Alpha-Synuclein Overexpression Causes Mild Hearing Loss and Protection from Noise Trauma
Alpha synuclein (α-Syn) is a small synaptic protein expressed within the central nervous system (CNS). α-Syn has been implicated in the pathogenesis of Parkinson’s disease (PD), and current theory suggests that α-Syn over-expression may lead to some of the CNS changes seen in PD, but the precise function of α-Syn remains elusive. α-Syn has also been identified in the auditory system and it is predominantly localized to the efferent auditory synapses below outer hair cells. In these studies, we studied a transgenic mouse model over-expressing α-Syn to investigate whether over-expression of α-Syn leads to pathologic changes within the cochlea. In the cochleae of wild-type (WT) mice, immunofluorescence using an anti-α-Syn antibody revealed strong labeling only in the base of outer hair cells. In contrast, in mice over-expressing α-Syn, there was robust staining in cells throughout the organ of Corti. Auditory brainstem responses (ABR) showed significant threshold elevations and a significant decrease of the ABR wave I amplitude in the transgenic mice as compared to WT mice. No significant differences were seen between WT and transgenic mice in hair cell counts, morphology or distortion product otoacoustic emissions. Following noise exposure, WT mice demonstrated an ABR threshold shift of 10-20dB whereas transgenic mice showed no significant threshold shift, suggesting a protective effect of α-Syn over-expression against noise-induced hearing loss. Together, these studies demonstrate that α-Syn over-expression causes a mild hearing loss and a protective effect against noise damage with no apparent damage to the cochlear hair cells. These findings suggest that the hearing loss seen with α-Syn over-expression occurs more centrally, while the mechanism of noise protection with over-expression remains unknown.
Introduction
Synucleins are a family of short proteins that share a high level of sequence homology with each other and across species) [1, 2]. There are four recognized members of the synuclein family, termed α-synuclein, β-synuclein, γ-synuclein and synoretin [3, 4, 5, 6]. Both α- and β-synuclein are principally localized to the central nervous system (CNS), γ-synuclein is found in the peripheral nervous system and synoretin is present in the retina [6, 7, 8].
α-synuclein (α-Syn) has been implicated as a component of pathological inclusions found in many neurodegenerative disorders such as Parkinson’s disease (PD), dementia with Lewy bodies and multisystem atrophy, although its precise function and the mechanism underlying its aggregation during the process of these diseases is not fully understood [9, 10]. The other members of the synuclein family have also been implicated in the development of the pathological changes characteristic of neurodegenerative disorders [11, 12, 13]. α-Syn mRNA is highly expressed in the brain and lower levels of expression are seen in other tissues such as the pancreas, kidney, skeletal muscle, lung, placental, and heart, suggesting functionality in these tissues as well [14, 15]. Co-localization of α-Syn and synaptophysin has been demonstrated during neural development, suggesting spatially regulated functions of these proteins, perhaps related to synaptic specialization or complementary roles in regulating presynaptic components [16].
α-Syn also has been shown to be involved in glutamate release [17]. Other work has suggested that α-Syn may be required for the genesis or maintenance of presynaptic neurotransmitter vesicle numbers and mobilization [8, 18]. These studies led to the hypothesis that α-Syn may be involved in synaptic development and/or function.
With the well-described role of cholinergic and catecholaminergic neuronal signaling within the cochlear organ of Corti [19, 20], as well as synuclein’s presumed role in Ca2+ homeostasis and effects on voltage-dependent Ca2+ influx [21, 22], it is not surprising that synucleins have also been identified in the auditory system. Previously, we demonstrated the presence of α-, β- and γ-synuclein within the cochlea [23]. In that initial study, all 3 isoforms were predominantly localized to the efferent auditory synapses below outer hair cells. In subsequent studies on an α-Syn knockout mouse, there was no evidence of hearing loss or cochlear abnormalities through post-natal day 21 (P21), providing no additional insight into the role of α-Syn in auditory function [24]. One current theory regarding the link between PD and α-Syn postulates that over-expression of the protein leads to the CNS changes seen Park SN, et al. [24]; Singleton AB, et al. [25]; Chartier Harlin MC, et al. [26]; Maraganore DM, et al. [27]. A prior study in a transgenic mouse over-expressing α-Syn demonstrated pathological changes in the spinal and brainstem motoneurons [28, 29]. In a related study using this same transgenic mouse over- expressing α-Syn, neurotransmitter release in the brain was found to be inhibited and this was hypothesized to lead to some of the symptoms associated with PD [30]. In the present study, we perform further studies on this same transgenic mouse to investigate whether α-Syn over-expression affects the cochlea. Here we report that α-Syn overexpression is associated with both mild hearing loss and also a protective effect against noise-induced hearing loss. The absence of pathological changes to cochlear hair cells suggests that α-Syn over-expression effects hearing at the level of the central auditory pathways, though precisely where remain uncertain. Lastly, these findings lead to two major questions: 1. Why does over expression of α-syn lead to hearing loss; and 2. Why does this confer protection against a permanent threshold shift from noise?
Material and Methods
Animals
FVB alpha synucleins (α-Syn) over-expression mice were provided by Dr. Robert Edwards of the University of California, San Francisco, and Department of Neurology. The general and central nervous system phenotype of this mouse has been described previously Nemani VM, et al. [30]. All procedures and animal handling were done according to national ethical guidelines and were approved and complied with all protocol requirements of the University of California, San Francisco Institutional Animal Care and Use Committee.
Auditory Tests
Hearing tests were performed as previously described Akil O, et al. [31] with α-Syn over-expression (transgenic) and wild-type (WT) mice littermates at different postnatal ages (1-10 months (P1-P10mo)). Briefly, all auditory testing was performed in a sound-proof chamber. Before acoustic testing, mice were anesthetized by intraperitoneal injection of a mixture of Ketamine hydrochloride (Ketaset, 100 mg/ ml) and xylazine hydrochloride (xyla-ject, 10mg/ml), with supplements of one fifth the original dose administered as required. Body temperature was maintained with a heating pad and monitored with a rectal probe throughout recordings. Auditory Brainstem Response Measurement: The auditory brainstem response (ABR) thresholds were differentially recorded from the scalp of the transgenic and WT mice littermates. Responses were recorded using subdermal needle electrodes at the vertex, below the pinna of the left ear (reference), and below the contralateral ear (ground). The sound stimuli were clicks (5ms duration; 31Hz) and tone pips at 8, 16, and 32 kHz (10ms duration; cos2 shaping; 21Hz). Measurements were recorded using the TDT BioSig III system (Tucker Davis Technologies). For each stimulus, electroencephalographic (EEG) activity was recorded for 20 ms at a sampling rate of 25kHz, filtered (0.3- 3kHz), and waveforms from 512 stimuli were averaged for click responses, and 1000 stimuli for tonal stimuli (8, 16 and 32 kHz). ABR waveforms were recorded in 5 dB sound pressure level (SPL) intervals down from the maximum amplitude. The threshold was defined as the lowest stimulus level at which response peaks for wave’s I-V were clearly and reproducibly present upon visual inspection. These threshold judgments were confirmed by analysis of stored waveforms. Data were obtained from the transgenic (n=9), and WT (n=6) mice littermates at varying ages including P30 to P10months. The comparisons of animal groups were performed using 1-way ANOVA with Bonferroni post-hoc testing. Significance was defined at p <0.05. To asses changes in ABR latencies, the ABR peaks and troughs were identified automatically by cursers programmed to identify each peak using the Tucker-Davis Technologies system software, and then verified by visual inspection of the recorded wave forms. Latencies were calculated from the onset of the stimulus, and the interpeak latency calculated by subtraction of wave II-I (P2 - P1). Differences in interpeak latency were then calculated at two different intensity levels (90 and 80 dB SPL). ABR Wave I amplitude analysis was also done by peak- to-peak measurement of the stored click ABR waveforms at two different intensity levels (90 and 80 dB SPL). Compound Action Potential Recording: Compound action potential (CAP) recording was performed as previously described Seal RP, et al. [32]. A dorsal surgical approach Akil O, et al. [33] was used to expose the left cochlea of the transgenic (n=6) and WT mice littermates (n=4) for recordings. After making a small opening in the bulla and visualizing the round window niche, a fine recording electrode, fashioned from Teflon coated-insulated silver wire, was positioned in the round window niche and another was placed in the soft tissue of the neck as a ground electrode. The sound stimulus was generated with Tucker-Davis System II hardware and software (Tucker-Davis). The CAP thresholds were measured using click stimuli. Response to these inputs were measured at the round window of the transgenics at P30 and compared to responses obtained from the WT littermate’s mice. Distortion Product Otoacoustic Emissions Measurement: The distortion product otoacoustic emissions (DPOAEs) were measured using an acoustic probe placed in the left external auditory canal. Stimuli consisted of two primary tones (f1/f2=1.25) digitally synthesized at 100 kHz using SigGen software. The primary tones with geometric mean (GM) frequencies ranging from 6 to 36 kHz and equal levels (L1 = L2 = 60 dB SPL) were presented via two separate speakers (EC1; Tucker Davis Technologies, Alachua, FL) to the acoustic probe. DPOAE 2f1–f2 responses were recorded using an ER10B (Etymotics Research, Elk Grove Village, IL) microphone assembly within the acoustic probe and the TDT BioSig III system. Responses were amplified, digitally sampled at 100 kHz, and averaged over 50 discrete spectra. Fast Fourier transforms were computed from averaged responses. For each stimulus set, the DPOAE amplitude level at 2f1–f2 was extracted, and sound pressure levels for data points 100 Hz above and below the DPOAE frequency were averaged for the noise floor measurements. DPOAE levels were plotted as a function of primary tone GM frequency. Statistical analyses were performed using ANOVA with Bonferroni post-hoc tests and significance defined as p < 0.05. Data were collected from the transgenic (n=9), and WT (n=6) mice at P30.
Noise Induced Hearing Loss
8-week-old transgenic and WT mice were exposed awake and unrestrained within small cells in a subdivided stainless- steel mesh cage with constant rotation. Noise stimuli (4-20 kHz) were presented at 105 dB peak equivalent SPL for 30 minutes in order to generate a permanent threshold shift. Stimuli were generated by a personal computer connected to a waveform generator (Tucker DavisTechnologies, Alachua, FL), amplified and delivered through a speaker inside the noise booth. ABR threshold was measured prior to and after 72 hours and 26 days of noise exposure. Data were collected from the transgenic (n=6), and WT (n=6) mice.
α-Synuclein RNA Expression
Reverse Transcriptase-Polymerase Chain Reaction: The total RNA harvested from both the transgenic and WT mice cochleae extractions (Trizol™, Invitrogen Corp) was reverse transcribed with superscript II RNase H− (Invitrogen) for 50 min at 42°C, using oligodT primers. Two microliters of RT reaction product was used for subsequent polymerase chain reaction (PCR; Taq DNA Polymerase, Invitrogen) of 35 cycles using the following parameters: 94°C for 30 s, 60°C for 45 s, 72°C for 1 min, followed by a final extension of 72°C for 10 min and storage at 4°C. Primers were designed to amplify a unique sequence of mouse α-synucleins. The PCR primers that were used to amplify mouse α-synuclein (NM_009221) were: forward primer (1)-gtggagcaaaaatacatctttag and reverse primer (1) -gctgctcttccatggcgtaca primers that were designed to amplify a 505-bp fragment spanning exon 2 through the 5′ portion of exon 6 and Forward primer (2)gtggagcaaaaatacatctttag and reverse primer (2)-tgtacgccatggaagagcagc primers that were designed to amplify a 397-bp fragment spanning exon 2 through the 5′ portion of exon 5. Controls were cochlear RNA without reverse transcriptase. Analysis of each PCR sample was then performed on 2% agarose gels containing 0.5 μg/ml ethidium bromides. Gels were imaged using a digital Camera and image processing system (Kodak, Rochester NY, USA). Candidate bands were cut out and the DNA was extracted (Qiaquick gel extraction kit, Qiagen) and sequenced (Elim Biopharmaceuticals, Inc. Hayward, CA, USA). The PCR product was then compared directly to the full mouse α-Syn sequence for identity.
Quantitative PCR: For real-time, quantitative PCR, cochlea from the transgenic and WT mice was dissected. RNA was extracted and processed for RT as described above. The expression level of α-Syn was measured by the iQ5 real- time PCR (Bio-Rad, Hercules, CA, USA). Amplification was performed in a total volume of 25 μl containing 12.5 μl SYBR Green I amplification master mix (Bio-Rad), 250 nM primers of mouse α-synuclein (NM_009221). Forward- gtgacaaatgttggaggagc (100% identical to the human) and reverse -tggatcctggcagtgaggctt (90% identical to human) primers were designed to amplify a 186-bp fragment spanning exon 4 through the 5′ portion of exon 5, and ribosomal protein L19 (BC083131); and forward— atgtatcacagcctgtacctg, and reverse-ttcttggtctcttcctccttg primers were designed to amplify a 232-bp fragment, and 3 μl of 1:5 diluted cDNA as a template. The amplifications were performed in triplicate wells, using a program that consisted of initial denaturation at 95°C for 3 min followed by a 40-cycle of denaturation at 95°C for 10 s and annealing at 61°C for 30 s. After amplification, a melting curve was generated for every PCR product to check the specificity of the PCR reaction (absence of primer dimers or other nonspecific amplification product). The 2-ΔΔCT method was used to calculate relative fold difference in α-Syn RNA expression determined from real-time quantitative PCR experiments normalized to the ribosomal RNA L19, an internal control, relative to the expression level of α-Syn [34]. Statistical differences were calculated using a one-way ANOVA with Bonfferoni post hoc test with significance noted for p<0.05.
α-Synuclein Protein Expression
Immunofluorescence: Transgenic and WT mice littermates at P30 were anesthetized and their cochleae were isolated, dissected, perfused through the oval and round windows by 2% paraformaldehyde in 0.1M phosphate buffer (PB) at pH 7.4 and incubated in the same fixative for 2 hrs. After fixation, cochleae were rinsed with PB and immersed in 5% EDTA in 0.1M PB for decalcification. When cochleae were fully decalcified (~2 days), they were incubated overnight in 30% sucrose for cryoprotection. The cochleae then were embedded in O.C.T. Tissue Tek Compound (Miles Scientific). Specimens were cryosectioned parallel to the modiolus at 10-12 µm thickness for immunofluorescence, mounted on Superfrost™ microscope slides (Erie Scientific, Portsmouth NH) and stored at -20°C until use.
For immunofluorescent staining of the transgenic and WT cochlea sections, a monoclonal anti-α-synculein antibody against rat synuclein-1 (BD Biosciences cat no. 610786) that recognizes both human and endogenous mouse proteins, and a rabbit anti-neurofilament 200 (NF- 200; Chemicon, Temecula, CA, USA) antibody that labels afferent as well as some efferent auditory fibers [35], were used. After incubation at 37°C for 30 min, the sections were rinsed twice for 5 min in 0.1 M PBS pH 7.4, and then preincubated for 1 hr in the blocking buffer (0.3% Triton X100 and 30% normal goat serum). The sections were then incubated in a humid chamber overnight at 4°C with the mouse anti-α-synculein antibody at 1:300 and the rabbit anti-neurofilament 200 at 1:200 (diluted in the blocking buffer). Slides were then rinsed (2×10 min) and incubated for 2 hrs at room temperature (RT) with goat anti-mouse IgG antibody conjugated to Cy3 and goat anti-rabbit IgG antibody conjugated to Cy2 diluted to 1:2000. Following the removal of the secondary antibodies and prior to the final washes, the sections were exposed for 15 min at RT to the fluorescent dye 4, 6-diamidino-2-phenylindole (DAPI, 1.5μg/ml in PBS, Sigma) which allowed visualization of the nuclei along with immunofluorescence. The sections were then rinsed in PBS (2×10 min) and mounted in equal parts glycerol/PBS and cover slipped. The slides were observed with an Olympus microscope with confocal immunofluorescence. Cochlea Whole-Mount Immunofluorescence: Transgenic and WT mouse cochleae were perfused with 4% PFA in 0.1 M PB, pH 7.4, and remained in the fixative for 2 hrs at 4°C. Cochleae were washed with PB three times for 10 min and then decalcified with 5% EDTA in 0.1 M PBS for ~2 days. Following decalcification, the otic capsule was removed, followed by removal of the lateral wall, Reissner’s membrane, and the tectorial membrane. The remaining organ of Corti was incubated overnight at 4°C with the rabbit anti-myosin VIIa antibody (a hair-cell-specific marker; 1:50 dilution in PBS; Proteus Biosciences, catalog #25-6790). Whole-mount cochleae were rinsed twice for 10 min with PB and then incubated for 2hrs at RT with goat anti-rabbit IgG antibody conjugated to Cy2 (1:2000 dilutions in PB; Jackson ImmunoResearch, 111- 165-003). The whole mounts were rinsed with PB twice for 15 min and incubated with rhodamine–phalloidin (stock solution of 200 U/ml methanol, diluted 1:100 in PB for working solution (Invitrogen)) for 1 hr. Whole mounts were then rinsed with PB, further microdissected into individual turns for surface preparation, and then incubated for 15 min at RT with the fluorescent DAPI to mark nuclei. The cochlear whole mounts were rinsed in PB and mounted on glass slides in antifade FluorSave reagent (Calbiochem, 34589). Hair cells in the organ of Corti were visualized with an Olympus microscope with confocal immunofluorescence. Western Blot Analysis: Total protein extract was prepared from the transgenic and the WT mice cochlea using a lysis buffer containing (20 mM Tris HCl pH 7.5–8, 150 mM NaCl, 0.5% sodium deoxycolate, 1% Triton X100, 0.1 sodium dodecyl sulfate (SDS), 1 mM EDTA, 1 mM phenylmethylsulphonyl fluoride, and protease inhibitor cocktail). The cochlear homogenate was spun 10 min at 14,000 RPM at 4°C and the protein concentration in the supernatant was determined by Bradford assay. Approximately 100 μg of the supernatant was fractioned by SDS-polyacrylamide gel electrophoresis (PAGE) and transferred onto nitrocellulose membrane. The blot membrane was incubated for 1 hr in blocking buffer containing 5% nonfat dry milk in PBS-T (phosphate buffer saline + 0.1% Tween 20) and then incubated overnight at 4°C with the monoclonal anti- α-synculein antibody (same antibody used for immunofluorescence) and with the rabbit anti-β-actin antibody diluted in 1% BSA in PBS-T. The blot was washed extensively 5 times with PBS-T and incubated with secondary IRDye fluorescent antibodies (Licor) (Alexa Fluor 680-conjugated anti-rabbit IgG (Molecular Probes) and IRDye800 conjugated anti-mouse IgG (Rockland, Gilbertsville, PA)). The membrane was exposed using the Odyssey Fc imaging system. α-synculein and β-actin signal intensities were analyzed by using the Odyssey infrared image system (LiCor) and quantified with Odyssey software.
Histology
Light Microscopy: Transgenic and WT mice littermates at P30 were anesthetized and their cochleae were isolated, dissected, perfused through the round and oval windows with a solution of 2.5% paraformaldehyde and 1.5% glutaraldehyde in 0.1 M phosphate buffered (PB) solution at pH 7.4, and then placed in the same fixative overnight at 4°C. The cochleae were rinsed with 0.1 M PB and post fixed in 1% osmium tetroxide for 2 hrs. Rinsed again and immersed in 5% EDTA for decalcification. The decalcified cochleae were then dehydrated in ethanol and propylene oxide and embedded in Araldite 502 resin (Electron Microscopy Sciences, Fort Washington, PA). Sections were cut at 5 µm thickness parallel to the cochlear modiolus. Finally, sections were stained with Toluidine Blue and mounted with Permount (Fisher Scientific, Houston, TX) on microscope slides and visualized with a Leica microscope. Electron Microscopy: The P30 transgenic and WT mice cochleae were surgically exposed, a stapedectomy was performed, and the round window perforated. Temporal bones were preserved by gentle in vivo perfusion of the fixative (2.5% paraformaldehyde and 1.5% glutaraldehyde buffered to pH 7.4 with 0.1M phosphate) through the perilymphatic channels, followed by post-fixation with a solution of 1% osmium tetroxide and 1.5% potassium ferricyanide phosphate buffered solution (potassium ferricyanide was used to improve contrast). The cochleae were decalcified, dehydrated, and then embedded in Epon. Thick sections (1μm) were mounted onto glass slides and counterstained with toluidine blue. Once the cochlear organ of Corti is observed, several ultrathin sections (silver to gray) were collected on Formvar films on slotted grids and stained with saturated aqueous 2% uranyl acetate and 0.4% lead citrate in 0.15N sodium hydroxide. The stained sections were then examined at 60 kV in a JEOL-JEM 100S transmission electron microscope.
Results
α-Syn Expression in the Cochlea
α-Syn RNA Expression: Reverse-transcriptase (RT)-PCR was used to screen for the presence of α-Syn within the microdissected cochlea of the WT and transgenic (mutant) mice at P30. As shown in (Figure 1A), α-Syn mRNA is identified in the whole cochlear extracts for two different set of primers (1 and 2).
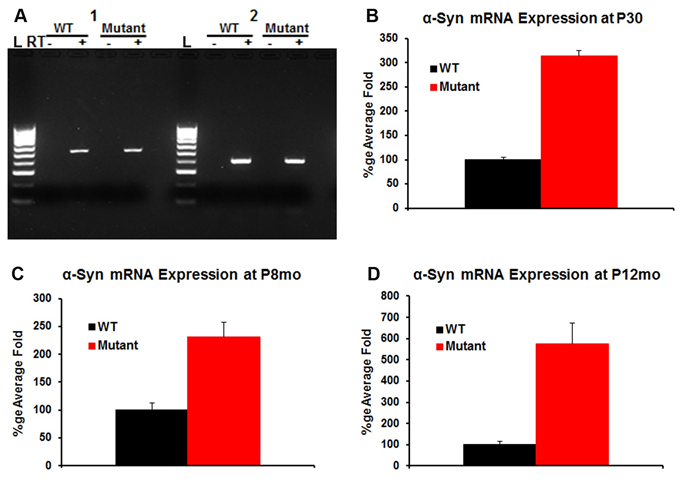
Figure 1: α-Syn mRNA in the WT and transgenic (mutant) cochlea: RT-PCR experiment was used to detect the presence of α-Syn mRNA in the WT and transgenic cochlea using two different set of primers 1 and 2 (Figure 1A). Controls lacking reverse-transcriptase (denoted with a ‘-‘) are included. The data demonstrate expression of α-Syn mRNA in the WT and transgenic cochlea (Figure 1A). Quantitative, real-time PCR was used to determine relative levels of α-Syn mRNA within the cochlea in the WT and the transgenic mice at P30, P8mo and P12mo. The expression levels were compared to the ribosomal protein L19 used as a house keeping gene. α-Syn mRNA shows ~3x amplification in the transgenic cochlea compared to WT at P30 (Figure 1B). α-Syn expression slightly decreased at P8mo (Figure 1C) then increase to reach ~5x amplification in the transgenic at P12mo (Figure 1D) when compared to the WT. Differences between the levels of α-Syn mRNA were statistically significant (n=5) for each sample, p<0.05.
Quantitative, real-time PCR (qPCR) was next used to assess the relative level of the α-Syn mRNA in the WT and transgenic cochlea at P30, P8mo and P12mo. The expression levels were compared to the ribosomal protein L19 used as a house keeping gene. α-Syn mRNA showed ~3x amplification in the transgenic cochlea compared to WT at P30 (Figure 1B). α-Syn expression slightly decreased at P8mo (Figure 1C) then increased to reach ~5x amplification in the transgenic at P12mo when compared to the WT (Figure 1D). Together these data suggest that α-Syn mRNA has significantly higher expression levels in transgenic mice cochlea compared to WT, documenting the correct α-Syn over-expression phenotype. Differences between the levels of α-Syn mRNA were statistically significant (n=5 for each sample, p<0.05). α-Syn Protein Expression: Anti-α-Syn antibody (red), which recognizes both human and mouse proteins, and anti-neurofilament 200 (NF200, green) were used for immunofluorescence within cochlear tissue (Figure 2). As expected, in the WT α-Syn localized predominantly to the base of the OHCs [23]. In contrast, in the transgenic mice α-Syn is additionally found throughout the inner and outer hair cells, as well as Deiters’ cells and inner phalangeal cells (Figure 2A). Western blot analysis was also performed to quantify α-Syn protein in the WT and the transgenic cochlea (Figures 2B & 2C). Similar to what was seen with immunofluorescence and qPCR, there was an increase of α-Syn protein in the transgenic cochlea as compared to the WT, but with a higher ratio of α-Syn protein expression than RNA expression at P30. In addition to providing further proof of the correct phenotype of the α-Syn over-expression mice, these data confirmed the specificity of the α-Syn antibody.
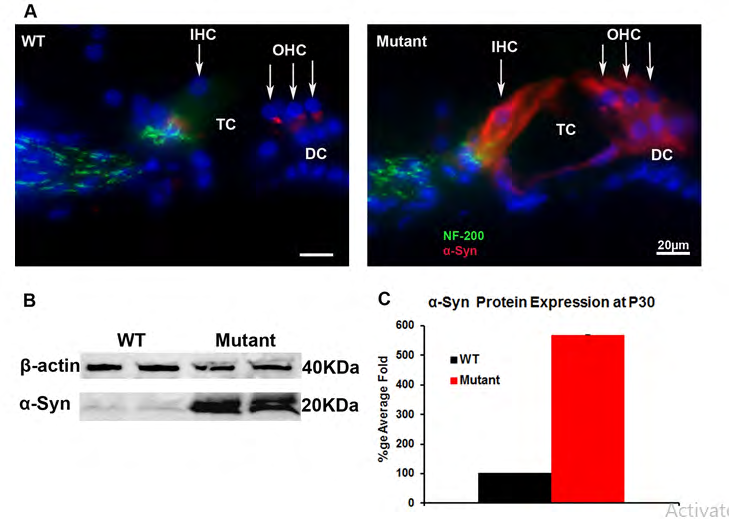
Figure 2: α-Syn protein expression in the WT and transgenic (mutant) mice cochlea: Immunofluorescence. was used to detect α-Syn and neurofilament protein expression using anti α-Syn antibody (Red) and anti-neurofilament 200 (NF200, green) antibodies (Figure 2A). The α-Syn protein localizes predominantly to the base of the cochlear OHCs and weak labeling to the base of the IHCs in WT. In transgenic mice over-expressing α-Syn, staining is seen in many cell types in the OC, including IHCs, OHCs, and Deiters’ cells (Figure 2A). This increase of the α-Syn protein in the transgenic was further demonstrated by western blot (Figure 2B) and quantified in Figure 2C relative to the protein β actin. OC: organ of Corti; DC Deiters’ cells; TC: Tunnel of Corti. OHC: outer hair cells; IHC: inner hair cells.
Effect of α-Syn Expression on Hearing: Auditory function in both WT and transgenic mice was tested to determine the effect of α-Syn over-expression on hearing.
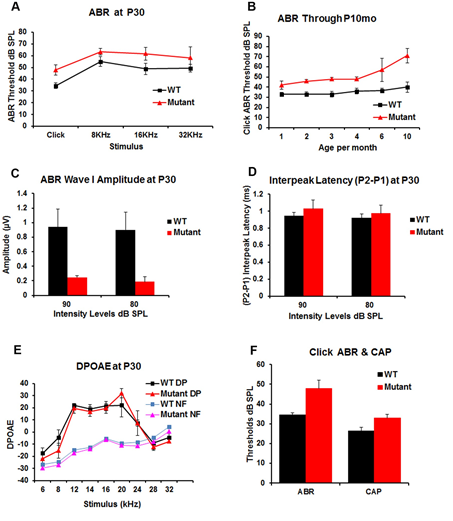
Figure 3: Auditory function was tested in WT and transgenic (mutant) mice to determine the effect of α-syn over-expression on hearing function. Acoustic brainstem response (ABR) testing (Figure 3A) at P30 demonstrates a statistically significant increase of ABR threshold of about 15dB in transgenic mice for click and pure tones (8, 16 and 32 KHz) stimuli; Figure 3B shows the changes in hearing in the transgenic and WT mice over time as measured by click ABR at P1-10months. This demonstrates an elevated ABR threshold (~10-20 dB) in the transgenic vs. WT littermates that remains relatively stable until P4mo then progressively elevate in the transgenic as compared to WT mice to reach ~30dB at P10mo. Figure 3C demonstrates a significant decrease in ABR wave I amplitudes in the transgenic mice measured at two different intensity levels (80, and 90 dB SPL) when compared with the WT littermates. Figure 3D, ABR P2–P1 interpeak latencies showed a non-significant increase at P30 in the transgenic when compared to the WT mice. Distortion product otoacoustic emissions (DPOAE) at P30 (Figure 3E) showed no differences between the WT and the transgenic mice. DP means (DPOAEs) and NF means (Noise Floor). Comparison of click ABR and CAP thresholds in the WT (n=4) and transgenic (n=6) mice. Which demonstrated threshold elevation is maintained in CAP measurements but has a lower magnitude (~8dB difference between WT and transgenic) than the one observed for ABR (Figure F).
Auditory brainstem response (ABR) was used to test hearing in the WT and transgenic mice at P30 using broadband clicks and tone-burst stimuli (8, 16, and 32 kHz) at varying ages (P1mo- P10mo). At P30 (Figure 3A), ABR demonstrated a statistically significant elevation of ABR thresholds (average= 15dB) in transgenic mice for both click and pure tone-burst stimuli in the transgenic compared to WT mice. This ABR threshold elevation is relatively stable until P4mo (Figure 3B), after which the ABR threshold difference progressively increases to a 30dB difference by P10mo. At P30 there was a significant decrease in the ABR wave I amplitude of the transgenic vs WT mice for click stimuli when measured at either 80dB or 90dB (Figure 3C). To analyze whether the ABR threshold increase in the transgenic mice also had an effect on ABR wave morphology, the ABR interpeak latency between P1 and P2 was measured at P30. The transgenic mice demonstrated no significant P2-P1 delays for click stimuli (Figure 3D). While the precise cause of the decrease in ABR amplitude is not known, we initially postulated that it was a likely a consequence of either the inhibition of OHC contraction and/or a reduction of glutamate release by the IHC due to α-Syn over-expression. To test this, OHC and efferent neuronal function were measured using distortion product otoacoustic emissions (DPOAE) at P30 (Figure 3E). No significant differences were noted between the WT and the transgenic mice.
To identify more precisely the location of the defect identified in sound-evoked potentials in the transgenic mice, auditory nerve compound action potentials (CAP) were recorded at the round window and compared with ABR recordings in the WT and transgenic mice (Figure 3F). Under these conditions, the same differences were observed between the ABR threshold in the WT and the transgenic mice, approximately 15dB. This threshold elevation was also seen in CAP measurements, but with a smaller 8dB difference between WT and transgenic mice. Thus, while α-Syn over- expression clearly causes an ABR threshold elevation, it is unclear whether this results from changes in IHC glutamate release, changes in CNS synaptic function or a combination of these factors.
Effect of α-Syn Expression on Susceptibility to Noise: Because α-Syn protein is expressed in the base of the OHCs, we aimed to examine the effects on the efferent system by examining susceptibility to noise-induced hearing loss in the transgenic mice. To test this hypothesis, WT and transgenic mice were exposed to 105 dB, 4-20 kHz octave band noise for 30 minutes. The ABR thresholds were measured before and after 72 hrs and 26 days following the noise-exposure (Figures 4A & 4B). At 72 hrs, WT mice had a significant threshold shift of ~33dB (Figure 4C) which remained at 26 days post exposure. In contrast, in mice over-expressing α-Syn, there was only a ~7 dB threshold shift with click stimuli (Figure 4C). After 26 days (Figure 4B), the ABR threshold levels in WT mice showed a small recovery but remained elevated by 10-20dB compared to the transgenic mice, whose thresholds did not significantly change over this time period. These data suggest a protective effect of α-Syn over-expression against a permanent threshold shift after loud noise exposure. These findings are in line with other studies that have documented decreased susceptibility to noise trauma with increased activity of the efferent auditory pathway [36, 37, 38].
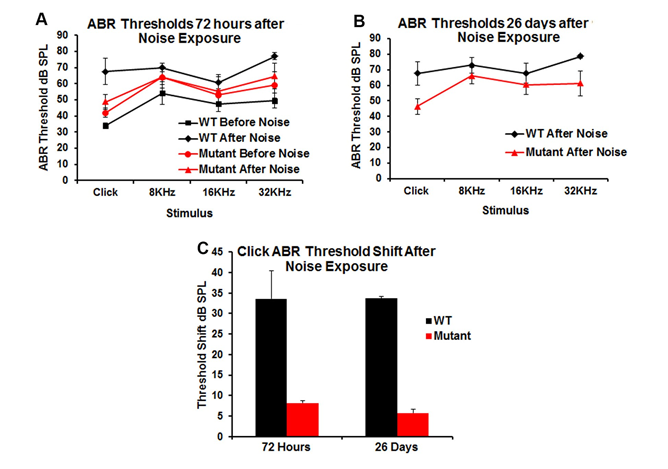
Figure 4: Changes in hearing threshold following noise exposure (105 dB, 4-20 kHz band, 30 minutes). Data are presented as mean ± SD. The threshold shift was statistically different in pre- vs post-noise exposure mice 72 hrs and 26 days following noise exposure for both WT and transgenic (mutant) mice (Figures 4A & 4B). However, WT mice showed a greater threshold shift (33dB) from baseline (before exposure) and when compared to transgenic mice after 72 hrs and 26 days post exposure (Figure 4C).
Histology
To determine whether there were anatomical correlates that might account for elevated ABR thresholds in the transgenic mice, we examined the morphology of the cochlea using both light and electron microscopy at P30. Standard histological analysis of toluidine blue stained sections of the organ of Corti and the spiral ganglion from the mid-turn of the WT and transgenic cochleae demonstrated no apparent histological differences; each mouse cochlea showed normal organ of Corti light microscopic morphology and spiral ganglion cell counts (Figure 5A). The cochlear whole-mount immunofluorescent staining using an anti- myosin 7a antibody, a marker for hair cells, demonstrated normal inner (IHC) and outer hair cell (OHCs) counts, without loss at any location along the cochlea (Figure 5B).
Additional labeling with rhodamine phalloidin, a stain for actin, also revealed that OHC stereocillia and cuticular plates were present and intact (Figure 5B) in both WT and transgenic mice. Transmission electron micrographs of the medial olivocochlear (MOC) efferent synapses (Figure 6A) demonstrated that the general appearance of MOC presynaptic efferent nerve terminals of the WT and transgenic mice were normal. Presynaptic nerve terminals of WT and transgenic mice did not show any noteable differences in vesicle shape or changes in the vesicle pool in the transgenic. We also noted normal IHC ribbon synapses in both transgenic and WT littermates, having normal appearing morphology with no obvious changes in the vesicle pool (Figure 6B).
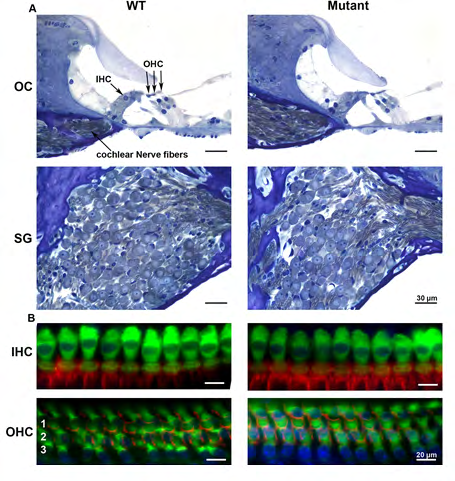
Figure 5: Light microscopy of the WT and transgenic (mutant) mice inner ear demonstrates no significant histological differences. Toluidine blue stain of the organ of Corti (OC) and spiral ganglion (SG) region from mid-turn of the WT and transgenic cochlea at P30 shows normal OC morphology and SG cell count (Figure 5A). Figure 5B shows cochlear whole-mount immunofluorescent staining using anti-myosin 7a antibody and rhodamine phalloidin staining. There is no significant difference between WT and transgenic hair cells at P30. Hair cell counts using representative samples of surface preparations show a normal complement of IHC and OHC and no IHC or OHC loss at all locations along the cochlea. Scale bar, 20µm. OHC rows 1–3 are indicated by 1, 2, and 3, respectively.
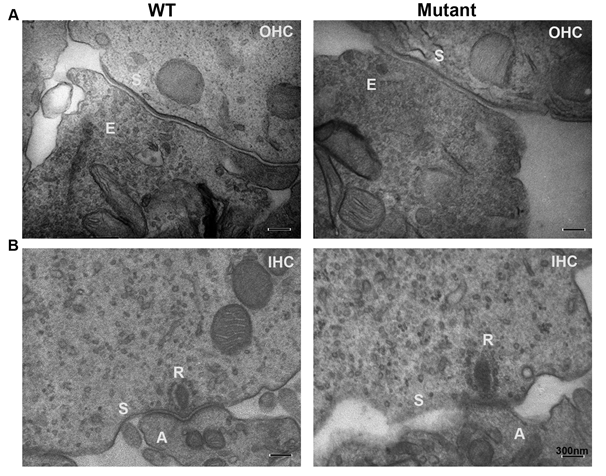
Figure 6: Transmission electron micrographs of medial olivocochlear (MOC) efferent synapses (Figure 6A) demonstrate that general appearance of MOC presynaptic efferent nerve terminals of the WT and transgenic (mutant) mice are normal with normal vesicles with any significant differences in vesicle shape or significant changes of the vesicle pool in the transgenic. We also noted normal IHCs ribbon synapse in the transgenic when compared to WT littermates. Transgenic’s IHC ribbon synapses showed normal appearing shape similar to the WT with no significant increase of vesicles pool (Figure 6B). OHCs efferent terminal labeled (E); synaptic membrane (S); Outer hair cells (OHC); Inner hair cells (IHC); afferent terminals (A); IHC ribbon synapse (R).
Discussion
Effects of α-Synuclein Over-Expression on Hearing and Cochlear Morphology
α-Syn has been implicated in the pathogenesis of Parkinson’s Disease as well as other neurodegenerative disorders. While the precise mechanism underlying this association is unknown, it is thought to be mediated through an effect on synaptic transmission. Many patients with PD also have hearing loss. However, because PD occurs primarily in the elderly, a population in whom a majority has age-related hearing loss, distinguishing hearing loss from presbycusis versus that induced by pathologic changes due to PD is challenging. Despite this confounding influence, there are studies that document an increase in hearing loss in patients with PD [39, 40]. One recent study documented high frequency hearing loss with elevated speech recognition thresholds and impaired speech discrimination, which correlated with more advanced disease stages in PD patients as compared to age-matched controls [39]. With α-Syn’s presumed role in Ca2+ homeostasis and effects on voltage- dependent Ca2+ influx [21, 22], and its localization mainly to the OHC synapse Akil O. et al. [23], it is not surprising that abnormalities in α-Syn may have a detrimental effect in the auditory system as well. However, previous studies in a α-Syn knockout mouse at P21 failed to document any changes in auditory morphology or physiology [24]. Contemporary theories suggest α-Syn over-expression is linked to PD Singleton [25, 26, 27], due to inhibited neurotransmitter release [30] which in turn leads to pathological changes in the spinal and brainstem motoneurons [7, 29]. We thus sought to determine if α-Syn over-expression in a α-Syn over- expressing transgenic mouse model leads to any pathologic changes in auditory morphology or physiology. The results of these studies document that over-expression of α-Syn in the cochlea leads to a significant elevation of the ABR threshold and a decrease in wave I amplitude, but no discernable effect on DPOAEs, no loss of inner or outer hair cells, spiral ganglion neurons, or alterations in the morphology of the afferent IHC (ribbon) or efferent OHC synapses. Further, over-expression of α-Syn confers protection against a permanent threshold shift from noise exposure.
α-Syn Over-Expression Protects Against Noise- Induced Hearing Loss
The two major questions arising from these findings are: why does over-expression of a-syn lead to hearing loss, and why does this confer protection against a permanent threshold shift from noise? With regard to the hearing loss in the transgenic mice, a possible mechanism may be found in decreased neurotransmitter release at the afferent inner hair cell ribbon synapse, in a manner similar to that described in the CNS [30]. Reduction in neurotransmitter release should lead to an elevation of the ABR threshold and a decrease in wave I amplitude, which was seen in our model. Further, there was also a reduction in the compound action potentials in the transgenic mouse, which supports this concept. Loss of hair cells or abnormalities in synapse morphology does not account for these changes, since they were normal in the transgenic mice. To definitively pinpoint elevation of ABR thresholds at the level of the synapse, electrophysiologic studies would be required. However, such studies would not likely answer how α-Syn over-expression leads to a reduction in glutamate release, resulting in the changes seen.
These studies also demonstrated that α-Syn over- expression leads to protection against a permanent threshold shift from noise exposure. This occurred in the setting of normal OHC numbers, normal hair cell morphology, and normal DPOAE measures. It seems likely that changes in the efferent auditory pathway are responsible for this finding. Firstly, α-synuclein localizes to the efferent synapse [23].
Additionally, it has been shown by a number of investigators that the efferent system mediates protection against noise-induced hearing loss for review see Fuente A, et al. [41]. In particular, it has been documented that the strength of the efferent reflex mediated by the olivocochlear system is proportional to the susceptibility to noise-induced hearing loss [36, 37], while genetic alterations of this pathway can lead to increased protection against noise trauma [36, 38]. In our current model, over-expression of α-Syn also provides protection against a permanent threshold shift after noise exposure, suggesting that the efferent system and olivocochlear reflex are strengthened. Whether this is a central phenomenon, due to increased olivocochlear synaptic activity arising from the brainstem, or a peripheral response, due to decreased OHC function due to changes at the efferent synapse, is not known. At the cochlear level, α-Syn clearly localizes to the OHC synaptic region. In prior studies by Taranda J, et al. [38], a point mutation in the OHC nicotinic cholinergic receptor (nAChR) led to an increase in receptor conductance and enhanced noise protection. This implies that α-Syn over-expression in the OHC and at the synapse may increase the amount of acetylcholine release at the OHC synapse or alter the post-synpatic response of the OHC. Thus, to explain both findings, the reduction in hearing (ABR threshold elevation) and the enhanced noise protection, we suggest that α-Syn over-expression reduces the afferent response while enhancing the efferent response. Again, some clarity may be provided by the findings from Taranda J, et al. [38]. In that study ABR and DPOAE thresholds were elevated in ‘gain of function’ nAChR α9 knock-in transgenic mice. One tested and confirmed hypothesis in the Taranda J, et al. [38] study was that ongoing acetylcholine release produced a much stronger inhibitory effect, so acoustic thresholds were higher due to that enhanced inhibition. Thus, if α-Syn overexpression enhances efferent release of acetylcholine at the efferent synapse, this would have an effect similar to that seen with the nAChR α9 gain of function knock-in mouse, decreasing acoustic thresholds, with no obvious change in morphology. This also fits with the protection from acoustic trauma as seen in both the nAChR α9 gain of function knock- in mouse and the α-synuclein over-expression mice.
What is the Role of α-Synuclein in Inner Hair Cells and Supporting Cells?
There remains the question of the effect of α-Syn’s presence in inner hair cells and supporting cells in the over- expression transgenic mice. However, given that α-Syn is not normally functional in these cells, it is likely that α -Syn’s presence in these cells in the transgenic mice is similarly non-functional and does not affect IHC neurotransmitter release. This is supported by three pieces of evidence:
- The IHC ribbon synapse differs in many ways from other neuronal synapses (e.g., different class of voltage-gated calcium channels, no synapsin, etc.);
- α-Syn does not localize to IHCs in the WT mouse; and
- In the α-Syn knockout mouse no functional deficit was seen [24].
Thus, all the changes seen in the α-Syn over- expression mice can be explained by increased efferent activity.
It should be noted that prior studies have also demonstrated that α-Syn over-expressing mice exhibit a number of abnormalities within the central nervous system, including pathological changes in the spinal and brainstem motoneurons [28, 29, 42, 43]. Thus, we cannot definitely rule out central pathology as a cause or contributor to the hearing changes observed in the transgenic mice. But the absence of pathological changes to the cochlear hair cells suggests that α-Syn over-expression affects the more the central auditory pathway than the cochlea. Future studies could potentially resolve the question of whether the abnormalities observed are peripherally vs. centrally mediated in this mouse model.
Conclusion
In summary, over-expression of α-Syn in the mouse cochlea leads to several interesting findings. Despite normal cochlear morphology, with normal hair cell and spiral ganglion counts and synaptic morphology, animals display an elevation in ABR thresholds, a decrease of wave I amplitude, and a reduction in compound action potentials, while also showing normal DPOAEs and enhanced protection against noise-induces hearing loss. These findings suggest that α-Syn over-expression leads to an enhancement of the efferent response, similar to mutations which enhance nAChR α9 function, supporting the hypothesis that ongoing acetylcholine release produced a much stronger inhibitory effect, so acoustic thresholds become elevated due to this enhanced inhibition. However, central pathology observed in this transgenic mouse cannot be ruled out as a cause or contributor to the hearing changes observed. Further studies are necessary to determine the exact mechanism of function of the α-Syn over-expression on the hearing in this mouse model and could potentially resolve the question of whether these abnormalities observed are mediated peripherally vs. centrally (or both) in this mouse model.
Conflict of Interest
The authors declare no competing financial interests.
Acknowledgement
We would like to acknowledge Hearing Research Incorporated (HRI) for their generous financial support and Dr. Robert Edwards of the University of California, San Francisco, and Department of Neurology for generously providing the alpha-synuclein overexpressing transgenic mice for this study.
References
-
Lavedan C (1998) The Synuclein Family. Gen Res 8(9): 871-880.
-
Biere AL, Wood SJ, Wypych J, Steavenson S, Jiang Y, Et Al. (2000) Parkinson’s Disease-Associated Α-Synuclein Is More Fibriollogenic Than Β- And Γ-Synuclein And Cannot Cross-Seed Its Homologs. J Biol Chem 275(44): 34574-34579.
-
Maroteaux L CJ, Scheller RH (1988) Synuclein: A Neuron-Specific Protein Localized To The Nucleus And Presynaptic Nerve Terminal. J Neurosci 8(8): 2804-2815.
-
Jakes R, Spillantin MG, Goedert M (1994) Identification Of Two Distinct Synucleins From Human Brain. FEBS Lett 345(1): 27-32.
-
Nakajo S, Aiuchi T, Shibayama T, Okahashi I, Ochiai H, et al. (1990) Purification And Characterization Of A Novel Brain-Specific 14-Kda Protein. J Neurochem 55(6): 2031-2038.
-
Surguchov ASI, Solessio E, Baehr W (1999) Synoretin-A New Protein Belonging to the Synuclein Family. Mol Cell Neurosci 13(2): 95-103.
-
Giasson BI DJ, Forman MS, Lee VM, Trojanowski JQ (2001) Prominent Perikaryal Expression of Alpha- and Beta-Synuclein in Neurons of Dorsal Root Ganglion and in Medullary Neurons. Exp Neurol 172(2): 354-362.
-
Murphy DD, Rueter SM, Trojanowski JQ, Lee VM (2000) Synucleins Are Developmentally Expressed, And Alpha- Synuclein Regulates The Size Of The Presynaptic Vesicular Pool In Primary Hippocampal Neurons. J Neurosci 20(9): 3214-3220.
-
Saha AR NN, Hanger DP, Anderton BH, Davies AM, Buchman VL, et al. (2000) Induction Of Neuronal Death By Alpha-Synuclein. Eur J Neurosci 12(8): 3073-3077.
-
Tofaris GK, Spillantini MG (2007) Physiological And Pathological Properties of Alpha-Synuclein. Cell Mol Life Sci 64(17): 2194-2201.
-
Baba MN, Tomita T, Nakaya K, Lee VM, Tojanowski JQ, et al. (1998) Aggregation Of Α-Synuclein In Lewy Bodies Of Sporadic Parkinson’s Disease And Dementia With Lewy Bodies. Am J Pathol 152(4): 879-884.
-
Galvin JE UK, Lee VM, Trojanowski JQ (1999) Axon Pathology In Parkinson’s Disease And Lewy Body Dementia Hippocampus Contains Alpha-, Beta-, And Gamma-Synuclein. Proc Natl Acad Sci USA 96(23): 13450-13455.
-
Galvin JE GB, Hurtig HI, Lee VMY, Trojanowski JQ (2000) Neruodegeneration With Brain Iron Accumulation Type 1 Is Characerized By Α-, Β- And Γ-Synuclein Neuropathology. Am J Pathol 157(2): 361-368.
-
Ueda K FH, Masliah E, Xia Y, Iwai A, Yoshimoto M, et al. (1993) Molecular Cloning Of Cdna Encoding An Unrecognized Component of Amyloid in Alzheimer Disease. Proc Natl Acad Sci USA 90(23): 11282-11286.
-
Ueda K ST, Mori H (1994) Tissue-Dependent Alternative Splicing Of Messengeγ-RNA For MACP, The Precursor Of Non-Α-B Component Of Alzheimer’s Disease Amyloid. Biochem Biophys Res Commun 205(2): 1366-1372
-
Quilty MCGW, Pountney DL, West AK, Vickers JC (2003) Localization of A-, B-, and R-Synuclein During Neuronal Development and Alterations Associated with the Neuronal Response to Axonal Trauma. Exp Neurol 182: 195-207.
-
Gureviciene I, Gurevicius K, Tanila H (2007) Role Of Alpha-Synuclein In Synaptic Glutamate Release. Neurobiol Dis 28(1): 83-89.
-
Cabin DE, Shimazu K, Murphy D, Cole NB, Gottschalk W, et al. (2002b) Synaptic Vesicle Depletion Correlates With Attenuated Synaptic Responses To Prolonged Repetitive Stimulation In Mice Lacking Alpha-Synuclein. J Neurosci 22(20): 8797-8807.
-
Eybalin M (1993) Neurotransmitters And Neuromodulators Of The Mammalian Cochlea. Physiol Rev 73(2): 309-373.
-
Puel JL (1995) Chemical Synaptic Transmission In The Cochlea. Prog Neurobiol 47(6): 449-476.
-
Ueda K, Shinohara S, Yagami T, Asakura K, Kawasaki K (1997) Amyloid Beta Protein Potentiates Ca2+ Influx Through L-Type Voltage-Sensitive Ca2+ Channels: A Possible Involvement Of Free Radicals. J Neurochem 68(1): 265-271.
-
Adamczyk A, Strosznajder JB (2006) Alpha-Synuclein Potentiates Ca2+ Influx Through Voltage Dependent Ca2+ Channels. Neuroreport 17(8): 1883-1886.
-
Akil O, Weber C, Park S, Ninkina N, Buchman V, et al. (2008) Localization Of Synculeins In The Mammalian Cochlea. J Assoc Res Otolaryngol 9(4): 452-463.
-
Park SN, Akil O, Weber C, Lustig LR (2009) Early Onset Hearing Loss And Efferent Auditory Deficits in Mice Lacking Α-Synuclein. In: Abstract Book of the Thirty- Second Annual Midwinter Research Meeting, pp: 13.
-
Singleton AB, Farrer M, Johnson J, Singleton A, Hague S, et al. (2003) Alpha Synuclein Locus Triplication Causes Parkinson’s Disease. Science 302(5646): 841.
-
Chartier Harlin MC, Kachergus J, Roumier C, Mouroux V, Douay X, et al. (2004) Alpha-Synuclein Locus Duplication As A Cause Of Familial Parkinson’s Disease. Lancet 364(9440): 1167-1169.
-
Maraganore DM, De Andrade M, Elbaz A, Farrer MJ, Ioannidis JP, et al. (2006) Collaborative Analysis Of Alpha- Synuclein Gene Promoter Variability And Parkinson Disease. JAMA 296(6): 661-670.
-
Giasson BI, Duda JE, Quinn SM, Zhang B, Trojanowski JQ, et al. (2002) Neuronal Alpha-Synucleinopathy With Severe Movement Disorder In Mice Expressing A53T Human Alpha-Synuclein Neuron 34(4): 521-533.
-
van Putten HD, Wiederhold KH, Probst A, Barbieri S, Mistl C, et al. (2000) Neuropathology In Mice Expressing Human Alpha-Synuclein. J Neurosci 20(16): 6021-6029.
-
Nemani VM, Lu W, Berge V, Nakamura K, Onoa B, et al. (2010) Increased Expression Of Alpha-Synuclein Reduces Neurotransmitter Release By Inhibiting Synaptic Vesicle Reclustering After Endocytosis. Neuron 65(1): 66-79.
-
Akil O, Chang J, Hiel H, Kong JH, Yi E, et al. (2006) Progressive Deafness And Altered Cochlear Innervation In Knockout Mice Lacking Prosaposin. J Neurosci 26(50): 13076-13088.
-
Seal RP, Akil O, Yi E, Weber CM, Grant L (2008) Sensorineural Deafness And Seizures In Mice Lacking Vesicular Glutamate Transporter 3. Neuron 57(2): 263- 275.
-
Akil O, Rouse SL, Chan DK, Lustig LR (2015) Surgical Method For Virally Mediated Gene Delivery To The Mouse Inner Ear Through The Round Window Membrane. J Vis Exp 16(97): 52187.
-
Livak KJ, Schmittgen TD (2001) Analysis Of Relative Gene Expression Data Using Real-Time Quantitative PCR And The 2(-Delta Delta C (T)) Method. Methods 25(4): 402-408.
-
Berglund AM, Ryugo DK (1991) Neurofilament Antibodies and Spiral Ganglion Neurons of the Mammalian Cochlea. J Comp Neurol 306(3): 393-408.
-
Maison SF, Liberman CM (2000) Predicting Vulnerability To Acoustic Injury With A Noninvasive Assay Of Olivocochlear Reflex Strength. J Neurosci 20(12): 4701- 4707.
-
Luebke AE, Foster PK (2002) Variation In Inter-Animal Susceptibility To Noise Damage Is Associated With Alpha 9 Acetylcholine Receptor Subunit Expression Level. J Neurosci 22(10): 4241-4247.
-
Taranda J, Maison SF, Ballestero JA, Katz E, Savino J, et al. (2009) A Point Mutation In The Hair Cell Nicotinic Cholinergic Receptor Prolongs Cochlear Inhibition And Enhances Noise Protection. Plos Biol 7(1): 18.
-
Pisani V, Sisto R, Moleti A, Di Mauro R, Pisani A, et al. (2015) An Investigation Of Hearing Impairment In De- Novo Parkinson’s Disease Patients: A Preliminary Study. Parkinsonism Relat Disord 21(8): 987-991.
-
Lai SW, Liao KF, Lin CL, Lin CC, Sung FC, et al. (2014) Hearing Loss May Be A Non-Motor Feature Of Parkinson’s Disease In Older People In Taiwan. Eur J Neurol 21(5): 752-757.
-
Fuente A (2015) The Olivocochlear System And Protection From Acoustic Trauma: A Mini Literature Review. Front Syst Neurosci 9: 94.
-
Maison SF, Luebke AE, Liberman MC, Zuo J (2002) Efferent Protection From Acoustic Injury Is Mediated Via Alpha9 Nicotinic Acetylcholine Receptors On Outer Hair Cells. J Neurosci 22(24): 10838-10846.
-
Lee MK, W Stirling, Xu Y, Xu X, Qui D, et al. (2002) Human Alpha-Synuclein-Harboring Familial Parkinson’s Disease-Linked Ala-53→Thr Mutation Causes Neurodegenerative Disease With Alpha-Synuclein Aggregation In Transgenic Mice. Proc Natl Acad Sci USA 99(13): 8968-8973.
- 4th Branchial Cleft Sinus Anomaly Presenting as Recurrent Thyroid Abscess in A Child: A Case Report
- Parotid Duct Injury Repaired Using an Angiocatheter Stent: A Case Report
- Organization and Functionality of the Referral and Counter-Referral System for ENT Disorders in District Hospitals of N’Djamena, Chad: A Cross-Sectional Analytical Study
- Facial Metastases from a Gastrointestinal Stromal Tumor: A Case Report
- Panorama of Ent Cancers and Literature Review: Epidemiological Profile and Therapeutic Management
- Could Antimicrobial Resistance Prove to Be Both a Threat and an Opportunity for us?