Rethinking Nucleolar Evolution: New Insights into Structure and Function
The nucleolus, earlier considered a ribosome production site, presently recognized as a dynamic center that influences all aspects of cell genome composition, from stress effects to the aging process. The nucleolus generally exists in eukaryotic cells; however, recent research provides evidence of nucleolus-like structures in some prokaryotes, indicating an ancient origin for nucleolus-related functions. This review explores the evolutionary aspect of the nucleolus and evaluates its structural and functional diversity among primary to developed organisms. The proteins in the nucleolus serve a vital role in ribosome biogenesis and also in cell-cycle regulation, DNA repair, and cellular stress, directly linking its activity to health and disease. The nucleolar dysfunction implicated in ribosomal pathology, neural dysfunction confirmed the nucleolus as a critical target for prospective therapeutic approaches. Thus, the nucleolus may be considered as a model for how cellular prospects can evolve to cater to complex biological functions. The presented review about the organization and function of the nucleolus can suggest new insights about basic concepts that conduct cellular organization
Introduction
The nucleolus is a highly regulated substructure lacks membrane and located inside the nucleus covers rRNA gene- groups called as nucleolar organizer regions or NORs (Figure 1) [1].
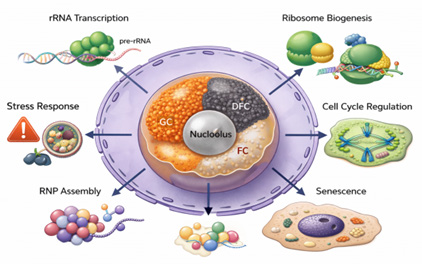
It imparts a major role in ribosomal RNA (rRNA) synthesis, regulate the transcription of rRNA genes, processing nascent rRNAs and assembling ribosomal subunits essential for ribosome biogenesis in eukaryotic cells [2]. The nucleolus is also participated in the cell cycle regulation, sensitive to cellular stress and influencing ageing [3]. It also facilitates the assembly of signal recognition particles for protein synthesis and modifies small nuclear RNAs (snRNAs), which are essential for gene expression [4]. The proteins in the nucleolus involved in cellular metabolism and gene regulation that functioning as a site for the organization and modification of these important cellular components [5, 6].
The changes in structure and function of nucleolus have been associated with various diseases, including cancer, neurodegenerative disorders and viral infections [7]. These links conclude nucleolus as a critical site in cellular regulation and a valuable target for therapeutic pronunciations [8]. A deeper understanding of its diverse functions is essential to uncovering its broader implications in cellular biology and disease.
Results
The Nucleolus Aspects are Essential for its Role in Cellular Processes
The role of nucleolus in ribosome synthesis is well- established, whereas its major impartment in cellular processes remains less clear. This gap needs about deep insights on nucleolar structure and physiology to explore its influence on cellular physiology and in this aspect study of nucleolus evolution is crucial to defined complexity of ribosomal biogenesis and cellular physiology across diverse organisms (Figure 1). The bacteria and archaea ever not possess nucleolus, however, rely upon analogous nucleolus-like structure essentially for the accurate function in their life cycle. In present, it must be clarified about evolutionary adaptation during the life evolution which exhibiting selection pressures and alteration for cellular efficiency and so knowledge about nucleolar processes not only valuable to appreciate cellular functions but also significantly insights on nucleolar dysfunction which is linked with several human diseases as cancer and neurodegenerative disorders [9]. The disease can progressed through nucleolar function through unbalanced ribosome synthesis and alteration in cellular homeostasis. Thus deep insights about nucleolus physiology consists potential to uncover novel diagnostic and therapeutic strategies become possible through hope and inspiration in modern cellular and molecular biology.
There recent progress in imaging, molecular techniques and proteomic approaches have revealed previous unexplored vision and perspective approach into dynamics and functions of nucleolus through high-resolution polar microscopy, genome editing and proteomic analysis of nucleolar components [10]. These approaches have certainly improved nucleolus display in cellular biology. The complexity and mystery of the nucleolus can provide more inspiration to further research in the field of cellular and molecular biology.
Structural Complexity of the Nucleolus: Morphological and Functional Insights
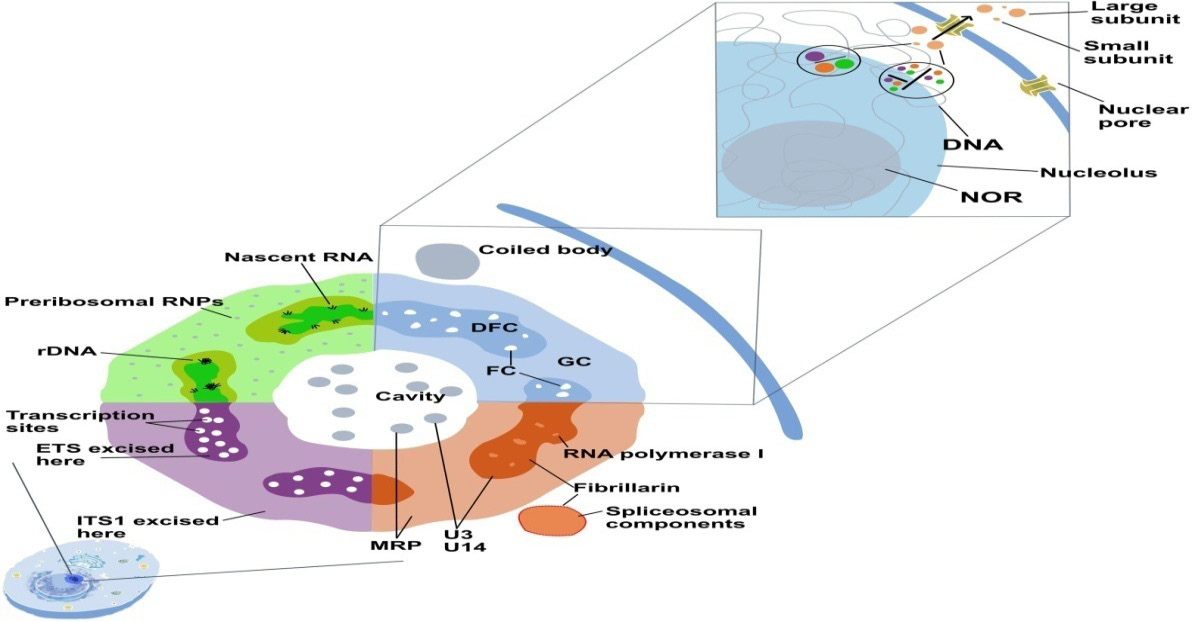
There are key substructures has emerged as central to nucleolar function through transmission electron microscope (TEM). The fibrillar centre (FC) is critical area for ribosomal DNA (rDNA) storage. The transcription of nascent rDNA initially drives at the junction of FC and covering dense fibrillar component (DFC) with the help of RNA polymerase I complexes for pre-45S rRNA synthesis. The DFC exhibits role about processing of pre-45S rRNA into the mature 18S, 5.8S and 28S rRNA segments as key building blocks of eukaryotic ribosomes.
The DFC is also site for adaptive changes as methylation and pseudo-uridylation to organizing the rRNAs for their convergent integration into ribosomal domains. This DFC also activates the assembly of imported ribosomal proteins from the cytoplasm with the processed rRNAs to sustain the complete maturation of ribosomal subunits, where harmonized progression from rRNA processing to ribosome assembly occurs through interaction with granular component (GC) [11, 12].
The granular component (GC), which covers the DFC, is needful to complete the ribosome biogenesis, where maturation and combination of pre-ribosomal particles with nucleophosmin (B23) exhibiting key roles in this event. The size of the GC is directly related to the cell metabolism reflecting need to ribosome synthesis about cellular growth and cell division [13]. The regular integration between the nucleolar regions ascertain the normal production and maturation of functional ribosomes, which are essential for cellular homeostasis.
The recent researches identified nucleolar ‘vacuoles’ (NoVs) as membrane-less structures primarily observed in plants and some animals like Caenorhabditis elegans consists few granules and fibrils and association with FCs suggests a significant role in nucleolar dynamics [14]. Also, more recently identified nucleolar rim (NR) as distinct component signified by its special and more discrete proteomic composition is perhaps tether the nucleolus to chromatin and potentially impacting processes such as cell cycle regulation and ribosome biogenesis.
The recent studies aids in the complexity is physicochemical barcodes within the nucleolus that create unique micro-environments with varying features, which could uncover about additional nucleolar adaptations for diverse cellular functions [15]. The continuing research about emerging aspects about nucleolus structure can reveal vital role of each component in maintaining cellular function and supporting essential biological processes.
Communication and Interaction between Nucleolus and Nuclear Regions
The nucleolus not solely involved in ribosomal genesis, but also imparts an important role in the genome organization within the nucleus. This interacts with chromatin through nucleolar-associated domains (NADs) characterizes as domain of low transcriptional performance due to repressive transcription due to excessive histone modification consist causative H3K9me2 markers. This organization of repressive chromatin regions prevail such nucleolus, which easily divisible into euchromatic (active) and heterochromatic (inactive) regions [16]. The nucleolus is often observed as a centre for inactive chromatin and in such condition responding gene expression and preserving genomic fidelity.
The nucleolus operates as a central node that modulates localized pattern and activities of other nuclear domains as targeted depletion of specific genes led to nucleolar organization like localization of nuclear Cajal bodies, which are engaged in the maturation and modification of snRNAs and snoRNAs. These Cajal bodies are also needful for ribosome synthesis, telomere stability, transcriptional control, chromatin stress response and RNA breakdown [17] .
Nucleolar proteins regularly cycle between the nucleolus and the nucleoplasm, particularly in the cellular stress conditions and in case of such event nucleoplasmin (NPMI) translocated from the nucleolus to the nucleoplasm to reduce cellular stress involving the activation of the p53 signalling pathway as a critical stress response regulator. The damage in nucleolar structure in addition to impairing ribosome biogenesis but also cause abnormal nuclear architecture, defective signaling, and compromised cellular function [6].
The Nucleolar Proteins and their Functions
The diverse array of proteins in the nucleolus is crucial for revealing its functions and dynamics. The nucleolus contains both ribosomal proteins and enzymes involved in ribosome biogenesis, as well as a wide range of non-ribosomal proteins that contribute to processes like DNA repair, RNA processing, chromatin remodelling, transcription regulation and cell death [11, 18, 19, 20]. Comparative studies have shown that the composition and organization of the nucleolus can vary among species. For example, yeast nucleoli lack certain proteins found in higher Eukaryotes, reflecting evolutionary divergence in nucleolar function.
Both nucleophosmin (NPM1) and nucleolin play central roles in ribosome biogenesis, but their functions extend beyond this process. NPM1 regulates cell death pathways in response to cellular stress and DNA damage, specifically during apoptosis, where it is involved in both intrinsic and extrinsic apoptotic pathways. Mutations in NPM1, such as those found in acute myeloidleukaemia (AML), lead to its mislocalization, disrupting its ability to regulate apoptosis. Additionally, NPM1 acts as a chaperone of proteins involved in chromatin remodeling and DNA repair mechanisms and interacts with tumour suppressor factors like ARF [21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33].
Nucleolin is essential for cell growth and proliferation, regulating gene expression through interaction with nucleic acids involved in transcription and translation. Interestingly, nucleolin also facilitates viral replication by interacting with viral proteins, highlighting its role in viral pathogenesis [22]. Fibrillarin (FBL), a key player in rRNA methylation, is involved in the 2′-O-methylation of numerous pre-rRNA sites, a process crucial for ribosome biogenesis. As a core component of small nucleolar ribonucleoprotein (snoRNP) complexes, fibrillarin is vital for rRNA maturation and initiation of transcription by RNA polymerase I. Fibrillarin can undergo liquid–liquid phase separation (LLPS), facilitating the formation of membrane-less organelles like nucleolus [23]. This protein also interacts with p53 and ARF, suggesting a potential role in cancer progression [24]. Importantly, NPM1, nucleolin and fibrillarin all contain intrinsically disordered regions, which may regulate their localization and function within the nucleolus.
Recent studies have shown the presence of telomerase components in the nucleolus, suggesting that part of telomerase biosynthesis may occur there. This discovery has implications for cellular ageing and genomic instability, suggesting they influence telomere maintenance [20]. Moreover, some nucleolar proteins re-localize to the chromosomal periphery during mitosis, indicating a role in nucleolar disassembly and reassembly during cell division [19]. These proteins can be classified into two groups: those recruited early during pro-metaphase and those recruited later during post-metaphase. Understanding these recruitment dynamics helps clarify how the nucleolus disassembles and reassembles during the cell cycle.
Dynamic Nucleolar Organization Model as Liquid–Liquid Phase Separation
The nucleolus is a liquid-like substructure within cell assembled by LLPS pattern through modification of certain bio-molecules to arise distinct region which facilitates the formation of dense and dynamic liquid state not covered by membranes. The LLPS causes multivalent interactions within nucleus region involving intrinsically disordered regions (IDRs) of proteins and RNA jointly mediates the formation of dynamic membrane-less compartments [24, 25].
The nucleolus is organized into three functionally and physically distinct sub-compartments as the fibrillar centre (FC), dense fibrillar component (DFC) and granular component (GC)-whose spatial segregation is governed by LLPS (Figure 2). The FC is enriched in rDNA and gene transcription, while the surrounding DFC concentrates early RNA processing factors and small nucleolar RNPs that mediate initial transcription maturation. In parallel, the granular component (GC) forms through LLPS governed by nucleophosmin and related scaffold proteins which IDRs drive the assembly of liquid-like condensates that support the final steps of ribosome biogenesis [26].
The LLPS provides a versatile biophysical framework for regulating nuclear function by dynamically concentrating or excluding specific macromolecules in response to cellular cues beyond driving nucleolar structures [24]. Phase-separated condensates enhance reaction kinetics by increasing local concentrations of enzymes, substrates, and RNA, while simultaneously buffering stochastic fluctuations in gene expression. LLPS further enables rapid, reversible reorganization of nuclear architecture during stress, cell- cycle progression, and differentiation, thereby coordinating transcription, RNA processing, and ribo-nucleoprotein assembly in a context-dependent manner [25]. In addition, condensates can act as selective molecular filters, modulating the diffusion and residency time of regulatory factors to fine- tune signaling outputs and dysregulation of these material properties shifts condensates toward aberrant, less dynamic states, linking altered phase behavior to impaired genome maintenance, defective RNA metabolism, and the emergence of pathological protein aggregation observed in cancer and neurodegenerative disorders [27] (Table 1).
| Features | Fibrillar centre | Dense fibrillar component (DFC) | Granular component (GC) |
|---|---|---|---|
| Location in Nucleolus Appearance (EM) | Central pale region covered by DFC Light, less electron-dense | Covers FC as a dense lay Highly dense and fibrillar | Outer region of Nucleolus Moderately dense and granular |
| Main structural nature | Loosely packed chromatin- loops | Dense-fibrillar nature of nascent transcripts and proteins | Granule-rich matrix of RNP particles |
| Major contents | rDNA, RNA Poly1 & transcription factors (UBF, SL1) | Nascent pre-Rrna, fibrillarin, SnoRNPs, Nop56/58 | Pre-ribosomal particles (Pre-40S & Pre-60S), Ribosomal protein |
| rRNA transcription | rDNA generation and transcription initiation | Active elongation of pre-rRNA held | No transcription |
| rRNA Processing | No | Basic site of poly-rRNA and modification | Late-stage processing and maturation |
| Ribosomal subunit assembly | No | Start-site of basic assembly steps | Major-site of ribosome subunit assembly |
| Key molecular functions Associated proteins | Organization of rRNA and transcription works | Pre-rRNA co- translational processing | rRNA packaged with ribosomal proteins |
| Dynamic behavior (LLPS) Final product | UBF, RNA Poly I, TIF-1A Nucleation-site for phase- separation rRNA transcripts initiated | Fibrillarin, Nop- proteins, SnoRNPs Highly dynamic with liquid-like condensate Modified & processed pre-rRNA | Ribosomal proteins, assembly factors Most-fluid, droplet-like region Pre- 40S+Pre-60S subunits exported to cytoplasm |
Table 1: Comparative Nucleolar components: structure and functions.
Stress granules represent prototypical bio-molecular condensates formed through liquid–liquid phase separation (LLPS) that transiently sequester un-translated mRNAs and RNA-binding proteins to preserve cellular homeostasis during environmental stress. The persistent stress or disease-associated mutations can shift these assemblies from dynamic, liquid-like states toward more rigid, gel- or solid-like phases, thereby impairing granule clearance and normal RNA metabolism. Such aberrant phase transitions promote the accumulation of insoluble protein aggregates and are increasingly implicated in the pathogenesis of neurodegenerative disorders, including amyotrophic lateral sclerosis and fronto-temporal dementia. Thus, LLPS not only enables adaptive stress responses but also constitutes a critical vulnerability point where altered condensate material properties can precipitate cellular dysfunction and disease [26].
The Evolutionary Origins of the Nucleolus
The nucleolus is the most prominent compartment of the eukaryotic nucleus and arises through self-organization around actively transcribed ribosomal DNA (rDNA) loci.
The distinctive architecture of nucleolus suggests that the nucleolus did not emerge as a static organelle but rather evolved as a dynamic, self-assembling ribonucleoprotein condensate, shaped by increasing cellular and genomic complexity in eukaryotes.
From an evolutionary perspective, the nucleolus likely originated from primitive ribosome biogenesis centers present in ancestral prokaryotic-like cells. In bacteria, rRNA transcription, processing, and ribosome assembly occur in spatially enriched regions termed nucleoid- associated ribosome assembly zones, although these lack stable compartmentalization [30]. Nevertheless, recent studies indicate that bacterial cells exhibit functional micro- domains enriched in ribosome biogenesis factors, suggesting that spatial organization of ribosomal production predates the emergence of the eukaryotic nucleus [34]. These early organizational tendencies may represent evolutionary precursors to the nucleolus, where clustering of rRNA synthesis and processing activities conferred kinetic and regulatory advantages (Figure 2).
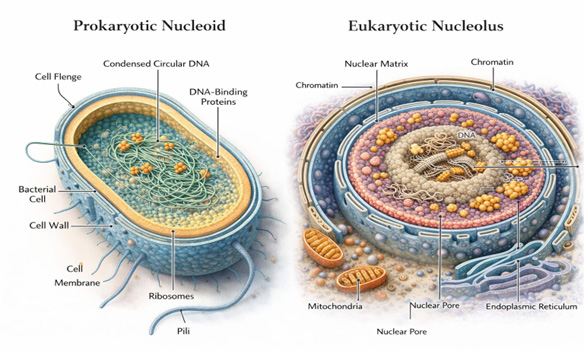
The transition from prokaryotic to eukaryotic cellular organization, marked by the appearance of the nuclear envelope and linear chromosomes, likely intensified selective pressure for spatial coordination of ribosome biogenesis. Expansion of genome size, increased gene regulatory complexity, and separation of transcription and translation created a need for specialized nuclear sub-domains that could efficiently integrate rRNA synthesis with processing and ribonucleoprotein assembly [35, 36]. The nucleolus, in this context, can be viewed as an emergent structure arising from the concentration of rDNA repeats and associated processing machinery within confined nuclear space.
Comparative genomics provides further insight into nucleolar evolution. Core components of ribosome biogenesis including RNA polymerase I subunits, small nucleolar RNAs (snoRNAs), ribosomal proteins, and processing enzymes are highly conserved across eukaryotes and share ancestry with bacterial ribosomal assembly factors [35]. However, eukaryotes possess an expanded repertoire of nucleolar proteins enriched in intrinsically disordered regions (IDRs) and low-complexity domains, which facilitate multivalent interactions and promote phase separation. This expansion suggests that the evolution of protein disorder was instrumental in enabling the formation of stable yet dynamic nucleolar compartments, supporting the hypothesis that biophysical self-assembly mechanisms were co-opted to generate higher-order nuclear organization.
The LLPS offers a parsimonious mechanism by which cellular systems can generate compartmentalization without membranes, relying instead on physicochemical properties of macromolecules. The emergence of LLPS-capable proteins may therefore have been a critical step in transforming dispersed ribosome biogenesis reactions into spatially coherent nucleolar condensates [26].
Phylogenetic analyses further indicate that structured nucleoli appear only in eukaryotes, whereas simpler ribosome assembly zones exist in archaea. Interestingly, some archaeal species possess nucleolus-like bodies enriched in rRNA processing factors, suggesting an intermediate organizational state [33]. These observations support a gradualist model in which increasing molecular crowding, genome expansion, and diversification of RNA- processing pathways favored the stabilization of ribosome biogenesis condensates into discrete nuclear bodies. Thus, the nucleolus likely evolved not as a novel invention but as an elaboration of pre-existing spatial clustering mechanisms that became progressively more refined.
The co-evolution of nucleolar structure and rDNA organization is also notable. Eukaryotic genomes typically contain hundreds of tandemly repeated rDNA units, which serve as nucleation platforms for nucleolar assembly. These repeats enable high transcriptional output while simultaneously providing a scaffold for recruitment of processing factors. Evolutionary amplification of rDNA arrays may therefore have reinforced nucleolar formation by enhancing local macromolecular concentration, creating positive feedback between gene organization and nuclear architecture [37]. In this sense, the nucleolus represents an example of how genome structure and cellular compartmentalization co-evolve to optimize essential biosynthetic processes.
Beyond ribosome production, the nucleolus has acquired additional regulatory roles during evolution, including involvement in cell-cycle control, stress sensing, and sequestration of regulatory proteins. These expanded functions likely emerged as secondary adaptations once nucleolar condensates became stable nuclear landmarks [38, 39]. The capacity of the nucleolus to act as a molecular reservoir or stress-responsive hub may reflect selective advantages conferred by pre-existing phase-separated architecture, which could be readily repurposed for regulatory integration. Such functional diversification underscores how biophysical properties can facilitate evolutionary innovation without requiring extensive structural redesign.
Importantly, the evolutionary persistence of the nucleolus across all extant eukaryotic lineages highlights its fundamental role in cellular fitness. Any perturbation of nucleolar integrity directly compromises ribosome biogenesis and protein synthesis, imposing strong selective constraints on its maintenance. At the same time, variability in nucleolar size, number, and organization among species indicates evolutionary plasticity, allowing adaptation to differing metabolic demands and developmental strategies. This balance between conservation and flexibility reflects the dual nature of the nucleolus as both an indispensable biosynthetic center and a tunable regulatory platform.
In summary, the nucleolus likely emerged through the evolutionary stabilization of ancestral ribosome biogenesis clusters, driven by genome expansion, increased regulatory complexity, and the appearance of proteins capable of multivalent, phase-separating interactions. Rather than being a static organelle defined by rigid boundaries, the nucleolus represents an emergent, self-organizing nuclear condensate whose architecture and functions have been progressively refined throughout eukaryotic evolution. This perspective not only clarifies the origins of nucleolar compartmentalization but also illustrates how fundamental biophysical principles can shape cellular organization over evolutionary timescales.
Conclusions: the Nucleolus in Focus
The continuing research into the nucleolus and its evolutionary origins remains essential for resolving fundamental questions about this structure and its broader biological and clinical significance [40]. This review has examined the nucleolus through structural, functional, and evolutionary lenses, underscoring its central role not only in ribosome biogenesis but also in an expanding array of cellular processes. Far from being a passive ribosome factory, the nucleolus emerges as a dynamic and multifunctional nuclear domain whose complexity reflects both deep evolutionary conservation and remarkable adaptability. Elucidating its origins and functions therefore provides critical insight into core principles of cellular organization [28].
Rapid advances in molecular labelling, high-resolution imaging, and integrative omics technologies are transforming our ability to interrogate the nucleolus in space and time, revealing its highly dynamic and context-dependent nature [29, 41, 42, 43, 44]. While its canonical roles in rRNA transcription, ribosome assembly, and cell-cycle regulation are well established, increasing evidence points to diverse non- canonical functions, including roles in stress sensing, protein quality control, and regulatory sequestration. These emerging functions remain incompletely characterized, yet they are likely to be central to understanding how nucleolar organization integrates cellular metabolism, signaling, and environmental responsiveness [39].
Addressing unresolved questions surrounding nucleolar function and evolution also emphasizes its critical contribution to cellular homeostasis [18]. Comparative analyses across phylogenetic lineages, developmental stages, and physiological states may uncover fundamental design principles linking nucleolar architecture to cellular complexity, robustness, and adaptability [31]. Such perspectives are particularly important for understanding how perturbations in nucleolar structure and activity contribute to disease [7, 8].
Ultimately, continued exploration of the nucleolus promises to advance not only basic cell and evolutionary biology but also translational research. By illuminating how this ancient, self-organizing nuclear condensate coordinates essential biosynthetic and regulatory pathways, future studies may identify novel molecular vulnerabilities and therapeutic strategies relevant to cancer, neuro-degeneration, and other nucleolus-associated pathologies. References
1. Scheer U, Hock R (1999) Structure and function of the nucleolus. Curr Opin Cell Biol 11: 385-390.
2. Tartakoff A, DiMario P, Hurt E, McStay B, Panse VG, et al. (2022) The dual nature of the nucleolus. Genes Dev 36: 765-769.
3. Dubois ML, Boisvert FM (2016) The nucleolus: structure and function. In: Bazett-Jones DP, Dellaire G (Eds.), The functional nucleus Cham, Switzerland: Springer International Publishing, pp: 29-49.
4. Massenet S (2019) In vivo assembly of eukaryotic signal recognition particle: a still enigmatic process involving the SMN complex. Biochimie 164: 99-104.
5. Grummt I (2013) The nucleolus-guardian of cellular homeostasis and genome integrity. Chromosoma 122: 487-497.
6. Yang K, Yang J, Yi J (2018) Nucleolar stress: hallmarks, sensing mechanism and diseases. Cell Stress 2: 125-140.
7. Stochaj U, Weber SC (2020) Nucleolar organization and functions in health and disease. Cells 9: 2073-4409.
8. Núñez Villacís L, Wong MS, Ferguson LL, Hein N, George AJ, et al. (2018) New roles for the nucleolus in health and disease. Bioessays 40: e1700233.
9. Thiry M, Lamaye F, Lafontaine DLJ (2011) The nucleolus: when 2 became 3. Nucleus 2: 289-293.
10. Radhakrishnan S, Martin CA, Rammohan A, Vij M, Chandrasekar M, et al. (2023) Significance of nucleogenesis, ribogenesis, and nucleolar proteome in the pathogenesis and recurrence of hepatocellular carcinoma-Expert Review. Gastroenterol Hepatol 17(4): 363-378.
11. Raška I, Shaw PJ, Cmarko D (2006) New insights into nucleolar architecture and activity. In International review of cytology: a survey of cell biology, Amsterdam, The Netherlands: Elsevier, pp. 177-235.
12. Hernandez-Verdun D (2006) Nucleolus: from structure to dynamics. Histochem. Cell Biol 125(1-2): 127-137.
13. Sirri V, Urcuqui-Inchima S, Roussel P, Hernandez-Verdun D (2008) Nucleolus: the fascinating nuclear body. Histochem. Cell Biol 129(1): 13-31.
14. Xu D, Chen X, Kuang Y, Hong M, Xu T, et al. (2023) rRNA intermediates coordinate the formation of nucleolar vacuoles in C. elegans. Cell Rep 42 (8): 112915.
15. King MR, Ruff KM, Pappu RV (2024) Emergent micro- environments of nucleoli. Nucleus 15(1): 2319957.
16. Vertii A, Ou J, Yu J, Yan A, Pagès H, et al. (2019) Two contrasting classes of nucleolus-associated domains in mouse fibroblast heterochromatin. Genome Res 29(8): 1235-1249.
17. Wang C, Ma H, Baserga SJ, Pederson T, Huang S (2023) Nucleolar structure connects with global nuclear organization. Mol Biol Cell 34(12): ar114.
18. Hua L, Yan D, Wan C, Hu B (2022) Nucleolus and nucleolar stress: from cell fate decision to disease development. Cells 11(19): 3017.
19. Saraogi I, Shan Sou (2011) Molecular mechanism of co- translational protein targeting by the signal recognition particle. Traffic 12(5): 535-542.
20. Rubtsova M, Dontsova O (2020) Human telomerase RNA: telomerase component or more?. Biomolecules 10(6): 873.
21. Gibbs E, Miao Q, Ferrolina M, Bajpai R, Hassan A, et al. (2023) p14ARF forms meso-scale assemblies upon phase separation with NPM1. Res Sq 3: 3592059.
22. Abdelmohsen K, Gorospe M (2012) RNA-binding protein nucleolin in disease. RNA Biol 9(6): 799-808.
23. Hernández-Guzmán J, Arias CF, López S, Sandoval-Jaime C (2024) Nucleolin-RNA interaction modulates rotavirus replication. J Virol 98(2): e0167723.
24. Guillen-Chable F, Bayona A, Rodríguez-Zapata LC, Castano E (2021) Phase separation of intrinsically disordered nucleolar proteins relate to localization and function. Int J Mol Sci 22: 1422-0067.
25. Nguyen Van Long F, Lardy-Cleaud A, Carène D, Rossoni C, Catez F, et al. (2022) Low level of Fibrillarin, a ribosome biogenesis factor, is a new independent marker of poor outcome in breast cancer. BMC Cancer 22(1): 526.
26. Brangwynne CP, Mitchison TJ, Hyman AA (2011) Active liquid-like behavior of nucleoli determines their size and shape in Xenopus laevis oocytes. Proc Natl Acad Sci USA 108(11): 4334-4339.
27. Feric M, Vaidya N, Harmon TS, Mitrea DM, Zhu L, et al. (2016) Coexisting liquid phases underlie nucleolar sub- compartments. Cell 165(7): 1686-1697.
28. Mitrea DM, Kriwacki RW (2016) Phase separation in biology; functional organization of a higher order. Cell Commun Signal 14: 1-31.
29. Yoneda M, Nakagawa T, Hattori N, Ito T (2021) The nucleolus from a liquid droplet perspective. J Biochem 170(2): 153-162.
30. Shin Y, Brangwynne CP (2017) Liquid phase condensation in cell physiology and disease. Science 357(6357): eaaf4382.
31. Pandey P, Gupta R, Tiwari S, Gupta A, Mondal S, et al. (2024) Carbon concentrating mechanism in cyanobacteria: necessity and evolution. Cyanobacteria pp: 57-67.
32. Huber ST, Terwiel D, Evers WH, Maresca D, Jakobi AJ (2023) Cryo-EM structure of gas vesicles for buoyancy- controlled motility. Cell 186(5): 975-986.
33. Kuzminov A (2024) Bacterial nucleoid is a riddle wrapped in a mystery inside an enigma. J Bacteriol 206(3): e0021123.
34. Sanapala P, Pola S (2021) Understanding the cell organization in planctomycetes. In Developmental biology in prokaryotes and lower eukaryotes. In: TG Villa, T De Miguel Bouzas (Eds.), Cham, Switzerland: Springer International Publishing, pp: 229-248.
35. Scott DD, Oeffinger M (2016) Nucleolin and nucleophosmin: Nucleolar proteins with multiple functions in DNA repair. Biochem Cell Biol 94(5): 419- 432.
36. Feng JM, Yang CL, Tian HF, Wang JX, Wen JF (2020) Identification and evolutionary analysis of the nucleolar proteome of Giardia lamblia. BMC Genom 21(1): 269.
37. Martínez-Calvillo S, Florencio-Martínez LE, Nepomuceno-Mejía T (2019) Nucleolar structure and function in trypanosomatid protozoa. Cells 8(5): 421.
38. Jin DJ, Mata Martin C, Sun Z, Cagliero C, Zhou YN (2017) Nucleolus-like compartmentalization of the transcription machinery in fast-growing bacterial cells. Crit Rev Biochem Mol Biol 52(1): 96-106.
39. Riback JA, Zhu L, Ferrolino MC, Tolbert M, Mitrea DM, et al. (2020) Composition-dependent thermodynamics of intracellular phase separation. Nature 581(7807): 209- 214.
40. Carmo-Fonseca M, Mendes-Soares L, Campos I (2000) To be or not to be in the nucleolus. Nat Cell Biol 2: E107-E112.
41. Tsai RYL, Pederson T (2014) Connecting the nucleolus to the cell cycle and human disease. FASEB J 28(8): 3290- 3296.
42. Corman A, Sirozh O, Lafarga V, Fernandez-Capetillo O (2023) Targeting the nucleolus as a therapeutic strategy in human disease. Trends Biochem Sci 48(3): 274-287.
43. Pederson T (2011) The nucleolus Cold Spring Harb. Perspect Biol 3(3): a000638.
44. Van Teeseling MCF, Mesman RJ, Kuru E, Espaillat A, Cava F, et al. (2015) Anammox planctomycetes have a peptidoglycan cell wall. Nat Commun 6: 6878.
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica