Revisiting Hunterella nodulosa Mackiewicz and McCrae, 1962 (Cestoda: Caryophyllidae) from Catostomus commersoni (Lacépède) with a Special Treatment of its Ecological Diversity in Wisconsin Waters
Specimens of Hunterella nodulosa Mackiewicz and McCrae, 1962 (Caryophyllidae) were first collected in SE Wisconsin from the white sucker Catostomus commersoni (Lacépède) in Tichigan Lake and the Root and Pike rivers. We update our first and only 1986 account of these Wisconsin populations from Tichigan Lake, with new observations and measurements, and compare them with those in the original and other descriptions. The morphology of H. nodulosa is revisited using light microscopy revealing new features. Emphasis is placed on the morphological variability of its unusual scolex and reproductive structures with variable ovarian shapes using many microscopic images depicting real-life forms. Measurements of 10 specimens and the re-examination of 12, 85, and 33 more specimens from Tichigan Lake, Root River, and Pike River for the first time, respectively provided an opportunity to review the taxonomy and description of the species, distinguish morphometric locality differences, and correct and update other statements and concepts by other observers. The disconnected distribution of worms from the same host species in separate waters within Wisconsin and in various states may be related to the disjunct post-glacial distribution of their primary hosts.
Omar M Amin* and Nataliya Yu Rubtsova
Introduction
Mackiewicz [1] reviewed the morphology, anatomy, development, zoogeography, and systematics of the Caryophyllidea van Beneden, 1893 (Cestoidea) and indicated the existence of 89 species and 37 genera in these monozoic cestodes. Mackiewicz [2] provided perspectives on the evolution, biology, and zoogeography of 128 species in 45 genera of caryophyllids from the Nearctic, Neotropical, Palearctic, Ethiopian, Oriental, and Australian regions. Mackiewicz [3] later recognized about 150 species in 41 genera worldwide (except for the Neotropical Region). Of
these, 14 genera and 90 species belonging to 3 families were described from the Indo-Malayan region from Bagridae, Heteropneustidae, Schilbeidae and Siluridae, Cypriniformes and cobitid fishes [3]. Ash, et al. [4, 5] invalidated a large number of inadequately described species from clariid and catfishes in India and neighboring countries. Most recently, however, Scholz, et al. [6] and Scholz and Kuchta [7] recognized only 117 species and 46 genera in order Caryophyllidea Carus, 1863 parasitizing teleost fish (Cypriniformes, Siluriformes, and some catfish). Chubb JC [8] reviewed the seasonal ecology of the Caryophyllidea in the various climate zones of the world.
Amin OM [9, 10, 11, 12] recognized 15 species in 8 families of caryophyllid cestodes mostly from suckers (Catostomidae) in lake and river systems in Arizona and Wisconsin. These included Hunterella nodulosa Mackiewicz and McCrae, 1962 from Catostomus commersoni (Lacépède) in one lake and two river systems in Wisconsin. Hunterella nodulosa has had a turbulent nomenclature and taxonomic history since its original description as Monobothrium terebrans Linton, 1893 from Catostomus adrens (Jordan and Gilbert) in Wyoming. Nybelin [13] recognized Linton’s species in Monobothrium and Woodland [14] and transferred it to the genus Caryophylleaus Gmelin, 1790 as amended. Hunter [15] provided a detailed description of Linton’s [16] species as Caryophyllaeus terebrans (Linton, 1893) Char. emend, and coined his redescription as a “revised description … based on the original paper of Linton [16] upon some but not all of his original slides and some of the same species collected from Ictiobus bubalus (Rafinesque).” Hunter [15] (p. 30) considered this cestode as “the first cestodarian parasite to be recorded from North America.” Hunter [15] may have not picked up only Linton’s slides of Caryophyllaeus terebrans. In their description of H. nodulosa, Mackiewicz, et al. [17] (p.804) noted that the slides of Linton’s Caryophyllaeus terebrans paratypes included two different genera: H. nodulosa and “another worm corresponding to the “adult specimen” of Linton and Caryophyllaeus terebrans (Linton) as understood by Hunter [15]”. The primary difference between worms in these 2 genera was cited to be in the reproductive structures as depicted in figures 15 and 13. These authors Mackiewicz, et al. [17] ventured that this “other worm” could be Glaridacris catostomi Cooper, 1920 which often appeared in cross infections of C. commersoni with H. nodulosa. At this point in time, one wonders about the justification for creating a new taxon for C. terebrans since the generic diagnosis of Hunterella according to Mackiewicz and McCrae [17] perfectly matches the relevant elements of Linton’s [16] description and the follow-up by Hunter [15] instead of simply and selectively redescribing or revising Linton description based on slide USNM Helminthol. Coll. No.
51074 & 35.51a of the worm species Mackiewicz, et al. [17] considered H. nodulosa, as Hunter [15] did.
One will find the description of Linton [16] based on the above two slides to have embodied every single detail of the diagnosis of H. nodulosa in Mackiewicz and McCrae [17] quoted verbatim following. “Caryophyllaeidae without suckers, loculi, or other organs of attachment on scolex. Cirrus opens separately from female gonopore. Ovary H-shaped. Coils of uterus not extending anteriorly beyond cirrus pouch. Longitudinal muscles diffuse, not forming definite layers except in the posterior regions of the body. Inner longitudinal muscles loosely arranged in a broad band around the medullary region. Vitellaria embedded within this medullary region, generally surrounded on all sides by fibers of the inner longitudinal muscles. Post-ovarian vitellaria and external seminal vesicle present.” Based on the arguments of Mackiewicz and McCrae [17], other authors like Barčák, et al. [18] mistakenly listed the current taxonomic status or valid name of “Caryophyllaeus terebrans (Linton 1893) Woodland, 1923 as Glaridacris terebrans (Linton, 1893) Mackiewicz, 1974 (which was) “originally described in Monobothrium but Woodland [14] synonymized this genus with Caryophyllaeus.” What Barčák, et al. [18] were actually referring to is “the other worm” (slide no. 32.51b) and not slide no. 32.51a which contained Linton’s H. nodulosa as Mackiewicz [19] later on acknowledged. Mackiewicz [19] may have resolved this dilemma by declaring that “there are no authentic records of Caryophyllaeus Gemlin (with typical flabelliform scolex) in the Nearctic.” In Wisconsin, Amin [12, 20] gave an account of H. nodulosa from C. commersoni in the Pike and Root rivers and Tichigan Lake, SE Wisconsin, and made a brief distinction between it and the description of Mackiewicz, et al. [17].
Materials, Collections, and Methods
Collections
In SE Wisconsin, 22 specimens of H. nodulosa were collected from 7 of 105 individuals of C. commersoni in Tichigan Lake, SE Wisconsin between 1976 and 1984 [12]. Tichigan Lake is a relatively large eutrophic impoundment on the Fox River which is a tributary of the Mississippi River in Racine County (42°48′20″N,88°13′00″W). A total of 85 and 47 specimens of H. nodulosa were collected from C. commersoni in the Root River, Racine, and Milwaukee counties (42°44’1” N-87°46’42” W), and the Pike River, Racine, and Kenosha counties (42°38′40″N,87°51′09″W), respectively in SE Wisconsin in 1973 and 1974 (Table 1).
The 2 rivers are not connected to each other or to Tichigan Lake and they both flow eastwards into Lake Michigan independently. “One juvenile and one gravid Biacetabulum biloculoides Mackiewicz and McCrae, 1965 were recovered from the stomach of a 36-cm-long female C. commersoni collected from Tichigan Lake which also harbored one gravid H. nodulosa, 3-juvenile Glaridacris catostomi Cooper, 1920 and 5 juvenile Biacetabulum sp. in the stomach and post- gastric region [12]. White suckers from the Root River were moderately infected with B. biloculoides in the fall of 1974 and some were concurrently infected with specimens of H. nodulosa and Biacetabulum macrocephalum McCrae, 1962 (miscellaneous).
Methods
After capture and transfer to the laboratory in coolers on ice, the fish were dissected within a few hours after capture. Upon recovery, worms were placed at once in warm 70% ethanol then processed and stained with Semichon’s carmine, destained in 4% hydrochloric acid in 70% ethanol, and dehydrated in ascending concentrations of ethanol. Worms were ultimately cleared in oil of wintergreen before the whole mounting in Canada balsam. Measurements are in mm or micrometers as noted in Table 1; the range is followed by the mean values between parentheses. Width measurements represent maximum width. Microscope images were created using 10X and 40X objective lenses of a BH2 light Olympus microscope (Olympus Optical Co., Osachi- shibamiya, Okaya, Nagano, Japan) attached to an AmScope 1000 video camera (United Scope LLC, DBA AmScope, Irvine, California), linked to an ASUS lab top equipped with HDMI high-definition multimedia interface system (Taiwan- USA, Fremont, California). Images from the microscope were transferred from the lab top to a USB and stored for subsequent processing on a computer. We found microscope images to be considerably more informative than schematic line drawings as they depict the natural appearance of anatomical structures.
Results
Amin [12] gave morphological notes of H. nodulosa from the Wisconsin population collected from the white sucker C. commersoni captured in Tichigan Lake, SE Wisconsin between 1976 and 1984. Amin [12] made initial taxonomic comparisons with specimens from other collections in Colorado and New York [17]. We have restudied specimens in our personal collection from Tichigan Lake as well as many other specimens from the Root and Pike Rivers in SE Wisconsin. Measurements of about 10 specimens were made in 1967-1968 [12] and new measurements and observations of additional specimens from the other collections in the Root and Pike Rivers were recently made and added to the earlier measurements creating combined measurements included in Table 1.
| State | Colorado & New York | Wisconsin | ||
|---|---|---|---|---|
| Locality | 4 sites in Colorado (CO) & 11 sites in New York (NY) | Tichigan Lake, Racine Co. | Root River, Racine & Milwaukee counties | Pike River, Racine & Kenosha counties |
| Authority | Mackiewicz & McCrae (1962) | Amin (1986) & this paper | This paper | This paper |
| Specimens measured | 30-50 (900 observed) | 10 + 12 | 85 | 47 |
| Collection date (N) | March, 1956-Nov., 59 (CO) July, 1955-June, 1958 (NY) | Autumn, 1977 (10); Spring, 1984 (12) | Autumn, 1974 (85) | Autumn (22), Spring (4), 1973 July, 1974 (21) |
| Body form | Elongate, broader anteriorly, bluntly pointed at both ends or variable | Elongate, bluntly pointed at both ends, broader anteriorly | Elongate, often broader & bluntly pointed anteriorly, often bilobed posteriorly | Short & stubby, often drop- shaped |
| Body length (mm) | 2.2-8.0 (from 5.0 to 15.0* mm long in live worms) | 2.6-6.0 (3.92) | 2.25-5.50 (3.53) | 1.22-3.95 (2.27) |
| Width at gonopore (mm) | 0.4-1.2 | 0.52-1.25 (0.85) | 0.67-1.37 (1.01) | 0.42-1.12 (0.70) |
| Scolex | Simple, unspecialized conical enlargement lacking specialized musculature, loculi or acetabulae | |||
| Length of scolex (mm) | --- | 0.35-1.08 (0.61) | 0.25-0.75 (0.50) | 0.22-0.62 (0.45) |
| Width of Scolex (mm) | 0.35-1.6 | 0.54-1.12 (0.74) | 0.50-1.12 (0.87) | 0.45-0.75 (0.65) |
| Vitellaria | Irregular, medullary, circular around testes, more crowded posteriorly | Somewhat irregular- ovoid-elongate laterally. Primarily lateral but merge medially, more crowded posteriorly | ||
| Pre- & post-ovarian vitellaria | “May be continuous” | Often continuous | ||
| Vitellaria length (µm) | --- | 62-198 (103) | 104-187 (149) | 104-156 (117) |
| Vitellaria diameter (µm) | 51-153 | 56-135 (95) | 73-135 (93) | 72-114 (79) |
| Testes (T) | Irregularly globose, extend anterior to ant. vitellaria | Multi-layered in 4 irregular rows. Extend anteriorly past anterior vitellaria and posteriorly before posterior end of pre-ovarian vitellaria | ||
| Testes (T) number | 95-175 (95-117 in N.Y.) | 59-120 (95) | 79-126 (93) | 60-119 (84) |
| Testis diameter (µm) | 50-100 | 56-208 (130) | 104-198 (160) | 94-166 (117) |
| First testis to ant. tip (mm) | --- | 0.36-0.97 (0.70) | 0.38-1.00 (0.66) | 0.25-0.67 (0.47) |
| % of worm length | --- | 10-19% (16%) | 11-19% (16%) | 13-23% (17%) |
| First pre-ovarian vitellarium to ant. tip (mm) | --- | 0.40-1.20 (0.91) | 0.52-1.50 (0.94) | 0.41-0.95 (0.69) |
| % of worm length | --- | 14-30% (20%) | 15-29% (23%) | 20-38% (25%) |
| Post-gonopore distance (mm) | --- | 0.80-1.45 (1.08) | 0.53-1.25 (0.92) | 0.52-0.78 (0.63) |
| % of worm length | --- | 19-33% (28%) | 19-27% (23%) | 20-27% (24%) |
| Cirrus sac | Ovoid | Relatively large, spheroid with concentric rings | ||
| Cirrus sac diameter (µm) | 114-205 | 98-266 (188) | 114-250 (215) | 124-229 (170) |
| Extruded cirrus length (µm) | 130 (Fig. 7) | 200 | --- | 124-166 (152) |
| External seminal vesicle | Large, reniform | Large, ovoid | ||
| Ext. seminal vesicle L. (µm) | 100-165 | 177-322 (250) | 104-208 (146) | 104-125 (114) |
| Ext. seminal vesicle W. (µm) | --- | 135-208 (171) | 83-156 (114) | 62-104 (83) |
| Cirrus & uterovaginal pores | Separate | Independent | ||
| Post-ovarian vitellaria | --- | Clustered, often in bird wing-form | ||
| No. of post-ovarian vitellaria | 16-19 (Figs. 3, 4) In all specimens | 14-40 (24) | 22-38 (27) | 15-26 (19) |
| Ovary shape | Compact, non-follicular, H-dumbbell-shaped with rounded short wings | Dumbbell-shaped. H-shaped with short wings, non-follicular. | ||
| Ovarian arms’ length (µm) | 110-480 | 168-416 (299) | 208-416 (398) | 177-353 (239) |
| Ovarian arms’ width (µm) | --- | 166-333 (247) | 177-343 (230) | 135-250 (180) |
| Commissure diameter (µm) | 40-160 | 135-229 (171) | 104 (N=1) | 94-135 (114) |
| Shape of posterior end | --- | Bluntly pointed or reniform or bi-lobed | ||
| Eggs | Operculate, smooth shell, with 4-11 yolk cells | Operculate, smooth, ovoid, with up to 10 yolk cells | ||
| Egg length (µm) | 50-61 in CO, 60-70 in NY | 48-63 (57) | 47-75 (58) | 42-57 (52) |
| Egg diameter (µm) | 30-37 in CO, 37-42 in NY | 32-50(38) | 31-45 (39) | 33-43 (38) |
| Eggs measured | 150 (50 in CO, 100 in NY) | 45 | 75 | 38 |
Table 1: Comparative morphometrics of adult Hunterella nodulosa from Catostomus commersoni in Tichigan Lake, Pike and Root rivers
Our Wisconsin specimens were smaller than those from Colorado and New York [17] and had fewer testes and larger cirrus sacs (Table 1). Compared to the Wisconsin populations from Tichigan Lake and the Root River, those from the Pike River assumed frequent odd shapes and were characteristically and distinctly smaller in body size and corresponding sizes of scolex, vitellaria, testes, post- gonopore distance, external seminal vesicle, cirrus sac, ovarian arms, commissure, and eggs (Table 1).
We provide below an elaborate revised description based on our Wisconsin material and give detailed comparative morphometrical data (Table 1) high-lightening extreme variations from the other description by Mackiewicz, et al. [17]. Relevant concepts of Hunter [15] are considered. Our description traces anatomical structures from the scolex anteriorly to the posterior end, not essentially by organ system.
Brief Morphological Description of Adults of H. nodulosa from Wisconsin
With characters of the genus Hunterella as described in Mackiewicz, et al. [17] and as originally included in Linton’s Monobothrium terebrans Linton, 1893, in part [16], later synonymized with Caryophyllaeus terebrans (Linton 1893) Woodland, 1923. Measurements and counts are in Table 1. Juveniles, shaped like adults (Fig. 1 from Root River), with underdeveloped testes and later developing vitellaria, and immature ovary and uterus. Body short, oblong, widest anteriorly or at gonopore especially in non-cylindrical short forms (Figs. 2, 3 from Pike River) and tapering at both ends. Fixed specimens may assume variable forms including drop- shaped ones (Figs. 2, 3). Not dorso-ventrally flattened, with unspecialized blunt cone-shaped scolex and no neck (Figs. 4-6 from Root River and Tichigan Lake). Thin cuticular and cortical layers cover and defuse inner longitudinal muscle layer encircling medulla; outer longitudinal muscles may be absent. Adults with 59-126 spheroidal testes beginning anterior to anterior vitelline glands (Figs. 4-6) and emerging sooner in younger stages, usually irregularly distributed in 4 dorsal-ventral medial rows occasionally appearing in partially uneven or random rows (Figs. 7,8 from Tichigan Lake and Root River). Mature adults with testicular field extending from 16-17% of anterior worm tip to just anterior to cirrus sac. Testes larger posteriorly where they may be obscured by posterior vitellaria. Paired osmoregulatory canals in testicular field. Cirrus sac large, oblong- round, near ventral surface at tip of anterior ovarian arms (Figs. 9-12 from Tichigan lake and Root River). External seminal vesicle large, ovoid, partially overlaying cirrus sac (Fig. 9, arrow). Pre-ovarian vitelline glands large, only slightly smaller than testes, ovoid-elongate, multi-layered, primarily in 2 lateral fields (Figs. 21,22), often extending and merging medially, especially when more crowded posteriorly and encircling obscured posterior testes; from 20-25% of anterior worm tip to anterior end of ovarian arms). Male and female reproductive systems open independently with post gonopore distance of 23-28% of worm length. Vas deference detectable about middle of testicular field ventrally. Distinctly dumbbell-shaped H-shaped ovary with short wings and straight commissures (Figs. 13-15 from Root River and Tichigan Lake & Fig. 17 from Pike River) except when showing unusual variations such as V-shaped commissure or commissure expanding on one side (Figs. 16,18 from Pike River). Ovarian commissure may fan at juncture with ovarian arms on one or both sides (Fig. 18). Vagina with seminal receptacle and running posteriorly to anterior aspect of ovarian commissure. Uterine coils pre- and post- ovarian but not extending anterior to cirrus sac. Post-ovarian vitellaria always present, especially large, often continuous with pre-ovarian vitellaria, with 14-40 glands clustered or in bird-wing--form (Figs. 13,19-22 from Root River & no. 17 from Pike River). Posterior end bluntly conical, rounded (Figs. 1, 15) or rump-like with slight or deep terminal middle invagination revealing excretory bladder (Figs. 20-23). Eggs small, ovoid, thin-shelled, operculate, with up to 10 yolk cells (Fig. 24).
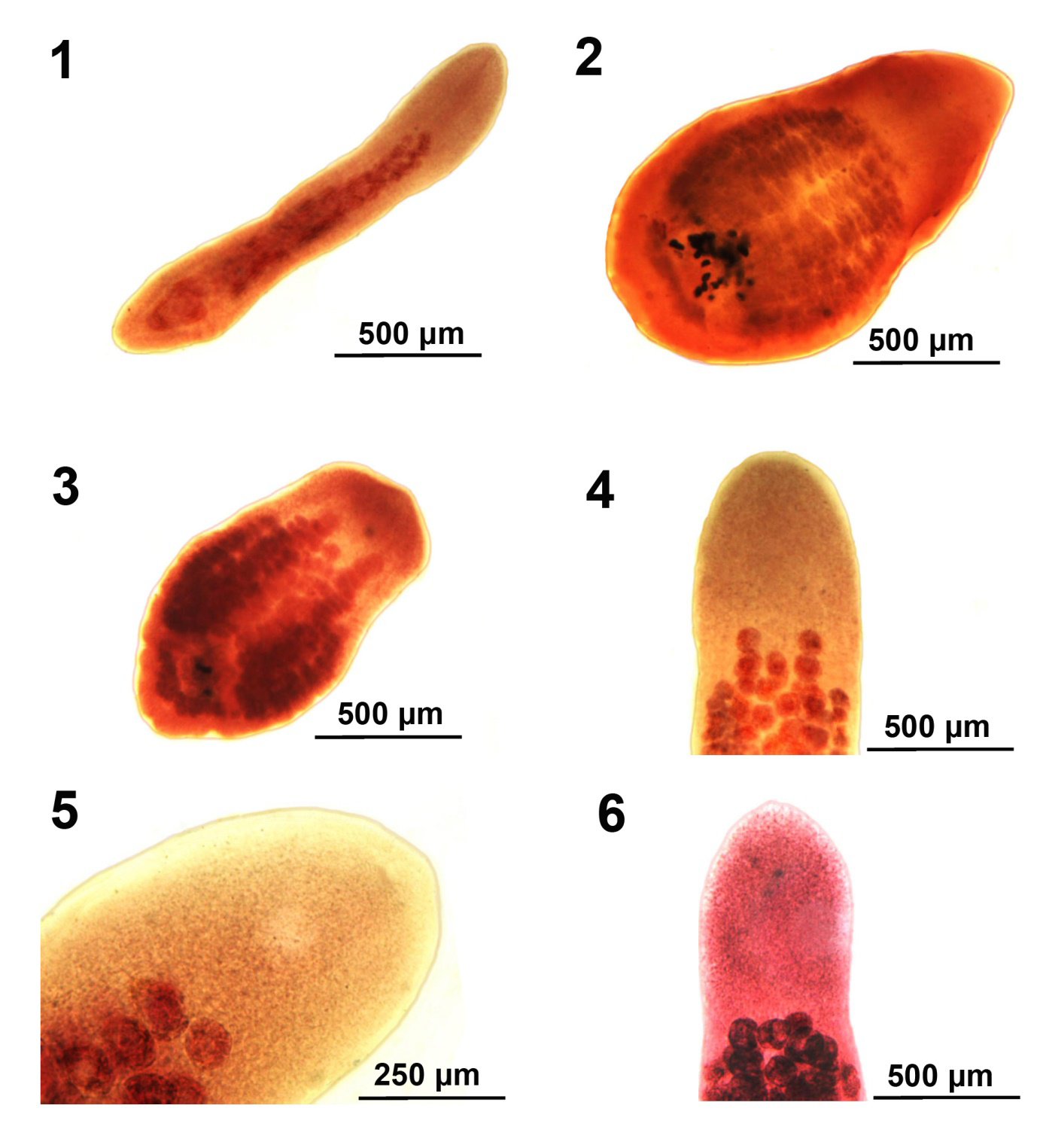
Figures 1-6: Body forms of specimens of Hunterella nodulosa from Catostomus commersoni in Tichigan Lake (T), the Pike River (P), and Root River (R), SE Wisconsin. 1. Usual form of a developing adult (juvenile) (R) showing incipient organs. 2, 3. Common forms of stunted specimens from the Pike River (P). 4-6. The anterior portion of worms (R, T) showing the anterior- most distribution of anterior testes beyond the distribution of anterior-most lateral vitellaria.
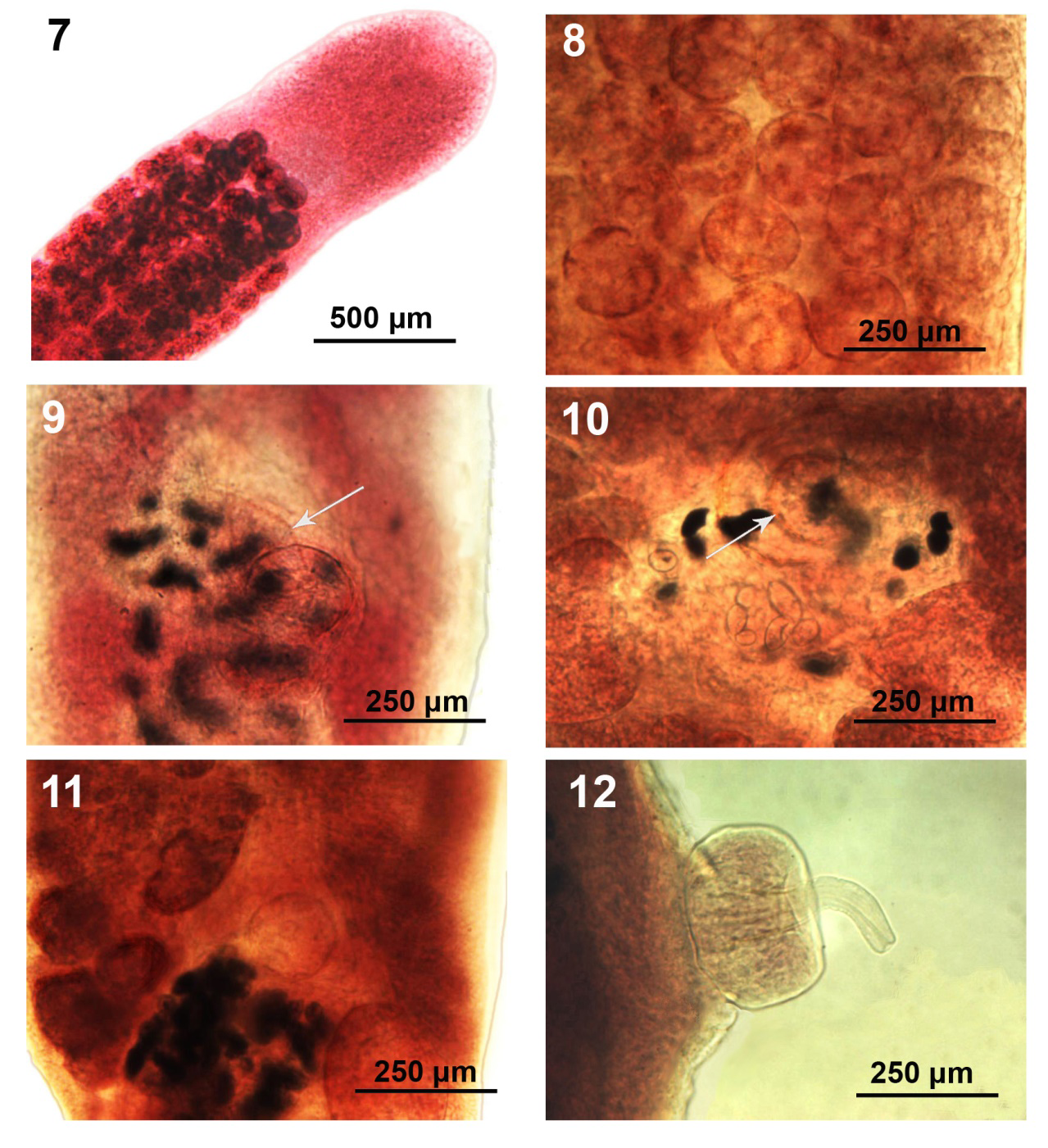
Figures 7-12: Variations in the distribution of testes and the morphology of the cirrus sac of specimens of Hunterella nodulosa from Catostomus commersoni in Tichigan Lake (T), the Pike River (P), and Root River (R), SE Wisconsin. 7 (T), 8 (R). The distribution of testes in longitudinal rows in the anterior medullary region surrounded by primarily lateral columns of vitelline glands. 9 (T)-10, 11 (R). Variations in cirrus sac (ventral). Figs. 9, 10, show the external seminal vesicle (arrows). 12 (T). Lateral view prolapsed cirrus sac with extruded cirrus.
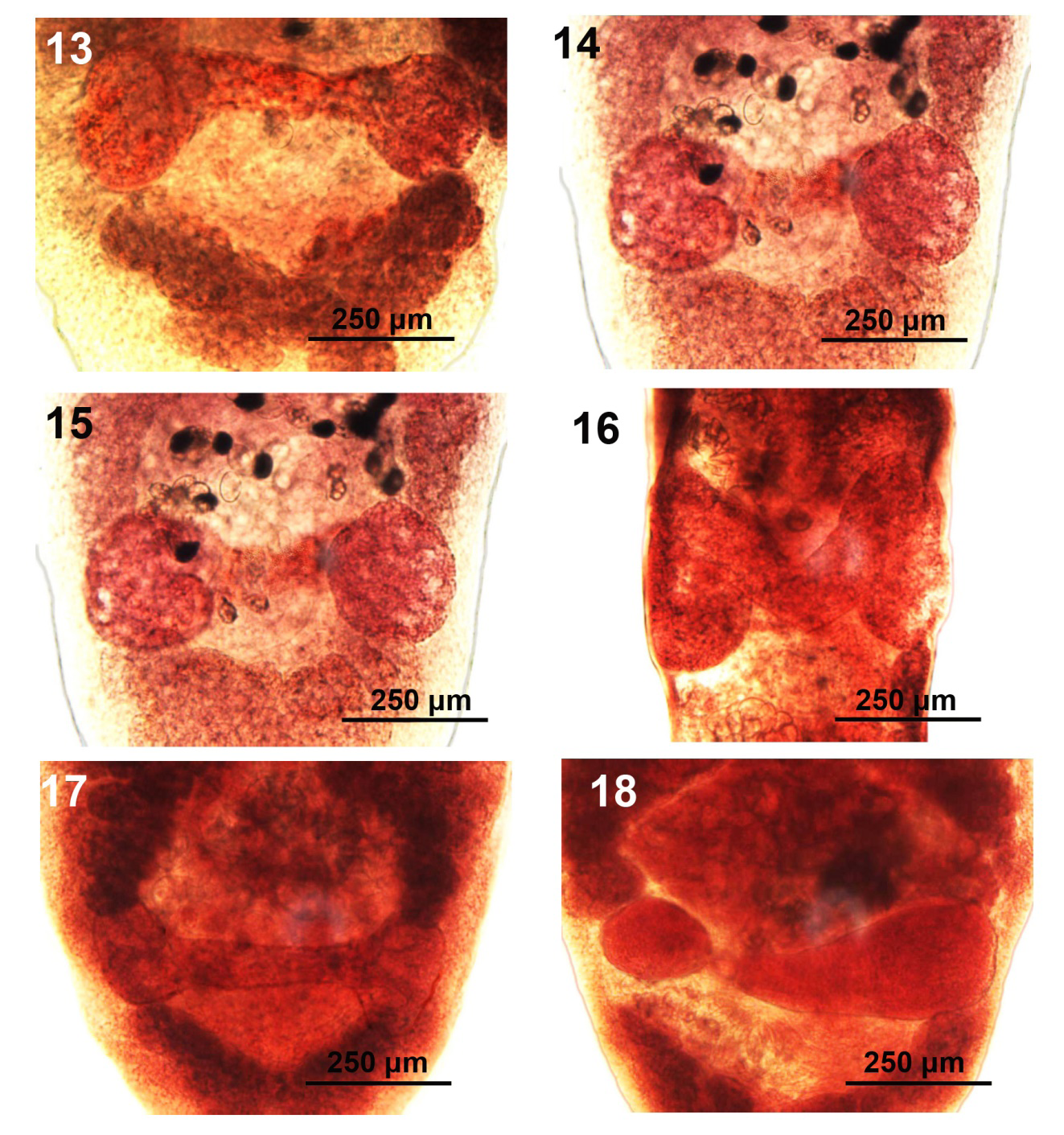
Figures 13-18: Variations in the shape and organization of the ovarian arms, the commissure, and post-ovarian vitellaria of specimens of Hunterella nodulosa from Catostomus commersoni in Tichigan Lake (T), the Pike River (P), and Root River (R), SE Wisconsin. 13 (R)-15 (T), 17 P). Usual variations in rounded ovarian arms, straight commissure, and bird-wing-shaped clustering of post-ovarian vitelline glands. The commissures adjoin the ovaries either anteriorly or at their middle or posterior half. 16 (P), 18 (P). Unusual/ anomalous commissure variations. 16. The commissure assumes a V-shape form. 18. One side of the commissure merges with one ovary as one organ.
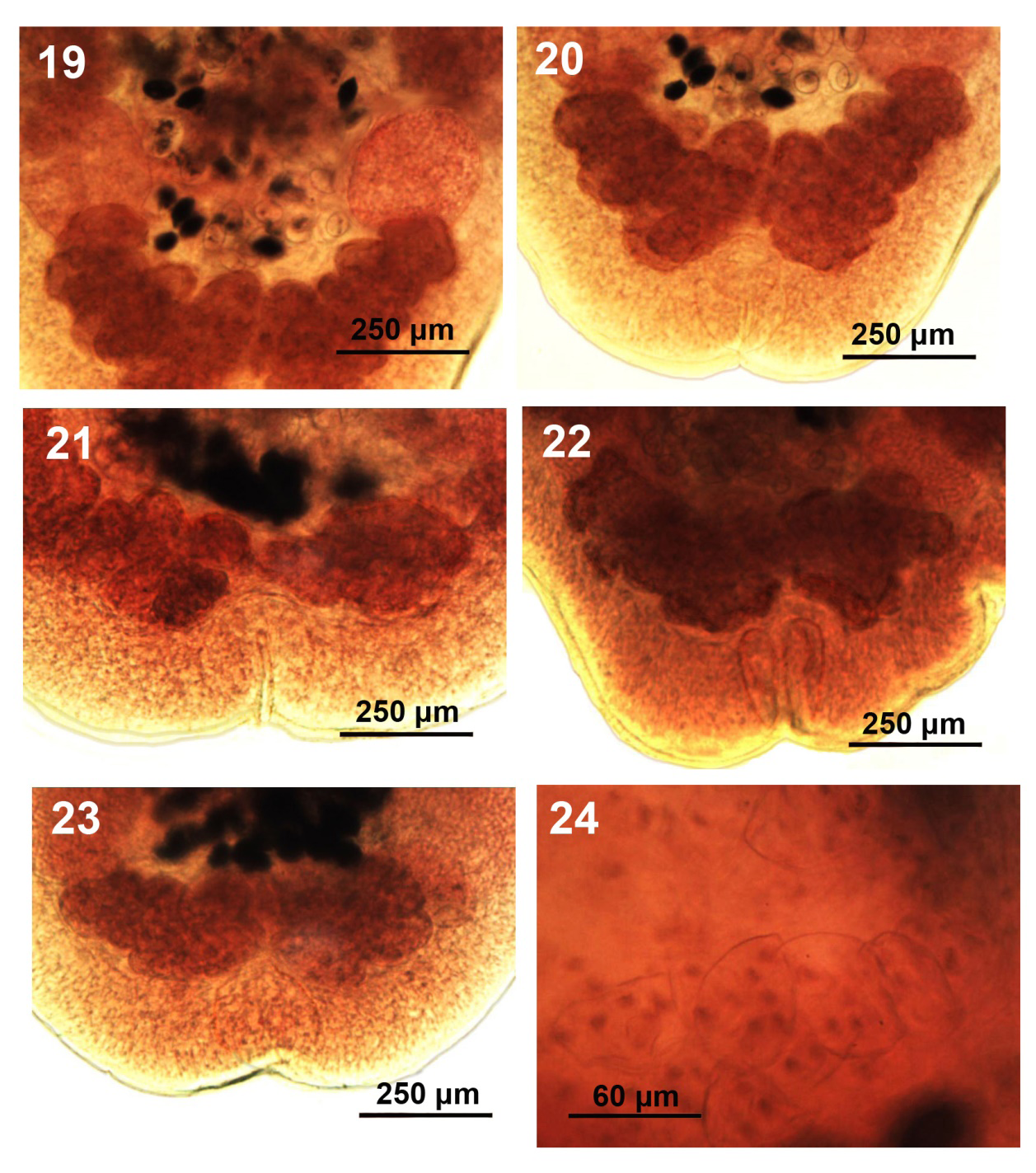
Figures 19-24: Variations in the shape and organization of the post-ovarian vitellaria, posterior body morphology, and eggs. All from Root River specimens of Hunterella nodulosa from Catostomus commersoni, SE Wisconsin. 19-23. The bird-wing cluster form of the post-ovarian vitellaria and the rump-like shape of the posterior end with invagination outlining the excretory bladder. 24. Eggs with yolk cells.
Taxonomic Summary
Synonymies: Caryophyllaeus terebrans, partim (Linton, 1893), Glaridacris catostomi, partim, (Cooper, 1920), Hunterella nodulosa, nomen nudum McCrae, 1961 as per Mackiewicz and McCrae [17]. Type host: Utah sucker Catostomus ardens Jordan and Gilbert of Caryophyllaeus terebrans, partim, in Linton [16].
Primary host: White sucker Catostomus commersoni (Lacépède). Other host: Largescale sucker Catostomus macrocheilus Girard in British Columbia (Canada) and Washington. Type locality: Heart Lake, Snake River drainage System, Yellow Stone National Park, Wyoming; in Linton [16]. Type host and type locality: Not listed in Mackiewicz and McCrae [17].
Other localities In Catostomus commersoni from the following states as per Mackiewicz and McCrae [17]: Colorado, Connecticut, Delaware, Michigan, Minnesota, New York, North Carolina, Ohio, Pennsylvania, South Dakota, Tennessee, Virginia, West Virginia, Wisconsin including the present investigation, and Wyoming. Deposited Wisconsin specimens: US National Museum Helminth. Coll. Nos. 78660, 78811.
Remarks
The taxonomic history of the evolution of the concept of H. nodulosa is replete with conflicting ideas and designations. The material described by Linton [16] as Monobothrium terebrans included two species one of which is currently known as H. nodulosa as recognized by Mackiewicz and McCrae [17] and Mackiewicz [19] based on Linton’s slide no, 32.51a. See the Introduction of this paper for nomenclature history and changes. Hunterella nodulosa is clearly a widely distributed species across North America albeit in a limited number of host species with C. commersoni being the most common. It is morphologically diverse with extremes of life forms and sizes that vary with geographical distribution (Table 1). This morphological diversity was observed among different populations collected from the same fish species in adjacent streams and bodies of water as noted in the present investigation from C. commersoni in Tichigan Lake, Root and Pike rivers in SE Wisconsin (Table 1). Specimens of the Pike River population were the smallest in body and other major anatomical structures as well as mostly assuming distinctly variable forms especially a drop-shaped form not usually seen in the other Wisconsin populations. Specimens from the Root River and Tichigan Lake were more comparable to those described by Mackiewicz, et al. [17] from Colorado and New York. Lumping all our SE Wisconsin together, we still had specimens considerably smaller than those described from New York and Colorado [17] (Table 1). Irrespective of the smaller size of the Pike River specimens compared to those of the Root River or Tichigan Lake, the ratios and proportions of body parts to body length were similar. For instance, among these populations, the distance between the first testis to the anterior tip divided by body length was 13- 23 %, 11-19%, and 10-19% in the same order. Corresponding ratios for the first pre-ovarian vitellaria to anterior tip divided by body length were 20-38%, 15-and 29%, 14-30%, and for post-gonopore distance divided by body length was 20-27%, 19-27%, and 19-33%.
For an additional measure of morphological diversity even only among the Wisconsin, New York, and Colorado populations, note the intra-populational differences noted within the 3 Wisconsin populations with bolded numbers representing extreme differences and compare with those noted for the New York and Colorado populations. Our figures will underline extremes of morphological forms not previously described and shown by line drawings anywhere.
Discussion
While specimens of H. nodulosa from SE Wisconsin were collectively markedly smaller than those studied in New York and Colorado (Table 1), the growth of those from the Pike River appear to have been stunted compared to those from the adjacent Root River even though the smallest Pike River specimens (1.22 mm long) were sexually mature with eggs. We attribute such morphometric differences to physiographic factors related to water quality of the streams. Differences in the levels of non-fecal organic pollutants in these two streams and the higher flow rates in the larger Root River as calculated by the Southeastern Wisconsin Regional Planning Commission [21] may be related. Additionally, the Pike River specimens manifested the widest diversity of body forms exclusively including drop-shaped specimens (Figs. 2, 3).
Hunter’s [15] revision of Caryophyllaeus terebrans based on Linton’s [16] Monobothrium terebrans after Woodland’s [14] synonymization of the two genera was detailed but included elements of 2 species, one of which was later described by Mackiewicz and McCrae [17] as H. nodulosa. Hunter [15] recognized the possibility that he may have been dealing with two species when confronted with the other longer forms later recognized as a species of Glaridacris Cooper, 1920 by stating that “either Linton had representatives of two species or that one matured too early before the male reproductive system had completed its development, the reverse of the normal condition” (p. 33). Hunter [15] then dismissed the first possibility by immediately stating that “The latter explanation has been accepted by the author” (p. 34). Amin [12] have often observed that specimens of H. nodulosa “cohabited with G. catostomi in anterior intestinal locations of C. commersoni” [12] (Figs. 1,2, p. 51, 52).
Some of the elements of Hunter’s [15] treatment of Linton’s [16] account attributable to H. nodulosa as we now recognize it includes being embedded in intestinal wall in pits, reproductive systems opening apart, scolex bluntly cone-shaped with no suckers or loculi, vitellaria being “marginal” and posterior, and H-shaped globular ovary. Some of his [15] elements not attributable to H. nodulosa include adults as long as 30 mm long, distinct neck, inner and outer longitudinal muscles present and equally prominent, and absence of seminal receptacle.
The description of H. nodulosa by Mackiewicz and McCrae [17] was thorough almost leaving no detail uncovered. The description of that new genus was almost identical to that of Linton’s, partim [16] and the distributional records and morphological treatment were meticulous. Few statements, however, produced the following questions. For example, unlike their Fig. 3 (p. 800) of a mature worm correctly showing anterior-most testes to be more anterior than anterior-most vitellaria, Fig. 6 of a “larva” (actually a young adult with an incipient reproductive system with identifiable male and female gonopores, testes, and vitellaria) depicting anterior-most testes well posterior to anterior-most vitellaria [17]. In our specimens developing juveniles have a complete set of testes before vitellaria began to emerge. They [17] described the vitellaria as “arranged circularly around testes” (p. 799). Their cross-section (Fig. 13, p. 802) does depict this arrangement which appears to be true but only at the posterior body as Fig. 3 of a mature worm suggests. However, for most of the length of the body, the pre-ovarian vitellaria appears to be primarily lateral with an only occasional incursion into the testicular medullary field (our figures 7, 8); microscope images don’t lie. Their fixed specimens (Fig. 9) assumed a wide variety of atypical forms [17] the last of which was similar to our drop-shaped fixed specimens from the Pike River (Figs. 2,3). A line drawing of one such specimen was incorrectly placed with others of Biacetabulum biloculoides Mackiewicz and McCrae, 1965 collected from C. commersoni in the Root River [11]. These authors [17] did not explain why the number of testes or the size of eggs were discretely different between specimens from New York and those from Colorado. The disconnected distribution of worms from the same host species in separate waters within Wisconsin and in various states may be related to the disjunct post-glacial distribution of their primary hosts.
Acknowledgments
This project was supported by an Institutional Grant from the Parasitology Center, Inc. (PCI), Scottsdale, Arizona.
Declarations
Compliance with ethical standards
- Conflict of interest: The authors declare no conflicts of interest or competing interests.
- Ethical approval: The authors declare that they have observed all applicable ethical standards.
- Availability of data: All presented and related data are available by contacting the senior author.
References
-
Mackiewicz JS (1972) Caryophyllidea (Cestoidea): a review. Exper Parasitol 31(3): 417-512.
-
Mackiewicz JS (1982) Caryophyllidea (Cestoidea): Perspect Parasitol 84: 397-417.
-
Mackiewicz JS (1994) Order Caryophyllidea van Beneden in Carus, 1863. In: Khalil LF, Jones A, et al. (Eds.), Keys to the cestode parasites of vertebrates. CAB Intern Wallingford UK, pp: 21-43.
-
Ash A, Scholz T, Oros M, Kar PK (2011a) Tapeworms (Cestoda: Caryophyllidea), parasites of _Clarias batrachus_ (Pisces: Siluriformes) in the Indomalayan region. J Parasitol 97(3): 435-459.
-
Ash A, Scholz T, Oros M, Levron C, Kar PK (2011b) Cestodes (Caryophyllidea) of the stinging catfish _Heteropneustes fossilis_ (Siluriformes: Heteropneustidae) from Asia. J Parasitol 97(5): 899-907.
-
Scholz T, Kuchta R, Oros M (2021) Tapeworms as pathogens of fish: A review. J Fish Dis 44(12): 1-18.
-
Scholz T, Kuchta R (2017) A digest of fish tapeworms. Vie et milieu - Life and environment 67(2): 43-58.
-
Chubb JC (1982) Seasonal occurrence of helminths in freshwater fishes Part IV. Adult Cestoda, Nematoda and Acanthocephala. Adv parasitol 20: 1-292.
-
Amin OM (1968) Helminth fauna of Suckers (Catostomidae) of the Gila River System, Arizona. PhD Thes Ariz St Univ 202 pp (No 68-1644) Univ Microfilms. Ann Arbor, Michigan; Dissert Abstr 28: 3521.
-
Amin OM (1969) Helminth fauna of suckers (Catostomidae) of the Gila River System, Arizona. II. Five parasites from _Catostomus_ spp. Am Midland Nat 82: 429- 443.
-
Amin OM (1974) Intestinal helminths of the white sucker, _Catostomus commersoni_ (Lacépède), in SE Wisconsin. Proc Helminthol Soc Wash 41: 81-88.
-
Amin OM (1986) Caryophyllaiedae (Cestoda) from lake fishes in Wisconsin with a description of _Isoglaridacris multivitellaria_ sp. n. from _Erimyzon sucetta_ (Catostomidae). Pro Helminthol Soc Wash 53:48-58.
-
Nybelin O (1922) Anatomisch-systematische Studien über Pseudophyllideen. Götenburgs Kungl. Vetensk. Vitterh. Samh. Handl. Fjärde Puljden 26: 1-228.
-
Woodland WNF (1923) On some remarkable new forms of Caryophyllaeidae from the Anglo-Egyptian Sudan, and a revision of the families of Cestodaria. Quart J Microscop Soc (new ser) 67: 435-447.
-
Hunter III GW (1930) Studies on the Caryophyllaeidae of North America. Ill Biol Monogr 11 (1927), pp: 186.
-
Linton E (1893) On fish Entozoa from Yellowstone National Park. Rep U S Comm Fish Fisheries, 1889 to 1891, pp: 545-564.
-
Mackiewicz JS, McCrae R (1962) _Hunterella nodulosa_ gen. n., sp. n. (Cestoidea: Caryophyllaeidae) from _Catostomus_ _commersoni_ (Lacépède) (Pisces: Catostomidae) in North America. J Parasitol 48: 798-806.
-
Barčák D, Oros M, Hanzelová V, Scholz T (2017) A synoptic review of _Caryophyllaeus_ Gmelin, 1790 (Cestoda: Caryophyllidea), parasites of cyprinid fishes. Folia Parasitol 64: 027.
-
Mackiewicz JS (1974) The genus _Caryophyllaeus_ Gmelin (Cestoidea: Caryophyllidea) in the Nearctic. Proc Helminthol Soc Wash 41: 184-191.
-
Amin OM (1977) Distribution of fish parasites from two southeast Wisconsin streams. Wis Acad Sci Arts Lett 65: 225-230.
-
(1966) Southeastern Wisconsin Regional Planning Commission. Water quality and flow of streams in southeastern Wisconsin. Tech Rep 4: 342.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework