, and Leporidae, while the adult females typically feed on great animals. Infestation of ticks on an infected host occurs during a blood meal. The Borrelia burgdorferi spirochetes move in the tick’s intestinal microvilli and salivary glands of ticks. Transmission of the disease occurs when ticks bite the host during a blood meal [11, 12].
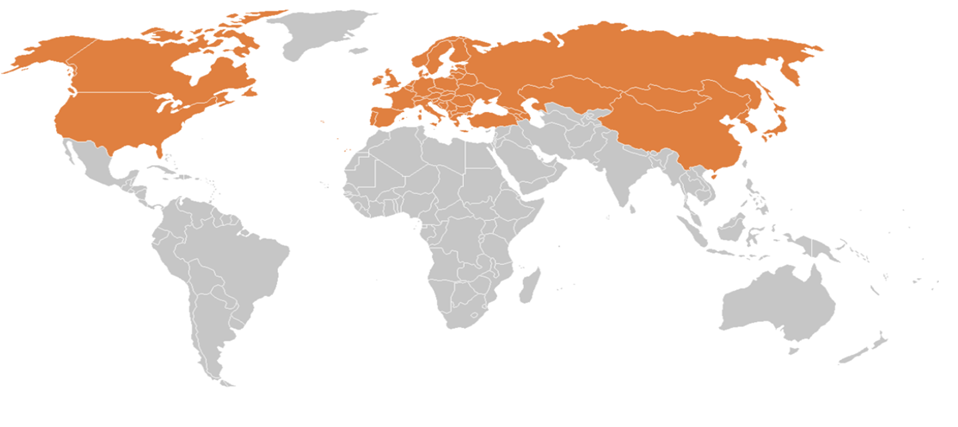
Environment affects geographical distribution, host diversity, and specificity of ticks. The molecular tools have made it easier to detect bacteria transmitted by arthropod vectors, such as ticks, that cause Lyme borreliosis. Ixodes ricinus ticks mainly transmit Lyme borreliosis, which is responsible for cases of the disease found in almost every country on the continent. Lyme borreliosis is an arthropod- borne disease that emerges and re-emerges in temperate regions of the northern hemisphere (Figure 3). It is the most prevalent vectorial disease in Europe and North America. Lyme borreliosis is a bacterial zoonosis that evolves with the changes in society, climate, and lifestyle. Factors such as human demographics, economic development, and microbial adaptation are involved in the spread of the disease [13, 14]. Studies conducted in Tunisia, Algeria, and Morocco have revealed the presence of Ixodes ricinus in fresher and more humid areas of North Africa. Lyme borreliosis occurs in this region based on clinical and serological features. Polymerase chain reaction (PCR) assays have confirmed the existence of Lyme borreliosis in Africa [15, 16]. In Asia, Lyme borreliosis- infected ticks are increasingly being found in Japan, as well as in northwest China, Nepal, Thailand, and far eastern Russia. The Borrelia species responsible for Lyme disease in northeastern China resemble those found in the far east of Russia and Japan. The clinical features of Lyme borreliosis differ from continent to continent due to the different genospecies in animal hosts [17, 18]. Lyme borreliosis, a zoonotic disease spread by ticks, is prevalent in Europe and North America. The primary cause of the disease in Europe is the Borrelia burgdorferi infection transmitted by the Ixodes ricinus tick. Adult females are more susceptible than nymphs and adult males, respectively. Furthermore, significant variations exist in infection rates across European regions, with Central Europe showing the highest rates [19, 20, 21]. The prevalence of Lyme borreliosis in ticks attached to pet dogs in the United Kingdom has served as an indicator of human exposure to infected ticks [22]. Lyme disease, the most prevalent vector-borne infection in North America, has been present in these regions for thousands of years before European settlements. It is considered the most common zoonosis transmitted by vectors in temperate zones globally and is an emerging infectious disease in Canada [23, 24]. In the United States, Lyme disease is one of the fastest-growing infectious diseases, and the geographic extent of endemic areas has dramatically increased. The population of Lyme disease-carrying ticks has reemerged from relict foci that have persisted since pre-colonial times. Tick-borne diseases are also becoming more recognized and prevalent in South America [25, 26].
These editorials highlight the clinical, epidemiology, and diagnosis of tick-borne pathogenic Borrelia. Ticks’ role in transmitting Lyme disease is significant, so highlighting the infectious agents tick-borne to humans and animals is primordial [27]. There are two barriers to tick-borne Lyme disease, the host’s immunity to tick bites and the tick’s immunity to pathogens [28]. In Lyme disease, exposure of the host’s immune system to the bacterium Borrelia induces chronic immune disease. Many targeting molecules have a role in modulating the immunity system against Lyme disease [29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45].
Accurate and rapid diagnosis with high sensitivities is one of the challenges in the medical field of infectious diseases for quick treatment in infected patients. Detection of the humoral immune response to Borrelia burgdorferi infection depends on detecting the antibody response. The diagnosis of Lyme disease is still a significant concern.
Acknowledgments
The author acknowledges Mrs. Norri Zahra and Mr. Regragui Moumaris. The author thinks Nisen Abuaf and Said
Youssouf Chanfi (Sorbonne University). The author thinks Jean-Michel Bretagne (AP-HP). The author thinks Marie- Hélène Maës and Monique Abuaf (16th district of Paris).
References
-
Killilea ME, Swei A, Lane RS, Briggs CJ, Ostfeld RS (2008) Spatial dynamics of lyme disease: a review. Ecohealth 5(2): 167-195.
-
Little EAH, Anderson JF, Stafford KC, Eisen L, Eisen RJ, et al. (2019) Predicting spatiotemporal patterns of Lyme disease incidence from passively collected surveillance data for Borrelia burgdorferi sensu lato-infected Ixodes scapularis ticks. Ticks Tick Borne Dis 10(5): 970-980.
-
Mitchell RD, Zhu J, Carr AL, Dhammi A, Cave G, et al. (2017) Infrared light detection by the haller’s organ of adult american dog ticks, Dermacentor variabilis (Ixodida: Ixodidae). Ticks Tick Borne Dis 8(5): 764-771.
-
Koloski CW, Cassone BJ (2022) Transcriptional profiling of Dermacentor variabilis (Acari: Ixodidae) provides insights into the role of the Haller’s organ in spatial DEET recognition. Ticks Tick Borne Dis 13(1): 101827.
-
Richter D, Matuschka FR, Spielman A, Mahadevan L (2013) How ticks get under your skin: insertion mechanics of the feeding apparatus of Ixodes ricinus ticks. Proc Biol Sci 280(1773): 20131758.
-
Vancová M, Bílý T, Šimo L, Touš J, Horodyský P, et al. (2020) Three-dimensional reconstruction of the feeding apparatus of the tick Ixodes ricinus (Acari: Ixodidae): a new insight into the mechanism of blood-feeding. Sci Rep 10(1): 165.
-
Pańczuk A, Tokarska-Rodak M, Plewik D, Paszkiewicz J (2019) Tick exposure and prevalence of Borrelia burgdorferi antibodies among hunters and other individuals exposed to vector ticks in eastern Poland. Rocz Panstw Zakl Hig 70(2): 161-168.
-
Sanchez E, Vannier E, Wormser GP, Hu LT (2016) Diagnosis, Treatment, and Prevention of Lyme Disease, Human Granulocytic Anaplasmosis, and Babesiosis: A Review. JAMA 315(16): 1767-1777.
-
Steere AC, Coburn J, Glickstein L (2004) The emergence of Lyme disease. J Clin Invest 113(8):1093-1101.
-
Hodzic E (2015) Lyme Borreliosis: Is there a preexisting (natural) variation in antimicrobial susceptibility among Borrelia burgdorferi strains?. Bosn J Basic Med Sci 15(3): 1-13.
-
Alcalay M, Debiais F (1994) Lyme disease. Rev Prat 44(12): 1611-1619.
-
Zhang YK, Zhang XY, Liu JZ (2019) Ticks (Acari: Ixodoidea) in China: Geographical distribution, host diversity, and specificity. Arch Insect Biochem Physiol 102(3): e21544.
-
Higgins R (2004) Emerging or re-emerging bacterial zoonotic diseases: bartonellosis, leptospirosis, Lyme borreliosis, plague. Rev Sci Tech 23(2): 569-581.
-
Philippon A (2006) Bacterial zoonoses: emerging concepts. Bull Acad Natl Med 190(3): 579-594.
-
Bouattour A, Ghorbel A, Chabchoub A, Postic D (2004) Lyme borreliosis situation in North Africa. Arch Inst Pasteur Tunis 81(1-4): 13-20.
-
Sarih M, Jouda F, Gern L, Postic D (2003) First isolation of Borrelia burgdorferi sensu lato from Ixodes ricinus ticks in Morocco. Vector Borne Zoonotic Dis Fall 3(3): 133-1339.
-
Li M, Masuzawa T, Takada N, Ishiguro F, Fujita H, et al. (1998) Lyme disease Borrelia species in northeastern China resemble those isolated from far eastern Russia and Japan. Appl Environ Microbiol 64(7): 2705-2759.
-
Ni XB, Jia N, Jiang BG, Sun T, Zheng YC, et al. (2014) Lyme borreliosis caused by diverse genospecies of Borrelia burgdorferi sensu lato in northeastern China. Clin Microbiol Infect 20(8): 808-814.
-
Rizzoli A, Hauffe H, Carpi G, Vourc H G, Neteler M, et al. (2011) Lyme borreliosis in Europe. Euro Surveill 16(27): 19906.
-
Strnad M, Hönig V, Růžek D, Grubhoffer L, Rego ROM (2017) Europe-Wide Meta-Analysis of Borrelia burgdorferi Sensu Lato Prevalence in Questing Ixodes ricinus Ticks. Appl Environ Microbiol 83(15): e00609- 00617.
-
Smith R, Takkinen J (2006) Lyme borreliosis: Europe- wide coordinated surveillance and action needed? Euro Surveill 11(6): E060622.1.
-
Smith FD, Ballantyne R, Morgan ER, Wall R (2012) Estimating Lyme disease risk using pet dogs as sentinels. Comp Immunol Microbiol Infect Dis 35(2): 163-167.
-
Ogden NH, Lindsay LR, Morshed M, Sockett PN, Artsob H (2009) The emergence of Lyme disease in Canada. CMAJ 180(12): 1221-1224.
-
Ogden NH, St-Onge L, Barker IK, Brazeau S, Bigras- Poulin M, et al. (2008) Risk maps for range expansion of the Lyme disease vector, Ixodes scapularis, in Canada now and with climate change. Int J Health Geogr 7: 24.
-
Hoen AG, Margos G, Bent SJ, Diuk-Wasser MA, Barbour A, et al. (2009) Phylogeography of Borrelia burgdorferi in the eastern United States reflects multiple independent Lyme disease emergence events. Proc Natl Acad Sci USA 106(35): 15013-15018.
-
Margos G, Tsao JI, Castillo-Ramírez S, Girard YA, Hamer SA, et al. (2012) Two boundaries separate Borrelia burgdorferi populations in North America. Appl Environ Microbiol 78(17): 6059-6067.
-
Ben Said M, Diaz Sanchez S, Bastos A, Silaghi C (2022) Editorial: Current Knowledge on Pathogenic and Endosymbiotic Tick-Borne Bacteria. Front Vet Sci 9: 900510.
-
Tabor AE, de Miranda Santos IKF, Boulanger N (2021) Editorial: Ticks and Host Immunity-New Strategies for Controlling Ticks and Tick-Borne Pathogens. Front Immunol 12: 796558.
-
Moumaris M (2022) Lyme Disease: A Zoonosis Tick- Borne Borrelia Bacterium [1/4]. Int J Zoo Animal Biol 5(6): 000425.
-
Moumaris M, Bretagne JM, Abuaf N (2020) Nanomedical Devices and Cancer Theranostics. The Open Nanomedicine and Nanotechnology Journal 6: 1-11.
-
Moumaris M, Bretagne JM, Abuaf N (2019) Biological Membranes and Malaria-Parasites. The Open Parasitology Journal 7: 1-18.
-
Moumaris M, Bretagne JM, Abuaf N (2018) Hospital Engineering of Medical Devices in France. The Open Medical Devices Journal 6: 10-20.
-
Moumaris M, Rajoely B, Abuaf N (2015) Fluorescein Isothiocyanate-Dextran can track Apoptosis and Necrosis induced by heat shock of Peripheral Blood Mononuclear Cells and HeLa Cells. Open Biological Sciences Journal 1: 7-15.
-
Moumaris M, Rajoely B, Abuaf N (2012) The Naïve B Cells are the Lymphocytes with the Highest Anionic Phospholipid Binding Ratios. The Open Immunology Journal 5: 27-35.
-
Moumaris M (2008) Imagerie par résonance magnétique à l’Hôtel-Dieu, Appel d’offre. Paris, France.
-
Moumaris M (2005) Mise en évidence d’une nouvelle molécule pour suivre l’apoptose. Procédures de dépôt d’un brevet. Paris, France.
-
Moumaris M (2003) La recherche biomédicale, la loi de bioéthique relative au don et à l’utilisation des éléments et produits du corps humain. Paris, France.
-
Moumaris M, Abuaf N (2002) Use of labeled dextran for in-vitro assessment of increased cell permeability, cell death and apoptosis. Brevet n°00/09235, National Institute of Industrial Property (INPI – France) 2811682: A3.
-
Moumaris M, Benoliel S, Rouquette AM, Rajoely B, Abuaf N (2000) Phospholipid binding proteins on the plasma membrane of lymphocytes. J Autoimmun 15: A33.
-
Moumaris M, Ignoti S, Benoliel S, Oghina G, Rajoely B, et al. (1999) Characterization of B-cells adhering to the lamellar phospholipids. French Congress of Antiphospholipid Antibody Paris, France.
-
Moumaris M (1996) Membranes érythrocytaires dans le paludisme: modèle d’étude: Souris- Plasmodium berghei anka. Paris, France.
-
Moumaris M, Sestier C, Miltgen F, Halbreitch A, Gentilini M, et al. (1995) Effect of Fatty Acid Treatment in Cerebral Malaria-Susceptible and Nonsusceptible Strains of Mice. The Journal of Parasitology 81 (6): 997-999.
-
Sabolovic D, Moumaris M, Miltgen F, Sestier C, Halbreich A (1995) Characterisation of subpopulation of red blood cells as a preferentiel target for malaria invasion. French Congress of Electrophoresis, Cell Electrophoresis, Pastor Institute, Paris, France.
-
Sabolovic D, Moumaris M, Miltgen F, Sestier C, Halbreich A (1995) A subpopulation of red blood cells induced by bleeding or mosquito sucking. Chinese National Congress of Medical Biophysics Shanghai, China.
-
Moumaris M (1992) Maladie de Lyme: Étude sérologique. Orleans, France.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework