The Holobiot and the Possibilities for Intervention in the Interspecies Interactions of the Microbiotа
Introduction: Throughout our recorded history, we have repeatedly encountered pandemics with high mortality rates. Climate change, environmental changes, population mobility, uneven settlement, unequal sanitation, changes in animal habitats and rapidly increasing antibiotic resistance have also put increased pressure on the host-pathogen-environment system.Objectives: Viral pathogens facilitate the colonization of other pathogens by altering the diversity and functions of the microbiota. Studying the interspecies interactions between them, the components of the microbiota and the host, as well as tracking horizontal gene transfer, will help us to clarify the reasons for the different course of the same disease in patients. It will clarify the timing and scope of our interventions and how to improve their effect in each patient.Results: The concept of the holobiont considers the host and its taxonomically and ecologically related microbial communities as a whole. This will serve basis to understand how this complex relatively static and at the same time dynamically developing ecosystem adapts to continuous environmental challenges.Conclusion: Knowledge the relationships in the holobiont will help us avoid superficial handling of them. Even with their seemingly correct use, there is a risk of unintentionally triggering a cascade of negative relationships during our interventions, which leads to serious risks to the health of patients. Our task is to find the right time, type and volume of pharmacological, non-pharmacological and dietary interventions. When treating one of the components of the microbiota, we indirectly affect the others, which should be included in our therapeutic plan.
Abbreviations
ACE2: Аngiotensin Converting Enzyme 2; AIP: Autoinducing Peptide; AMP: Аntimicrobial Peptide; AMR: Аntimicrobial Resistance; АdV: Adenovirus; CoV: Corona Viruses; DАО: Diaminooxidase; DC: Dentritic Cells; DOL: Division of Labor; DSF: Diffusible Signal Factors; GIT: Gastro Intestinal Tract; GТDВ: Genome Taxonomy Database; HВV: Hepatitis В Virus; HCV: Hepatitis C Virus; HDV: Hepatitis D Virus; HE: Hemagglutinin Esterase; HERV: Human Endogenous Retroviruses; HMPV: Human Metapneumovirus; HNMT: Histamine N-Methyltransferase; HPV: Human Papilloma Virus; HRV: Human Rhinovirus; IAV: Influenza A Virus; ICIs: Immune Checkpoint Inhibitors; IFN: Interferon α/β and γ; IL: Interleukin; INKT: Invariant Natural Killer T Cells; IPI: Invasive Pneumococcal Infection; ISGˋs: Interferon Stimulated Genes;
LAB: Lactic Acid Bacteria; LEV: Live Enterovirus Vaccine; МС: Mast Cells; MDCK: Madin-Darby Canine Kidney Cells; MERS- CoV: Middle East Respiratory Syndrome Coronavirus; MHC-I: Мajor Histocompatibility Complex Class I; MHC-II: Мajor Histocompatibility Complex Class II; MPO: Мieloperoxidasa; NAAT: Nucleic acid Amplification Test; NETs: Neutrophil Extracellular Traps; NF-kB: Nuclear Factor Kappa-Light- Chain-Enhancer Of Activated B Cells; NLRP3: NLR Family Pyrin Domain Containing 3; OUT: Оperational Taxonomic Unit; OS: Overall Survival; PAF: Platelet-Activating Factor; PCV: Pneumococcal Conjugate Vaccine; PFS: Рrogression-Free Survival; PIV – Para Influenza Virus; PPSV – Pneumococcal Polysaccharide Vaccine; PspA: Surface Protein Antigen; PRB: Predation-Resistant Bacteria; PRRs: Pathogen Recognition Receptors; QS: Quorum Sensing; ROS: Reactive Oxygen Species; RSV: Respiratory Syncytial Virus; SARS-CoV: Severe Acute Respiratory Syndrome Coronavirus; SIDS: Sudden Infant Death Syndrome; TJ: Tight Junctions; WGS: Whole Genomic Sequencing.
Introduction
Throughout recorded history, we have faced pandemics caused primarily by RNA viral pathogens, with high mortality rates [1, 2, 3, 4, 5, 6, 7]. From the bacterial ones, only the second plague pandemic (1346-1350) caused by Yersinia pestis was more deadly, killing 30-50% of the population of Afro-Eurasia [8, 9]. Climate and environmental changes, population mobility, uneven settlement, unequal sanitary conditions, changes in animal habitats, as well as rapidly increasing antibiotic resistance, lead to increased pressure on the host- pathogen-environment system. This creates the opportunity for highly pathogenic viruses of zoonotic origin to cross the interspecies barrier and stabilize in humans. Infectious diseases have represented, represent and will probably represent for many years to come a serious challenge for us and not only for us clinicians, despite the rapid development of medicine. During the treatment we often forget that we are not only treating the host (our body) but also the microbial communities associated with it.
Over the four billion years since the origin of life on earth, 10130 protein molecules, 10700 chains and 101000 metabolic pathways have evolved. They are the basis of interspecific interactions between microbiota components, viral and bacterial pathogens and the host. Knowledge of the evolutionary changes that organisms have undergone before mammals and humans appeared will help us unravel the possibilities for the course of infectious diseases and improve the effect of our interventions. Information of the protective mechanisms, virulence factors and positive interactions that have evolved over billions of years will allow us to shed light on the ways in which they can bypass (remain unnoticed) for our immune system and to explain the different course of the same disease in patients. An example of this is the ability of some bacteria to be resistant to predation (PRB) by unicellular eukaryotes. Packaged in vesicles or fecal pellets, they have gained an evolutionary advantage for survival in nutrient-poor environments, are more infectious than free- living bacteria and possess increased resistance to antibiotics. Packaged bacteria are a form of pathogenic bacteria that cooperate with protozoa. Exocytosis is a process by which protozoa can release hundreds of vesicles containing bacteria from a single cell [10, 11]. Vesicles range in size from 2 to 6 µm in diameter [12, 13]. When inhaled particles are smaller than 3.5 µm, they can penetrate the alveoli and part of them are with this size. When the vesicles/pellets are aerosolized, they can be transmitted over long distances. The aerosol route of transmission is a major route of transmission for many human pathogens. It is estimated that up to 50% of lower respiratory tract infections of unknown etiology are transmitted in this way [14, 15, 16, 17, 18, 19, 20, 21, 22].
Understanding the interactions between the host and its associated microbial communities, and their central role in host biology, ecology, and evolution is essential for us. Individual phenotypes should be viewed as expressions of the relationships between the host and associated microbial genomes. Symbiosis between species is at the heart of the concept of the holobiont. It was formulated by Lynn Margulis in 1991 and initially considered a simple biological unit - a host and a single hereditary symbiont. Later, the holobiont was viewed as a single entity between the host and its associated taxonomically and ecologically diverse communities of microorganisms or the microbiota. Within this unit, interactions ranging from synergistic to antagonistic occur. In Rosenberg E, et al. [19] introduced the term hologenome, which represents the sum of the host genome and the genomes of the associated microbial communities or the collective genome of the holobiont. The human genome contains about 20,000 genes, but our hologenome contains >33 million genes [15, 16, 17, 18, 19, 20, 21, 22]. Understanding the causes of rapidly increasing antibiotic resistance has also expanded [23, 24]. In respiratory diseases, our area of interest is the upper and lower respiratory tracts, but in light of the holobiont concept, it is not correct to consider the interactions in the different ecological niches of the human body separately.
The human microbiota is a natural community in which microorganisms are in constant contact and natural selection governs its dynamics. Bacterial communities must meet the following criteria: stability, consistency, reproducibility under different conditions and metabolic activity. According to the “Black Queen” hypothesis, during the evolution of natural communities, positive interactions in complex habitats are strengthened by gene loss, leading to dependencies between interacting microorganisms [25, 26]. Over time, interactions between strains become so complex that individual strains cannot thrive on their own. The theory suggests that dependencies between microorganisms, especially those of a nutritional nature, not only improve their vitality, but also strengthen their bonds against competitors, cheaters and stress. Analyses show that metabolic dependencies are major drivers of species coexistence in nature [27]. Sharing primary metabolites is crucial for positive interactions. According to D‘Sousa, cross-feeding is extremely important. Microorganisms use two ways to obtain these metabolites. The first is by sharing hydrolytic extracellular enzymes such as invertases, lipases and proteases, which directly degrade the available substrates from the total pool of molecules. In the second, we observe cross-feeding, in which one microorganism/donor converts a primary substrate into a product that is subsequently used by another/acceptor [28].
Microbiota Composition
Four kingdoms - archaea, viruses, bacteria and fungi mainly fоrm up the human microbiota and in part of the population parasites. It performs trophic, metabolic and protective functions. The metabolic activity of the intestinal microbiota(predominant) is expressed in the regulation of positive or negative metabolic homeostasis, the formation of an anaerobic environment that suppresses the virulence of pathogens and induces immune tolerance. Its metabolites are products of the metabolic pathways of tryptophan (Trp), histidine (His) and phenylalanine (Phe). The protective function is expressed in the production of bacteriocins that inhibit the growth or survival of the pathogens and the trophic one is expressed in the mobilization of bacteriophages that attack specific bacterial strains, minimizing their impact on the commensal microbiota. The colonization of the host occurs in waves during ontogenesis and through various mechanisms enters into constant contact with it.
Archaea
The archaeome is the least studied component of the human microbiota. Archaea inhabit the skin, gastrointestinal, urogenital and respiratory tracts. The complexity of their study is due to their similarities and differences with bacteria and eukaryotes, and at the same time, the possession of a combination of their specific features. It is estimated that they represent 1.2% of the human microbiota. Their diversity, like other components of the microbiota, depends on ethnicity, diet, habitat and age. Most gut archaea metabolize H2 and CO2, and release methane during exhalation and flatulence. Therefore, they are called methanogens. Their abundance suggests two types of people – those emitting significant amounts of methane and those with about 1000 times less gut archaea. Methanogens are an important participant in the gut community. On the one hand, bacteria produce H2, a key substrate for the growth of archaea, and they support the growth and metabolism of their bacterial “partners” and thus participate in shaping the phenotype. The production of adhesins and the formation of biofilms facilitate their ability to survive in the host, which in turn uses various strategies to control them. Archaea-bacteria-host interactions are increasingly associated with human health, but are still in the early stages of study due to the availability of only indirect evidence for this. Alterations in the composition of the archaeome have been associated with inflammatory bowel disease (IBD), colorectal carcinoma (CRC), periodontitis and vaginosis. Methane can slow intestinal motility, causing constipation and is often associated with irritable bowel syndrome (IBS). Studies on animal models indicate that аrchaea can probably activate human immune cells and methane can provoke increased production of anti- inflammatory molecules, and promote reduced production of pro-inflammatory ones. So far, there is no evidence that they can independently cause disease in humans.
Virobiota
The virobiota is a poorly understood component of the microbiota. Its main components are prokaryotic (bacteriophages), eukaryotic, and human-specific viruses (human endogenous retroviruses - HERVs). [29, 30]. Eukaryotic viruses infect human cells, intestinal fungi and parasites. They are mainly RNA viruses Prokaryotic viruses infect bacterial cells. Bacteriophages were discovered and described by Twort in 1915. They are single-stranded(ss) and double-stranded(ds) DNA and RNA, with dsDNA phages predominating. They constitute about 90% of the intestinal virobiota. Phage infecting bacteria are about 10 times more numerous. Antibiotic treatment provokes increased interactions between phages and bacteria [31, 32, 33]. By interacting with symbiotic bacteria, phages colonizing the human body shape the structural and functional composition of bacterial communities in the different anatomical zones of the human body. This occurs by lysing bacteria and generating new phage particles or by integrating the phage genome into the bacterial one. This leads to the emergence of new phages, changes in bacterial vitality and phenotype (host bacteria), transmission of genes of antibiotic resistance and virulence, changes in the ability of bacteria to produce toxins, increased tolerance to oxidative and acid stress, and improved energy utilization.
To understand the interactions in the microbiota, it is necessary to know the composition and structure of the cytoskeleton in each ecological niche. This is important because changes in mucosal immunity have not only local but also systemic effects in distant areas of the human body. The intestinal barrier plays a critical role in our health, maintaining the tolerance of the intestinal microbiota, participating in the absorption of nutrients and protecting against pathogenic invasion. It is composed by mucus layer, epithelium and lamina propria. Mucus is a gel-like layer with variations in thickness, composed mainly of glycoproteins or mucins secreted by goblet cells. Beneath it is the epithelial layer, which includes intestinal epithelial cells, immune cells, microbiota and metabolites. Intestinal cells include: enterocytes, goblet cells, enteroendocrine cells, M cells and Paneth cells. They are connected by tight junctions (TJs), which regulate paracellular transport. Reduced expression of TJ proteins (ocсludin, claudin, JAMs and ZO-1) results in increased intestinal permeability. It is also the reason for bacteria and their components - lipopolysaccharides (LPS), antigens and toxins to enter the lamina propria and trigger an inflammatory response. Intestinal cells express specific pathogen recognition receptors (PRRs) on their surface: TLRs (Toll-like receptors), C-type lectin phage receptors, RLRs-retinoic acid inducible gene (RIG)-I-like receptors and NLRs-nucleoide binding oligomerization domain (NOD) like receptors. They control the composition of the intestinal microbiota - they distinguish pathogens from non-pathogens. These cells have effector functions - Paneth cells secrete antimicrobial peptides, M cells are responsible for antigen uptake, phago- and transcytosis, and goblet cells secrete mucus. The epithelium has various lymphoid accumulations, among which the most important are Peyer’s patches, composed of B-cell follicles and T-cell areas. They participate in the adaptive immune response, mucosal immunity and are the main source of IgA, an effector molecule of adaptive immunity. Beneath the epithelial layer is the lamina propria, rich in immune cells - macrophages, dendritic cells and T, and B cells. The balance between anti- inflammatory (Tregs) and pro-inflammatory T cells (Th1, Th2 and Th3/Th17) plays a major role in the homeostasis of the intestinal mucosa [34, 35, 36, 37, 38, 39, 40]. By interacting with host cells, phages induce inflammatory and antiviral immune responses through activation of virus-discern receptors, secretion of pro-inflammatory cytokines and activation of adaptive immune responses without the involvement of host bacteria. Phages also participate in the regulation of anti-inflammatory mechanisms of the immune system, on the one hand by eliminating bacterial pathogens and on the other hand by directly interacting with pro-inflammatory cytokines, they reduce the overproduction of reactive oxygen species (ROS). They can also directly suppress bacterial growth by binding to intestinal mucosal cells [41, 42, 43, 44, 45]. Phago specific secretory IgA is a key regulatory factor limiting phage activity [46]. Immune regulation against viruses is modulated primarily by the intestinal microbiota.
Human endogenous viruses (HERVs) have integrated into the human genome and represent about 8% of it. They are transmitted from generation to generation through vertical transfer. Until recently, they were considered „junk“ or DNA without biological function. As a result of accumulated mutations over tens of millions of years, the majority of them are „defective“ or non-pathogenic, but there are also those that have the potential to assemble and trigger the immune response through pathogen recognition receptors (PRRs) or „viral switches“. Transcriptional activation of HERVs can induce insertional mutagenesis and chromosomal rearrangements, leading to altered cellular gene expression. In addition to regulatory processes(immunity - participation in the interferon response, brain and placenta development), their DNA sequences are also associated with regulatory proteins in cancer cells (21 types of cancer) [47, 48, 49, 50].
Benign viral load was observed in healthy controls. Resident viruses (phages and HERVs) are a powerful regulator of immunity, especially intestinal, by sending constant stimuli to the immune system, but without the manifestation of symptoms. They maintain the balance between homeostasis and inflammation in the human body. The number of bacteriophages in the human intestine is estimated at 1015. Most enteric viruses (mainly phages) are strictly dependent on their hosts and are difficult to culture [51]. The commensals Firmicutes and Proteobacteria in the small intestine and Bacteroides in the colon are responsible for maintaining mucosal integrity and immune tolerance. The former two genera have been shown to harbor the majority of lysogenic phages, which in turn prevent their infection by other lytic and lysogenic phages through the phenomenon of superimmunity, in which preexisting infection protects against reinfection or infection by closely related viruses [52]. Eukaryotic viruses and bacteriophages interact with mucosal immune cells. Viral genomes are recognized by intracellular and cytosolic receptors in epithelial and innate immune cells and these are viral RNA - TLR3, TLR7 and TLR8, NLRs and RLRs and DNA receptors - endoplasmic TLR9 and the cytoplasmic cGAS-STING pathway. Activation of the receptors triggers the production of NF-κB, IRF3 and IRF7 inducing the production of antiviral mediators such as type I IFN, cytokines (IL-1 and IL-6) and chemokines (CXCL8 and CXCL10). The action of these mediators on epithelial and immune cells promotes the formation of an antiviral environment that prevents pathogenic viral colonization [53].
Rapid colonization of the human body begins immediately after birth through contact with the mother‘s skin, milk and vaginal mucosa. The gut of infants is free of microorganisms at birth, but within a few hours there is an increase in microbial diversity. During the first 4 days, there is a poor bacteriome and a rich phageome, while at 2 years of age the bacteriome is rich and diverse, and the phageome is poorer [54]. The phages that initially colonize the gut are induced phages, originating from the bacteria that first colonize the baby. For example, bifidobacteria transmitted through breast milk contain prophages known as bifidophages [55]. Early in life, the abundance of temperate phages is high because the biomass of bacterial species is low or there is a shortage of potential hosts. As the infant grows, along with the expansion of bacterial species in the different niches of the gut, there is also increased colonization by virulent phages from the crAss-like and Microviridae families, which stabilize in early childhood. In adults, the virome is already abundant and highly resistant and consists mainly of the aforementioned families. These families are considered specific to human hosts, with only some being shared between individuals - 50% of the phageome is unique to each individual [56]. The synchronous colonization of the human body by viruses and bacteria is due to their close interactions. The composition of the gut virome is very dynamic.
Interactions between phages and bacteria are positive - inactive/dormant lysogenic phage provides an evolutionary advantage in replication and survival of the host and negative - predatory association (lytic phages). Predatory association is highly specific because it selectively infects only a certain host ignoring other bacteria. Phages are temperate and lytic or non-temperate. The former prevail and are characterized by a lysogenic life cycle in which they integrate into the chromosomes of bacterial hosts as extrachromosomal episomes or prophages. The latter can target bacterial “prey” by recognizing specific receptors on the membrane and through lysis, can penetrate layer by layer into the bacterial biofilm and destroy it by their depolymerases, exopolymer degrading and endolytic enzymes [57]. Phage predation and lysogenic transformation of bacterial cells play a key role in horizontal gene transfer and the regulation of bacterial abundance. Phage insertion can either alter bacterial genes and suppress their functions or encode genes that enhance the ability of host bacteria to expand their ecological niche. The transduction of genes responsible for toxin production (Shiga toxin) and antibiotic resistance by phages has been well studied [58, 59]. Genes encoding prophages can cause lysis of related strains and reduce competition (kill relatives). Prophages also have the ability to be transmitted to the host‘s offspring through vertical transmission [60].
Adverse conditions can induce prophages to enter a lytic replication cycle. The mechanism underlying prophage induction is due to DNA damage, which destabilizes the repressors of prophage induction. The rate of phage induction is regulated by a signaling pathway called quorum sensing (QS). The effect of prophage induction can be beneficial or detrimental depending on whether it is activated in pathogenic or commensal bacteria. Their induction can create a huge burden on the bacterial host and the potential to alter the composition of the gut microbiota [57]. Another type of association is mutualism - virions carry lysogens (idiot genes) that have no role in the life cycle of the virus but provide benefits to the bacterial host. Most intestinal bacteria are colonized by more than one temperate phage.
On the one hand, temperate phages help the bacterial host to adapt due to the expression of newly acquired phenotypes -resistance to other phage superinfections, resistance to antibiotics, increased genomic complexity, evolution of new pathogenic forms and tolerance to stresses, while on the other hand, lysogenic phages, by integrating their genome with that of the bacterial chromosome, avoid recognition and elimination by macrophages [61].
The diversity and composition of the phageome are determined by diet, age, diseases, therapies and environment [62, 63]. Diet is the most significant differentiating factor for the gut virome. Identical diets across individuals have the potential to make the microbiota more similar in phage composition, but not identical [55].
Bacteriota
The bacterial component of the microbiota evolves simultaneously with its human host, as organisms constantly move through the commensal-pathogen continuum. Its composition and abundance can be regulated by phages. The upper respiratory tract microbiota of healthy individuals contains several genera. During the first year of life, the genera Staphylococcus, Streptococcus, Corynebacterium, Moraxella, Haemophilus and Alloicoccus/Dolosigranulum predominate. After the third year Staphylococcus, Streptococcus, Corynebacterium, Prevotella, Veillonella, Propionibacterium and Fusobacterium are most commonly found. Corynebacterium, Dolosigranulum, Streptococcus epidermidis, and Staphylococcus lugdunensis are primarily responsible for reducing the incidence of disease caused by S. pneumoniae, H. influenzae, S. aureus and M. catharralis. The gut microbiome is predominantly anaerobic and the predominant genera are Firmicutes, Proteobacteria, Bacteroides and Ruminococcus [64, 65, 66, 67, 68, 69, 70].
The bacterial component consists of transient and permanent members. Transients are opportunistic pathogens responsible for infectious diseases and most commercially available probiotics. The permanent members are most of the non-pathogenic bacteria - colonizers or commensals, living in symbiosis with the macro-organism. They colonize the host in waves during ontogenesis and through various adaptation mechanisms enter into a permanent relationship with it. While primary colonization can last for months, subsequent waves are shortened and in adults reach 2-4 weeks. Commensals are protected by innate immunity by localizing in the surface layer of the mucous membrane, while the acquired one kills pathogens that have invaded the deep layer through lysis. Pneumococcus, although considered the main cause of pneumonia, is a member of the healthy nasopharynx in eubiosis, as Staphylococcus aureus and other microorganisms. From
the permanent members, those bacteria that have an immunomodulatory effect are called autobionts and those that can cause diseases are called pathobionts. They are also permanent members, but in limited populations. In eubiosis, autobionts and pathobionts are in perfect balance with the host. Autobionts are an evolutionarily developed part of the normal microbiota, actively participating in the immune regulation of the host and the maintenance of health [71]. The regulation of the host immune responses is carried out through their influence on the maturation and functioning of different types of immune cells – IgA secreting plasmotic cells, Th17, Treg lymphocytes, invariant natural killer T cells (INKT), NK cells, macrophages, dendritic cells (DC) and etc [72]. The immune effects controlled by the microbiota also play an autoregulatory role on the microbiota itself. IgA secreting plasmatic cells induced by commensal bacteria are involved in the control of their number and composition, a mechanism also observed in limiting the activity of phages [73, 74]. The gut microbiota controls immune status in an effector or regulatory manner [75]. The barrier function in the nose and nasopharynx is expressed in direct competition and indirect immune modulation between commensal and potentially pathogenic bacterial species. When pathogens attempt to colonize mucosal surfaces, they elicit a strong immune response aimed at clearing them.
Mechanisms used by commensals in eubiosis to suppress pathogen colonization: Induction of IFN-λ secretion from the nasal mucosa.
- secretion of antimicrobial peptides, bacteriocins and proinflammatory cytokines.
- influence on the adaptive immune response and generation of immune memory. Streptococcus mitis induces cross-reactive immunity (antibodies and IL-17) to S. pneumoniae in mice. The same has been observed with Neisseria lactamica and N. meningitidis.
- production of antibiotics. Staphylococcus lugdunensis produces lugdunin, an antibiotic with bactericidal activity against S. aureus and S. pneumoniae. Lactobacillus reuteri produces reuterin, which inhibits the growth and development of a number of bacteria and fungi. Streptococcus salivarius produces salivaricin A and B.
- inhibition of binding to mucous membranes. Streptococcus salivarius limits the binding of S. pneumoniae. The genus Corynebacterium competes with pathogens of the URT.
- possibility of commensals of the genus Streptococcus to destroy the formed biofilms (hypothesis) [76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86].
Intestinal autobionts in healthy hosts produce indole-3-propionic acid (IPA), which regulates positive and negative metabolic homeostasis. IPA is a tryptophan product produced mainly by Clostridium sporogenes. It is associated with the maturation of lung cells, prevents allergic inflammation of the respiratory tract and the development of asthma. Patients with low IPA levels in the blood also have insulin resistance, overweight, a tendency towards low- grade inflammation and symptoms of metabolic syndrome, unlike those with high IPA. It is characterized by a negative relationship with polymorbidity. Patients with operational taxonomic units (OTUs) including Ruminococcus, Alistipes, Blautia, Butyrivibrio and Akkermansia are in the high IPA group, while in the low IPA group we observe an abundance of Escherichia-Shigella, Megasphera and the genus Desulfovibrio [87]. The use of antibiotics in the first year of life may have an undesirable effect leading to a decrease in beneficial bacteria in the intestinal microbiota and causing a cascade of potentially harmful effects. It is associated with a high risk of developing various health problems, including those mentioned above. Fungal dysbiosis and colonization with specific fungi can further exacerbate the manifestation of allergic diseases.
Extremely close relationships between microbiota components may have beneficial effects. C. albicans_biofilms promote recolonization of the gut by Bacteroides after antibiotic therapy, possibly due to the hypoxic environment created, facilitating the growth of anaerobic bacteria. In monocolonization, _C. albicans_or _Saccharomyces cerevisiae (together antagonists) support the establishment of intestinal homeostasis and protect us from virus-induced lung inflammation and intestinal barrier disruption [88, 89]. We need to better understand the interactions between the intestinal commensals responsible for IPA production, the factors activating or repressing the genes for production, and the possibility of transduction of genes responsible for toxin production or antibiotic resistance by phages. Changes in taxonomic composition have been directly linked to various inflammatory diseases such as inflammatory bowel disease (IBD) and asthma. An increased amount of histamine- secreting bacteria in the intestines has also been observed in patients with asthma [90, 91, 92]. In addition to antibiotics and diuretics, opiates, beta blockers and other medications can provoke histamine release in one or another way. Induction of the DNA repair system or SOS response is associated with the regulatory response of bacterial cells against the loss of phageome and gut microbiota diversity, pathogenic bacteria in the gut and the induction of prophages [93, 94] Lactobacillus reuteri is thought to induce the SOS response by activating specific metabolic pathways used in the GIT [61]. Our ability to use it as a probiotic monoproduct during the first 3 years of life after antibiotic therapy will likely allow us to reduce the potential risk of developing health problems in at least some of the patients, restoring the lineage specificity of the microbiota.
Antonie van Leeuwenhoek (1632-1723) was the first that investigate the possibilities of coexistence between bacteria and developed the concept of surface-attached microorganisms in the form of dental plaque. Bacteria exist in the human body as individual planktonic forms and in organized ecosystems (biofilms). Up to 99% of bacteria in the human body exist in the form of biofilms. It is a complex system in which individual planktonic forms fuse and attach to various surfaces through glucoconjugated bonds and form an exopolysaccharide matrix. After maturation, the biofilm releases cells that are directed to colonize another surface [95]. It usually contains several strains of bacteria and fungi. The oxygen tension gradient determines the increased metabolic activity at the surface of the matrix and the reduced/quiescentce in the deep layers. The spatial arrangement of the different strains in the biofilm determines the positive and negative(competition) interactions. Its cells are constantly in contact with each other through chemical signaling or quorum sensing (QS), similar to the regulation of the rate of phage induction. The molecules involved in QS can modulate the spatiality between the interacting microorganisms. Two types of signaling molecules determine the expression of specific genes responsible for the synthesis of biofilm components, bacteriocins, spousal transfer of plasmids and the stress response. These molecules or autoinducers have a function similar to signaling hormones and when accumulated, trigger a cascade of events when a threshold concentration or quorum is reached. Communication between bacterial cells involves the production of self-secreted extracellular signaling molecules that accumulate in the local environment and correlate with cell density. They are: acyl homoserine lactones (AHLs), autoinducer-2, oligopeptides, diffusible signaling factors (DSFs) and autoinducing peptides (AIPs). After reaching a threshold concentration, the molecules signal back to the cell, coordinating the expression of virulence factors, sporulation and biofilm formation. In gram-positive (+) bacteria, an autoinducing peptide (AIP) or peptide pheromone has been found that provides a species-specific communication signal [96, 97, 98]. Biofilms are characterized by their high degree of resistance to antibiotics and host immune mechanisms - low susceptibility to opsonization and phagocytosis [99].
Mycobiota
The mycobiota or fungal component consists of transient and permanent members similar to the bacterial ones. It is less abundant and diverse than the other components. It includes the following genera: Candida, Saccharomyces, Fusarium, Debaromyces, Penicillum, Pichia, Cladosporium, Malassezia, Aspergillus, Cryptococcus and others. It works in synergy with the other components of the microbiota, modeling the immunity and physiology of the host. Transient opportunistic fungal pathogens enter the body from environmental reservoirs and are responsible for diseases due to local or systemic suppression of the immune system. They cause superficial infections of the skin, hair and nails, chronic pulmonary fungal infections and systemic infections with a mortality rate up to 90%. Permanent members are commensals living in symbiosis with the macroorganism. Among them there are also opportunistic fungal pathogens. Colonizing the host during ontogenesis, through various adaptation mechanisms they enter into a permanent relationship with it. Fungi constantly pass through the commensal-pathogen continuum, but in eubiosis they are in balance with the host. From opportunistic fungal infections capable of causing systemic diseases in humans, the most studied are the species Candida albicans, Cryptococcus neoformans and Aspergillus fumigatus. Candida albicans is a commensal in about 50% of healthy adults, colonizing the gastrointestinal, urogenital tract and skin. Fungal pathogens undergo different morphological states using a complex of adaptive mechanisms and virulence factors, exude proteases and toxins that aid adhesion, penetration into host tissues, and evasion of the immune system [100, 101, 102, 103].
Complex interactions between viruses, bacteria and fungi, and possibly archaea, play an important role in human health. Fungi and viruses engage in symbiotic, synergistic, competitive and predatory interactions. Synergistic interactions such as disruption of epithelial integrity, suppression of cellular immunity through defective antigen-specific cytotoxic T cell responses, impaired phagocytic activity, production of cytokines and reactive oxygen species (ROS), and formation of extracellular neutrophil traps (NETs) create conditions for fungal coinfection. Mycoviruses can directly infect fungi, affecting their genotype and virulence. Bacteria influence the growth and virulence of fungi and the latter regulate bacterial pathogenesis. Bacteria modulate virulence characteristics - hyphal formation in Candida albicans, germination in Aspergillus fumigatus and capsule formation in Cryptococcus neoformans, melanization and induction of titan cells. Bacteria help in the formation of biofilms by C. albicans_and A. fumigatus - adhesion, hyphal morphogenesis, maturation and dispersion have been followed in detail in _C. Albicans (Table 1).
• Positive interactions between fungi and viruses:
| Fungi | Viruses |
|---|---|
| Aspergillus fumigatus | Influenza virus(104) |
| Aspergillus fumigatus | SARS-CoV-2(105) |
| Aspergillus fumigatus | PIV(106) |
| Aspergillus fumigatus | CMV(107) |
| Aspergillus fumigatus | HRV |
| Aspergillus fumigatus | AdV(108) |
| Mucorales - CAM(mucormycosis) | SARS-CoV-2(109-113) |
| Pneumocystis pneumonia(PCP) | HIV |
| Pneumocystis pneumonia(PCP) | Influenza virus(114-118) |
Table 1: Positive interactions between fungi and viruses.
**
• Observed negative interactions between fungi and bacteria: Inhibition of fungal growth and proliferation: 1. P. aeruginosa, due to the extracellular signal molecule N-(3-oxododecanoyl)-L-homoserine lactones(AHL) in the local environment, can adhere to the filaments of C. albicans, which leads to the death of the fungal cells [125, 126]. 2. P. aeruginosa can also inhibit the growth of C. albicans_by producing phenazine compounds - pyocyanin and in subconcentration inhibits hyphal morphogenesis [123, 127] _3. Lactobacillus spp. inhibit the proliferation of C. albicans_by secretion of fatty and other weak organic acids _4. Lactobacillus spp. by secretion of cyclic dipeptides inhibit fungal growth [128, 129, 130] 5. Actinomyces israelii, P. aeruginosa, Prevotella nigrescens and Porphymonas gingivalis in the oral cavity inhibit the growth of C. albicans. 6. In the intestine, Escherichia coli kills C. albicans_in a magnesium-CD4-dependent manner, depleting magnesium in _C. albicans.
7. Many bacteria, including opportunistic pathogens such as C. difficile, secrete short-chain fatty acids (SCFA) into the colon. These inhibit the growth, filamentation and biofilm formation of _C. albicans_in vitro. [11] When murine models susceptible to _C. albicans_infection were treated with antibiotics, significantly reduced levels of SCFA were observed in the cecum and high fungal loads were observed in the feces. This demonstrates the importance of SCFA in suppressing _C. albicans_over growth in vivo. Generation of strains of _C. albicans_that are unable to form hyphae but that stimulate anti-inflammatory cytokines, providing cross-protection against other intestinal inhabitants, can also be observed [131].
Inhibition of Fungal Virulence Markers Salmonella enterica serovar Typhimirium is able to regulate the virulence of _C. albicans_in an experimental models [123, 132].
Parasites as an additional member of the microbiota Helminths are parasitic worms that infect a variety of hosts. It is estimated that about 2 billion people worldwide are infected with helminths. It has been shown that the interaction between helminths and the host immune system induces immunomodulatory and immunoregulatory mechanisms that ensure their survival in the host.
Immune interactions between the human immune system and parasites Helminths induce a Th2 intestinal immune response characterized by activation of DCs, type 2 macrophages(M2), Tregs, Bregs, Eo, Ba and MCs. This leads to the release of cytokines (IL-4, IL-5, IL-9, IL-10, IL-13, IL-21, IL-25, IL-33) and transforming growth factor (TGF)-β signaling to CD4+ and CD8+ T cells of the adaptive immune system [133, 134, 135, 136, 137]. The chronic intestinal infection favors the pathogenesis of most viruses. Helminths induce strong Th2 cytokine responses that counteract the effect of IFN-γ on the Th1 response and polarize anti-inflammatory type 1 macrophages (M1) toward regulatory type 2 macrophages (M2). They are an example of the ability of a pathogen to modify the immune response to a related or unrelated pathogen, which results in enhancing or weakening tolerance, protective immunity and causing immunopathology. Heterologous immunity is observed in protozoa, parasites, bacteria and viruses.
There are observations in favor of this thesis in ulcerative colitis. The cause of the disease is unknown, but it is assumed to be related to abnormal immunological reactions of the body to bacteria found in the colon. Interestingly, infection with pig whipworm in some patients led to a decrease in inflammation, changing the local immune response. It is important for us to understand and use these changes in immune responses in the diagnosis and treatment of some viral infections. The
transition from a Th1 intestinal immune response to a Th2 response upon infection with intestinal helminths causes IL- 4-enhanced viral replication and blocks the antiviral effects of IFN-γ by regulating the viral latent-to-lytic switch gene 50 [138]. In infected mouse models, reactivation of latent herpes infection has been observed. Again, in such models, the induced Th2 immune response induces Eo recruitment, provokes Eo IL-5 and IL-33 inflammatory cascade, which enhances vaginal epithelial necrosis induced by HSV-2 infection.
In a study conducted in Peru, an increased level of human papillomavirus (HPV-DNA) infection was observed in women infected with helminths. HPV is considered one of the most common human viruses. Most HPV infections are transient, very often pass without almost any symptoms and do not lead to medical problems (clinically insignificant infections). 70% of present HPV infections disappear within a period of about one year and 90% of them are completely curable within two years. For us otolaryngologists, recurrent respiratory papillomatosis (RRP) represents of interest. The first reports of this were by Boyle, et al. [139], Spoendlin, et al. and Arnold [140]. The lesions received their current name by Morell Mac Kenzie in 1871 [141]. Many lesions are initially asymptomatic and subsequently manifest as a voice change. HPV infection/RRP is a disease characterized by recurrent proliferation of benign squamous cell papillomas in the larynx and other parts of the aerodigestive tract. Much has been written, but little is actually known about this relatively rare disease. Only 5% of patients have involvement of the trachea and proximal bronchi, and less than 1% have involvement of the lung parenchyma. In aggressive forms, the interval for spread from primary involvement of the larynx to spread to the lower respiratory tract is usually 12 years, but there are also patients in whom spread occurs much faster. It represents a serious clinical problem due to its localization, resistance to the therapies carried out, frequent relapses, spread to the lower respiratory tract and esophagus (aggressive forms) and the possibility of malignant transformation into squamous cell carcinoma. Suppression of the Th1 immune response may be the basis for its spread and frequent relapses.
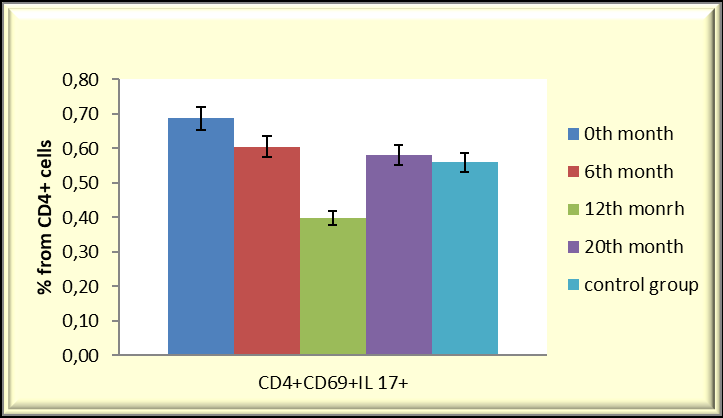
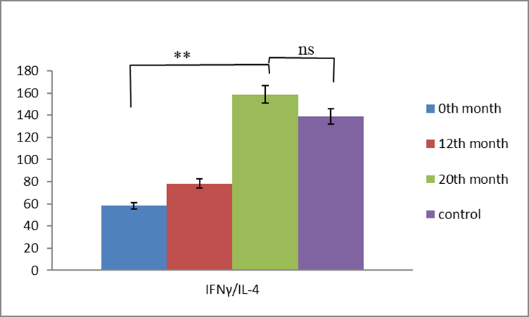
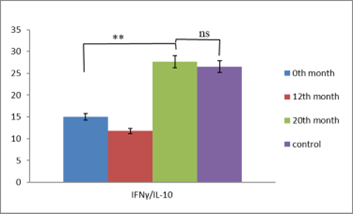
Our observations from immunological studies performed in patients with RRP treated with BCG (Calgevax) according to the scheme showed that it is a chronic viral infection and the effectiveness of the antiviral response depends on the cytokine response. With the administration of BCG, we increase the effectiveness of the antiviral T-cell response by restoring the Th1/Th2/Th17 cytokine balance and inducing Treg differentiation. The latter prevent the depletion of effector clones in the setting of chronic infection and promote the development of a protective T-cell immune response. Our results in the treatment of patients with persistent HPV infection showed that BCG (Calgevax) stimulates the secretion of Th1 cytokines (IFNγ and IL-2) and potentiates the differentiation of Treg at the expense of proinflammatory Th17. This prevents the deepening of inflammation, overstimulation and exhaustion of virus- specific T cell subpopulations [142, 143]. The treatment showed an increase in the duration of remissions in the patients, but not a cure (Figures 1 & 2).
Reduction in Th17 percentage after initiation of therapy
Normalization of IFNγ / IL-4 and IFNγ/IL-10 levels at the 20th month of immunotherapy as a result of increased IFNγ secretion.
In none of the patients treated and monitored by me has a family predisposition been established and I have not tested for helminth carriage.
Mohd Israr et al. in their studies, found Th2/Treg polarization of acquired immunity in papillomas and blood, increased numbers of immature Langerhans cells (iLCs) and increased expression of cyclooxygenase-2/prostaglandin E2 (PGE2) in the upper respiratory tract. When comparing monocytes (Mo) extracted from blood and iLCs from tissues of RRP patients and controls, innate immune dysregulation was observed in RRP patients. In these patients, monocytes generated fewer iLCs than controls due to a reduction in the subpopulation of classical Mo. The authors also observed increased levels of PGE2 in the plasma of patients, responsible for reduced Mo-iLC differentiation. Cytokine/chemokine responses in infected and healthy controls also showed differences. Freshly extracted tissue-derived iLC expressed little CCL-1 and increased CCL-20, which did not respond to IL-36γ stimulation. Therefore, the authors concluded that monocyte/iLC innate immunity is impaired in patients with RRP. They suggested that the immunosuppressive papillomavirus microenvironment is likely responsible for the altered responses of immature Langerhans cells and similar to PIV infection, supports Th2-like/Treg HPV-specific adaptive immunity in RRP [144].
It remains to be clarified whether helminths responsible for chronic intestinal infection, by modifying the immune response to related or unrelated pathogens, favor viral pathogenesis or genetically variations in the innate and acquired immunity of a part of the human population facilitate the modification of the host immune responses to helminths and viral pathogens. Since RRP is rare, it remains to be clarified whether the immunosuppressive papillomavirus microenvironment manifests itself only in a combination of the aforementioned factors.
The upper and lower respiratory tracts possess different ecological niches, which are a combination of taxonomically rich, such as the nose, naso- and oropharynx, and scarce, such as the sinuses and lower respiratory tract, as well as the middle ear. Anaerobic microorganisms are predominant. Loss of oral-nasopharyngeal distinction usually precedes respiratory tract infections [145, 146, 147, 148].
Benign carriage in healthy controls suggests a viral load similar to that of commensal bacteria [149]. Most eukaryotic viruses are RNA, whereas prokaryotic viruses are up to 99% DNA. DNA viruses are thought to have evolved over millions of years, while RNA viruses have adapted to the human population over about 1000 years. From DNA viruses, 87% have adapted, while only a small proportion have done so in RNA viruses. The adaptation has occurred through mutations, rearrangements or gene recombinations, leading to the formation of stable viral lineages in the human population. Some of them circulate asymptomatically until new clinical manifestations are discovered. The origin of the majority of human viruses is unknown, but the majority of them can be categorized as „diseases of the crowd“, requiring relatively high population densities to stabilize. Over time, some viral species tend to disappear, while others evolve in their natural hosts. Most often, new species arise as a result of jumping from one host to another, crossing the species barrier, with humans being simply an “accidental” host. A small proportion of viruses are able to persist in certain human populations (endemics) or spread between different populations (epidemics). The metagenomic analysis, in contrast to PCR-based analysis, showed the presence of many more viral sequences in children with unexplained fever. It is believed that the virome is a common cause of upper respiratory tract diseases.
The respiratory viruses that most commonly attack the human respiratory tract are: ssRNA - influenza virus, parainfluenza virus (PIV), RSV, measles virus, rhinovirus (HRV), coronavirus (CoV) and dsDNA - adenovirus(AdV) [150]. Viral recognition by cells of the innate immunity (detrital cells - DC and macrophages - M) activates a signaling cascade leading to NF-kB-mediated induction of proinflammatory cytokines IL-6, TNF and IL-1, and IFN regulatory factor 3 (IRF3), as well as IRF7 mediating the induction of IFN type I(IFN-α/β) and type III(IFN-λ). IFNs play a fundamental role in effective antiviral immunity [151, 152]. Most respiratory viruses, particularly influenza and SARS-CoV-2, activate the nucleotide-binding oligomerization domain (NOD)-like receptor family purine domain-containing 3(NLRP3) inflammasome via virally encoded hydrophobic proteins that oligomerize in host cells and form hydrophilic pores. The two-step process of tissue injury activates the NLRP3 inflammasome and through activation of caspase-1 and IL-1β, and IL-18 leads to the recruitment of macrophages, monocytes and neutrophils to the site of infection [153, 154, 155, 156, 157, 158, 159].
Рrovoked Immune Interactions
Various respiratory viruses bind to and infect epithelial cells of the respiratory tract expressing specific receptors. This requires knowledge of the cytoskeleton covering the different parts of the respiratory tract. Pseudostratified epithelium, composed of ciliated and secretory cells, covers most of the upper respiratory tract and the trachea. Cuboidal epithelium covers the lower respiratory tract. Squamous alveolar type I cells, which are involved in gas exchange, together with cuboidal alveolar type II cells, form the alveoli [160] Influenza virus entry into cells is due to binding of viral hemagglutinin to sialo-oligosaccharide receptors attached to the surface via α2,3 and α2,6 linkages. IAV (H1N1) binds primarily to α2,6-linked receptors, which are prevalent on non-ciliated cells of the upper respiratory tract, while avian influenza H5N1 and H7N9 bind to α2,3-linked receptors on ciliated epithelial cells of the lower respiratory tract.(161- 165) SARS-CoV and SARS-CoV-2 primarily target type II pneumocytes expressing angiotensin-converting enzyme 2 (ACE2), with infection of alveolar macrophages supporting viral replication. However, SARS-CoV-2 also replicates in the epithelium of the upper respiratory tract, allowing for efficient transmission [166, 167, 168, 169]. Productive viral infection of specific epithelial cells determines the clinical manifestations of the disease. Respiratory viruses cause a variety of changes in the respiratory tract, including alterations in extracellular matrix components that facilitate adhesion and damage to the epithelial cytoskeletal complex that compromises barrier function.
Determining the causes and mechanisms leading to the replacement of a “healthy carrier” by viral pathogens can help us understand the interactions between the virobiota and viral pathogens, the establishment of individual infectious risk and the dynamics of the course of the disease. Viral interactions or interferences depend on:
- the ability of the interfering virus to induce a rapid IFN response expressed in the expression of IFN-stimulating genes(ISGs) type I(IFN-α/β) and type III(IFN-λ), and providing temporary non-specific immunity to the host. The release of effectors that directly inhibit viral replication - chemokines and cytokines, triggering viral defense [170, 171].
- the degree of sensitivity of the second virus to immune mediators
- the extent to which different viruses counteract the induction and antiviral effects of IFN and
- the pattern of virus-induced innate immune responses in the respiratory tract [172].
- Depending on whether infection of the first virus enhances or attenuates infection and replication of the second virus, we observe a positive (synergistic) or negative (antagonistic) interaction. The viruses can infect the respiratory tract simultaneously or sequentially.
The creation of a division of labor (DOL) in positive interactions allows for the reduction of their metabolic burden. It is an association of strains that allows the performance of complex tasks. Energetically expensive pathways requiring cellular building blocks and ATP are shared between strains. Giri in 2019 formulated four criteria for determining whether interactions constitute DOL. They are:
- functional complementarity - each partner in the interaction performs a function better.
- the interaction involves a synergistic advantage.
- the interaction is maintained over a long period of time.
- natural selection favors positive assortment.
Positive interactions between viruses allow a reduction in metabolic burden by creating a division of labor. Positive interactions have been observed with:
- SARS-CoV-2, RSV and pandemic influenza A (pH1N1) [173]
- PIV1 and PIV2
- RSV and HMPV In HBV and HDV, the surface antigen of the former serves as a receptor for the second (HbSg of HBV for HDV). These are not respiratory viruses, but they clearly illustrate how our defenses can be circumvented. Coinfection increases the severity of the disease by excessive production of IFN and proinflammatory cytokines or by reduced secretion of non- inflammatory mediators such as IL10 [174].
In negative interviral interactions, we observe blocking and/or reduction of cell surface receptors and competition for cellular resources. They are homologous and heterologous depending on whether the viruses belong to the same or different families.
In homologous interaction, cross-reactive immunity against the first virus prevents infection by the second virus. A hierarchical pattern has been observed for IAVs (pH1N1, H1N1 and H3N2) and time pattern for RSV, HMPV and PIV. These are an expression of their taxonomic affiliation to the same family.
In heterologous interaction the provoking of a non- specific immune response by the first virus reduces or prevents infection and replication of the second virus:
- coinfection with IAVs (H1N1 or H3N2) in MDCK cells inhibits RSV replication by removing sialic acid from the cell surface and competing for viral protein synthesis [175, 176, 177].
- oral administration of live enterovirus vaccines (LEV) in children reduces the detection of some unrelated respiratory viruses - influenza, PIV, RSV, HRV and AdV [178, 179].
- previous infection with IAVs (H1N1 or H3N2) prevents subsequent infection with retroviruses.
- IBV prevents subsequent infection with RSV [180].
- IBV reduces the rate of AdV infection [181].
- RSV reduces the rate of HRV infection.
- RSV reduces the probability of detection of HMPV.
- HRV limits the replication of SARS-CoV-2.
- HRV reduces the likelihood of IAV detection.
- Influenza and SARS-CoV-2 viruses employ a broader range of mechanisms to evade IFN induction and signaling compared to RSV, HMPV and HRV.
The bacterial pathogens that most commonly attack the human respiratory tract are: Gram(+) - Staphylococcus aureus, Streptococcus pneumoniae, Streptococcus pyogenes and Mycobacterium tuberculosis, Gram(-) -Haemophilus influenzae, Neisseria meningitidis, Bordetella pertussis and in immunocompromised patients Pseudomonas aeruginosa [182].
Viral-bacterial interactions play a critical role in the pathogenesis of bacterial infections. Knowledge of the interactions between viral pathogens with pathobionts and exogenous pathogens will allow us to predict the severity of respiratory diseases. We observe three types of interactions:
- the virus potentiates bacterial colonization
- bacteria enhance viral infection by activating host proteases
- proteases of respiratory tract bacteria cause structural changes leading to increased pathogenicity and tissue tropism of the virus [183, 184, 185, 186, 187].
Together, viruses and bacteria cause diseases more severe than those caused by either pathogen alone, which explains why the majority of deaths during influenza epidemics are due to secondary bacterial infections [188, 189, 190, 191, 192]. They are expressed:
- A prior viral attack, by damaging the mucociliary system, facilitates bacterial colonization of the airway
- surface. In RSV infection, we observe loss of cilia in human bronchial cells in vitro, and in influenza virus, damage to the ciliated epithelium and bronchial epithelial lining [193, 194, 195]. Disruption of epithelial integrity impairs the mechanical clearance of pathogens and facilitates secondary infection. Injured cells or cells in an intermediate state of differentiation express apical α5β1 integrin receptors to which bacterial and fungal pathogens attach [196, 197] Respiratory viruses disrupt cytoskeletal organization, influence innate immune responses (IFN-dependent induction of IFN- stimulated genes) and using the enzyme neuraminidase, they cleave mucins, which form a primary protective barrier that prevents pathogens to reach the underlying epithelium [198, 199] Impairment of immunoregulatory mechanisms prevents epithelial cells to control hyperinflammation by releasing anti-inflammatory mediators. This leads to uncontrolled inflammation, tissue damage, immunopathology and increased susceptibility to bacterial and fungal coinfections. In severe influenza infection, the shedding of fibrinous material and cells in the distal airways causes reduced diffusion of oxygen and carbon dioxide. Hypoxia affects the virulence of the pathogen and the immune responses of the patient. Activated endothelial cells release mediators, pro-inflammatory cytokines, platelet activating factor and adhesion molecules that enhance tissue destruction and inflammation of the small airways [200, 201, 202, 203, 204, 205]. A hypoxic environment is an important factor facilitating coinfection.
- Virus-induced changes in host cell membrane potential lead to increased bacterial adhesion. Viral glycoproteins expressed on host cell membranes serve as bacterial receptors. Alteration of the glucoconjugate structure of murine nasopharyngeal mucosa caused by influenza infection is associated with changes in lectin binding [206, 207]. The influenza virus hemagglutinin esterase (HE) on infected MDCK cells serves as a receptor for group B streptococci. As a possible mechanism for staphylococci to attach to virus-infected cells in vivo, it is assumed that the viral antibody serves as a receptor for staphylococcal protein A. Staphylococci attack only those mucosal surfaces that are damaged by the virus. Immunoglobulin superantigens such as protein A of S. aureus can bind to immunoglobulins secreted by mast cells (MCs). Their activation leads to degranulation and release of histamine and leukotrienes. Examples of activation of MCs by bacterial superantigens include enterotoxins A and B and superantigen-like proteins (exotoxins) from S. aureus [208, 209, 210, 211]. Injury to the respiratory epithelium by other viruses may occur in a similar manner [212, 213]. RSV glycoproteins F and G induce increased attachment of Neisseria meningitidis to cells (214).
- Viral infections suppress host defence mechanisms against bacterial attack: nonspecific humoral factors, nonspecific phagocytosis by neutrophils and macrophages early in the infection, and later specific antibody- mediated immune responses. Suppressed chemokine production leads to reduced neutrophil recruitment and dysfunction due to defective myeloperoxidase (MPO) in the area of inflammation, reactive oxygen species (ROS) and formation of neutrophil extracellular traps (NETs). Influenza virus-induced polymorphonuclear dysfunction underlies secondary pneumococcal disease. Its action on human neutrophils in vitro causes reduced chemotaxis, phagocytic activity and reduced bactericidal potency of neutrophils and macrophages against staphylococci due to impaired lysozyme production by both phagocytes [215, 216, 217, 218]. Interactions between neutrophils and effector cells, monocytes and macrophages, lead to activation of the macrophage inflammasome by respiratory viruses and prevent their depletion during coinfection with S. pneumoniae [219, 220]. Monocytes induce neutrophil activation and ROS production through type I IFN secretion. Neutrophils and Mos also control the maturation and proliferation of DCs. In patients with severe COVID-19, errors in genes involved in regulation cause defective monocyte activation and dysregulated myelopoiesis with release of immature neutrophils into the circulation. Flow cytometry analyses reveal a redistribution of monocyte subsets with a predominance of intermediate monocytes with a hyperinflammatory signature and the appearance of suppressor-like monocyte myeloid cells. Excessive activation and/or recruitment of phagocytes can lead to lung injury [221, 222, 223, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238]. Influenza virus is the most studied example of a positive collaboration between a virus, bacteria and fungus. Influenza primarily causes upper respiratory tract infections, but when the lungs are involved it can be fatal due to pulmonary edema and hemorrhage. RSV results in suppression of TNFα production and bactericidal activity against H. influenzae and S. pneumoniae [239].
- Altered IFN response following viral infection. SARS-CoV and SARS-CoV-2 induce a lower antiviral transcriptional response, expressed as low levels of IFN types I and III and increased expression of chemokines, in contrast to other respiratory viruses including IAV. This is presumably due to antibodies against IFN type I or “errors” in genes involved in the regulation of IFN type I and III immunity. Uncontrolled IFN production causes tissue damage and immunopathology.
- Type I IFN(IFN-α/β) is responsible for lymphopenia in severe influenza and SARS-CoV-2 infections and increases the likelihood of secondary bacterial and fungal infections. The type I IFN response is critical for the development of ARDS and increased mortality in severe SARS-CoV-2 infection [240, 241, 242].
- Type III IFN(IFN-λ) reduces epithelial proliferation and differentiation, and may impair lung recovery after influenza, as well as increase susceptibility to coinfections [243, 244, 245, 246, 247, 248, 249] High levels of type III IFN in the upper respiratory tract and to a lesser extent type I IFN determine mild pathology in patients, whereas high levels in the lower respiratory tract are associated with severe COVID-19 [250, 251].
- Type II IFN in influenza infection contributes to increased susceptibility to secondary bacterial infections by depleting alveolar macrophages and suppressing their capacity. IFN-γ regulates Th17 memory responses and reduces bacterial clearance after influenza infection [252, 253, 254].
- A delayed type I IFN response is associated with increased viral persistence and inflammation, while an early type I IFN response, which limits viral replication, results in mild disease [255, 256].
- Impaired NK cell function during influenza and SARS- CoV-2 infections leads to increased susceptibility to coinfections (257).
- Short- and long-term effects of damage to dendritic cells (DCs) by viral infections. DCs are of two types- myeloid conventional (cDCs) and lymphoid plasmacytoid (pDCs). pDCs secrete type I IFN, causing expansion of antigen- specific T cells and inhibiting viral replication in airway epithelial cells. In severe COVID-19, apoptosis of pDCs is observed, correlating with their reduced number [258, 259, 260]. cDCs are involved in antigen presentation and T cell activation, which is the basis of adaptive immunity. After antigen recognition, uptake and processing, they mature and migrate to regional lymph nodes to present antigen peptides to CD8+ or CD4+ T cells. In severe viral infections, both short-term and long-term effects of DC damage are observed. In the former case, DC signaling pathways for antigen presentation are restored immediately after viral infection is cleared. Human herpesviruses (HHVs-DNA) affect the antigen presentation pathway. RNA viruses - influenza and SARS-CoV-2 use antigen cross-presentation - exogenous antigens loaded in the major histocompatibility complex class II(MHC-II) are transferred via the MHC-I pathway. In influenza infection, dendritic cells (DCs) that capture dead cells containing influenza virus are unable to activate CD8+T cells specific for cell-associated antigens on captured cells and show impaired antigen cross- presentation. SARS-CoV-2 infection induces reduced numbers of DCs. The functional impairment is expressed in impaired maturation, cytokine production and impaired T cell activation [261, 262, 263, 264] Long-term damage can persist for weeks or even months, during which time we observe an increased susceptibility to coinfections. Studies have shown that influenza infections induce metabolic reprogramming of DCs, leading to significant alterations in their innate immune functions and reduced motility and impaired T cell activation. However, the duration of this reprogramming is unknown [265].
Understanding the negative interactions between viruses and bacteria is crucial for our assessment of infection risk. It has been suggested that chronically resident viruses in human tissues such as HHVs, Polioviruses, AdVs, HPVs, HBVs, HCVs and HIV, by causing acute and chronic infections, may prevent colonization of bacterial pathogens in the gut. A mouse model with latent herpes infection has been shown to be resistant to infections caused by Listeria monocytogenes and Yersinia pestis. This is due to a activation of innate antiviral immunity, expressed in cytokine production and macrophage activation [266].
Jennifer Klunk et al.’s study of the impact of pathogens on the type of immune response and variations in immune- related genes provides some answers about how such pandemics contribute to our susceptibility to infection. It examines the impact of Yersinia pestis during the second plague pandemic in shaping the human immune system. Before the pandemic with the highest mortality in history, it is suggested that Europeans were likely an immunologically naive population with little or no adaptation to Yersinia pestis. This study tracks target immune genes of immune- related processes responsible for the expression of innate immune receptors, immune transcription factors, cytokines, chemokines and other effector molecules. To identify alleles conferring protection or increased susceptibility to Yersinia pestis, the authors searched for target regions for variants showing unexpectedly large changes in allele frequency in samples before and after the plague pandemic. They monitored their impact on the gene expression of immune cell types involved in the host response to Yersinia pestis infection. It was found that macrophages are recruited to sites of infection and phagocytize the bacteria. The infection spreads to the lymph nodes, where the bacteria replicate uncontrollably, so some pathogens survive. The authors used incubated macrophages derived from monocytes. They analyzed changes in gene expression of macrophages in their study using data from cross-infection with live Listeria monocytogenes Gram (+) and Salmonella typhimurium Gram (-), as well as monocytes activated by the Toll-like receptors (TLR, TLR1/2, TLR4 and TLR7/8) pathways. In vivo, TLR4 detects Yersinia pestis by recognizing lipopolysaccharide (LPS) on the bacterial membrane. Y. pestis attempts to evade detection by deacylating surface LPS. The authors identified a protective C allele that confers protection against Y. pestis by increasing sensitivity to LPS and promoting an effective immune response in contrast to the putatively deleterious T allele. They experimentally demonstrated that carriers of the locus with the protective C allele present a greater variety of antigens via major histocompatibility complex molecules to CD8+ T cells, stimulating a protective immune response against Y. pestis. In addition to its role in antigen presentation and activation of CD8+ T cells, the locus is involved in viral clearance and cytokine responses. Levels of the granulocyte colony-stimulating factor IL-1β are significantly reduced in the presence of the protective C alleles, while levels of CCL3, involved in neutrophil recruitment upon infection, are increased. Individuals carrying the locus with more copies of the selectively favored protective allele are better able to limit intracellular replication of Yersinia pestis. Macrophages of individuals, possessing a protective allele, engage in a unique cytokine response to the pathogen and are able to limit bacterial replication in vitro. The authors came across an interesting phenomenon. The selectively favorable variant, however, is a risk factor for Crohn’s disease and other candidate loci are associated with increased risk of rheumatoid arthritis and systemic lupus erythematosus. For now, the evidence for balanced selection, i.e. the relationship between autoimmune risk alleles and adaptation to past infectious diseases, remains tenuous, as the agents driving selection remain elusive. We need to investigate whether latent or chronic herpes intestinal infection due to vertical gene transfer provides a protective C allele against Y. pestis by increasing sensitivity to LPS and promoting an effective immune response. We also need to clarify the reason for the difference in HERV expression in patients with inflammatory bowel disease and healthy individuals [267, 268, 269, 270, 271].
When the immune system is weakened or damaged due to a viral infection, almost any infection can become opportunistic. Depending on the causative agent, they can be: viral, bacterial, fungal or parasitic.
- Viral opportunistic infections include: a. Cytomegalovirus (CMV) from the respiratory virus family, b. Human polyomavirus or John Cunningham virus causing multifocal leukoencephalopathy and Human herpesvirus 8 or Kaposi sarcoma.
- Opportunistic bacterial infections include: Clostridium difficile causing gastrointestinal infection, Legionella pneumophila-respiratory, Mycobacterium avium complex-a typical coinfection by two bacteria- Mycobacterium avium and М. intracellulare-respiratory, Mycobacterium tuberculosis-respiratory, Pseudomonas aeruginosa-respiratory, Salmonella-gastrointestinal, Staphylococcus aureus including methicillin-resistant strains, Streptococcus pneumoniae and Streptococcus pyogenes - respirator
- Fungal infections include: Aspergillus-respiratory, Candida albicans-most often oral and gastrointestinal, Coccidioides immitis - coccidioidomycosis or Valley fever, Cryptococcosis neoformans - cryptococcosis causing both respiratory and nervous system infections including meningitis, Histoplasma capsulatum-histoplasmosis - respiratory, Microsporidia - microsporidiosis mainly in immunocompromised patients, Pneumocystis jirovecii or Pneumocystis carinii - causes pneumocystis pneumonia.
- Opportunistic parasitic infections include: Cryptosporidium toxoplasma gondii.
Mast Cell Activation
Mast cells (MC) are responsible for the secretion up to 200 different mediators and the protection of the host from pathogens. They are found in “control” zones interacting with the environment such as the skin, mucous membranes, lungs and intestines. They are components of innate immunity. Located mainly in the subepithelial layer adjacent to blood vessels, they are also in contact with other control cells - dendritic cells. The connection between MCs and blood vessels help to quickly pull effector cells from outside the bloodstream. This facilitates the production of cytokines - TNF and IL-1β, activating the endothelium and lipid mediators facilitating vasodilation and the production of chemokines. MCs are a major local source of IFN types I and III. Their interactions with viruses and their products are complex and can lead to both harmful and positive effects. In infection, MCs can stimulate effective immunity in some cases and at the same time have the potential to cause tissue damage and endothelial barrier dysfunction in secondary infection when their numbers are increased. Their activation causes symptoms in the cardiovascular, digestive, nervous, respiratory systems, skin and mucous membranes as well as hormonal imbalance.
- When MCs are infected with PIV, histamine and leukotrienes are released [272]. In rat models, there is evidence of a higher number of activated MCs in the airways, Th2 dominance and more severe airway inflammation [235, 236].
- MCs degranulation associated with RSV infection in vivo has been observed in bovine models. The degranulation and release of lipid mediator is the cause of bronchospasm in infants with RSV infection. The presence or absence of virus-specific IgE during MC activation is associated with complement activation or not, with degranulation and release of lipid factor or limited degranulation and generation of leukotrienes. RSV demonstrates limited transcription in human MCs. Upon contact with them, the latter induce significant production of chemokines and type I IFN [273, 274, 275].
- In human MC lines, there is limited evidence for productive IAV replication and production of cytokines, chemokines and type I IFN [276].
- HRV infection is best studied as being associated with asthma exacerbations. MC lines release mediators, generate leukotrienes and induce IFN. In asthmatics, insufficient production of IFNβ is observed. HRV provokes apoptosis of human MC lines. In HRV infection, MCs are productively infected, but retain their ability to activate host defense processes. In influenza and RSV, MCs are resistant to productive infection, but trigger a protective response expressed in the production of cytokines, chemokines and the recruitment of antiviral effector cells. Human MCs produce significant amounts of type I IFN, leading to local antiviral response and increased resistance to infection. In cases of severe infection and increased numbers of MCs, these immune responses can lead to potentially damaging inflammation [277, 278, 279].
Seasonal associations are observed in viral and bacterial infections. Influenza, pneumococcal infection and meningococcal disease occur during the winter months [280, 281, 282]. Similar associations with S. pneumoniae have been found for RSV and PIV [283, 284]. There is no evidence of a seasonal association for the seasonal coronaviruses (HCoV-229E, HCoV-NL63, HCoV-OC43 and HCoV-HKU1) and they are thought to act independently [187].
Bacterial proteases in respiratory tract cleave influenza virus HA and increase viral pathogenicity in vivo. This was first demonstrated for the protease from S. aureus and subsequently to Streptomyces griseus and Aerococcus viridans. Cleavage of HA by S. aureus and A. viridans enhances viral replication and pathogenicity in mice in vitro [147, 285, 286, 287, 288] (Table 2).
| Viruses | Bacteria |
|---|---|
| Adenovirus | Moraxella catharralis |
| Adenovirus | Bordetella pertussis |
| Measles virus | Streptococcus pneumoniae |
| RSV | Haemophilus influenzae |
| RSV | Streptococcus pneumoniae |
| RSV | Bordetella pertussis |
| RSV | Staphylococcus aureus |
| PIV | Streptococcus pneumoniae |
| Rhinovirus A | Streptococcus pneumoniae |
| Rhinovirus A | Haemophilus spp |
| Rhinovirus C | Streptococcus pneumoniae |
| Rhinovirus C | Moraxella catharralis |
| Influenza A virus | Streptococcus pneumoniae |
| Influenza A virus | Staphylococcus aureus |
| Influenza A virus | Neisseria meningitidis |
| Influenza A virus | Moraxella spp. |
| Influenza A virus | Corynebacterium spp. |
| Influenza A virus | Haemophilus influenzae |
Table 2: Positive virus-bacterial interactions [284,289-293].
In children, RSV and bacteria interact more frequently than influenza virus and bacteria. Other positive viral- bacterial interactions are:
- Sendai virus enhanced respiratory infections with Mycoplasma pulmonis [294].
- Reovirus - Staphylococcal infections [295].
- CMV - P. aeruginosa infections [296]. Several respiratory viruses are thought to be associated with sudden infant death syndrome (SIDS). These include RSV, influenza virus, PIV, adenovirus and HRV. Changes in the bacterial population of the nasopharynx, particularly increases in Staphylococcus aureus and Enterobacteriaceae, have also been associated with SIDS [297, 298].
Bacterial pathogens compete with each other for space and energy resources, and information about their interactions is also important for us. Competitors of pathogens can sometimes become our allies. Intraspecific competition is observed when two different groups of S. aureus are co-infected. One group of S. aureus, through QS inhibition, suppresses the other by secreting autoinduced peptides (AIPs). In 5 to 40% of the human population, pneumococci are normal inhabitants of the upper respiratory tract. Nasopharyngeal colonization with S. pneumoniae protects against S. aureus and reduced S. pneumoniae counts after pharmacological intervention lead to increased S.
aureus. Reduced S. pneumoniae colonization also leads to increased H. influenzae, N. meningitidis and M. catarrhalis [299, 300, 301, 302]. Therefore, we need to carefully consider our antibiotic interventions. Invasive streptococcal infection (IPI) is caused by 20 to 30 serotypes of S. pneumoniae. To evade the immune response, pneumococci easily change their capsular serotype through genetic transformation. A phenomenon we are facing is that pneumococcal vaccines (PCV-10/13/21 and PPSV-23) due to limited coverage cause replacement of vaccine serotypes of S. pneumoniae with non- vaccine ones in the nose and nasopharynx and increased carriage of non-typable Haemophilus influenzae. Although they reduce disease caused by vaccine serotypes, overall colonization rates have not changed. This is where probiotics can be of benefit:
- Lactobacillus casei surface protein antigen (PspA) induces antibodies against S. pneumoniae.
- Streptococcus mitis generates immune memory, inducing cross-immunity (antibodies and IL-17) against S. pneumoniae in mice [303].
Microbiota imbalance in acute respiratory infections allows pathogens to spread to adjacent poorer ecological niches and cause disease. Cocolonization and polymicrobial interactions between pneumococci and other respiratory colonizers, nontypeable H. influenzae and M. catarrhalis lead to an increased risk of acute otitis media [304, 305, 306, 307].
Viral infections, by suppressing host defense mechanisms, help to replace the normal microbiota. The use of antibiotics, especially in the first year of life, may have an additional unintended effect of reducing the beneficial bacteria of the gut microbiota and a higher risk of developing various health problems The disturbed balance between auto-, pathobionts and external pathogens facilitates the colonization and replacement of histamine-secreting strains in five of the main genera of the human intestinal microbiota, which aggravates the course of the disease. Histamine is a biogenic amine that plays a role in vascular permeability, mucus secretion and neurotransmission. It is endogenous and exogenous. It is produced by decarboxylation of the amino acid histidine by the enzyme histidine decarboxylase. Two families of histidine decarboxylases have been identified: pyridoxal-5- phosphate (PLP)-dependent histidine decarboxylases and pyruvoyl-dependent histidine decarboxylases [308, 309]. Decarboxylation occurs in the bacterial cytoplasm and the histidine/histamine antiporter transports histidine into the cell and then removes histamine from the cell. Histamine- secreting bacteria, generating histamine and other amines, maintain a neutral cytoplasmic pH, which allows them to survive in acidic conditions. They are part of the transient and permanent members of the human microbiota. Exogenous histamine has been studied mainly in food poisoning. Foods with high concentrations of histamine cause neurological, gastrointestinal and respiratory disorders.(310-319) Knowledge of transient bacterial members secreting histamine - Morganella morganii (Proteus morganii) gram(-) bacterium, Hafnia alvei - gram (-) bacterium - commensal and probiotic strain causing diseases in immunocompromised patients, gram(+) members of the genus Clostridium, gram(-) members of the genus Yersiniaceae, mainly Yersinia enterocolitica and Pseudomonas aeruginosa - gram(-) bacterium in immunocompromised patients, is important for us. Antibiotics affect microbial diversity and this is particularly pronounced in anaerobic bacterial species of the intestinal microbiota. Bacteria important for our health that are damaged by their use include: Clostridia, Bacteroides, Lactobacilli and Bifidobacteria.(320) The increased presence of the genus Proteobacteria, species of which contain PLP-dependent histidine decarboxylase, is considered an indicator of intestinal dysbiosis and a cause of inflammatory diseases.(321) In the Genome Taxonomic Database (GTDB), five genera commonly found in the human gut microbiota have putative histamine-secreting bacteria: Bacteroides, Clostridium, Bifidobacterium, Fusobacterium and Lactobacillus [51, 322, 323]. The genomic potential for histamine secretion has been found to be strain- specific within each family. Streptococcus thermophilus, Staphylococcus warneri, Lactobacillus parabuchneri and Lactobacillus reuteri have been studied as specific strains for histamine production [324, 325, 326, 327]. It is due to horizontal gene transfer from mobile gene elements. In turn, temperate phages help the bacterial host to adapt better due to the expression of newly acquired phenotypes [328, 329, 330]. In the above-mentioned five genera, antibiotic therapy is a common reason for replacing their composition with histamine- secreting strains [51, 322, 331, 332]. The presence of them in the families Enterobacteriaceae and Lactobacillaceae in the small intestine can lead to serious immunological consequences for the organism [333, 334, 335, 336, 337]. Adventitious strains of histamine-secreting probiotic bacterial transient members Clostridium, Bifidobacterium and Lactobacillus should also be considered as they may aggravate the course of the disease.
In determining our susceptibility to infection, factors that play an important role are: age, gender, health status, diet, exposure to pathogens, concomitant infection, immune status and the genome of the individuals. Each of these has the ability to alter the abundance and species diversity of the microbiota.
Appropriate antibiotic therapy is an important component of our pharmacological interventions. But it must be based on evidence that it will provide patient benefits, not just putative ones. The observed global increase in antibiotic use between 2000 and 2015 was 65%. Sequencing patients at an early stage of the disease will help us identify increased viral load. Evidence of bacterial carriage by whole genome sequencing (WGS) is much more sensitive in detecting pathogens after antimicrobial treatment, and fluorescence in situ hybridization (FISH) will help us detect bacterial pathogens in biofilms [338, 339, 340, 341]. Amplicon-based sequencing techniques targeting the 16S ribosomal RNA gene (16S rRNA) reduce the time to 2 days and help identify bacterial pathogens in critical infections when standard tests are negative due to prior empirical antibiotic treatment.
Why should antibiotic therapy be well-targeted? Because it is the other most common cause of dysbiosis. The use of antibiotics, even over this relatively short period of time, approximately the length of a human lifespan, has had a profound impact on the evolution of the holobiont. It has led to increased interactions between phages and bacteria by enhancing gene exchange networks, facilitating colonization, bacterial growth and adaptation. The shift in equilibrium also affects other components of the holobiont due to their close relationships. Broad-spectrum antibiotics do not distinguish between commensal and pathogenic bacteria and can lead to a 30% reduction in the microbiota and disruption of bacterial and phage communities. Their use has a long-lasting effect on the intestinal microbiota, especially during the first years of life. Bacterial dysbiosis as a result of antibiotic use leads to an increased severity of influenza infection and a subsequent increased risk of secondary infections. Separately, influenza infection itself can complicate intestinal dysbiosis. Pathogens filling the gap, by secreting toxins, provide themselves with an additional advantage necessary for their survival [320, 342].
In recent decades, we have been faced with an increasing rate of antimicrobial resistance (AMR), which poses a threat to public health. In plasmid-mediated resistance mechanisms, genes can be shared between different microorganisms and spread [343]. The extent of damage to each microbial population depends on the type of antibiotic. They induce the expression of genes responsible for the induction and transcription of lytic genes in prophages and cause lysis of infected bacterial cells. Quinolones are considered to be major inducers of prophages, causing double-stranded DNA breaks. The mechanism of prophage induction is due to DNA damage destabilizing the repressors of prophage induction. The rate of phage induction itself is regulated by quorum sensing (QS). In the induction of prophages by fluoroquinolones and mitomycin C, the bacterial protein RecA serves as a response sensor [344, 345, 346]. When the diversity of the healthy phageome is altered, this provokes the activation of the DNA repair system, regulated by the repressor LexA and the inducer RecA or the SOS response. It is triggered by the loss of phageome diversity, pathogenic bacteria in the gut, and the induction of prophages [347]. During antibiotic treatment, the reduced amount of commensal bacteria is compensated by other pathogens that cause similar or more severe disease, further worsening the clinical picture of patients.
Diet has a significant impact on the microbial composition of the microbiota. It is determined by our habits, customs and individual preferences. Long-term dietary patterns are among the factors determining the enterotype of the gut microbiota [348]. The Bacteroides enterotype is associated with reduced microbial activity and genetic diversity, insulin resistance, risk of obesity and nonalcoholic steatosis, and the Prevotella enterotype with a diet dominated by plant carbohydrates [349]. Changes due to the type of diet play a role in the health-disease balance. The microbiota of infants is much richer even than that of children fed with formula milk with added Dolosigranulum and Corynebacterium. The genus Lactobacillus has probiotic characteristics and is a common inhabitant of the gastrointestinal tract of humans and animals [350]. It belongs to the lactic acid bacteria with a protective function and was described by Döderlein in 1892. It is widely used in the food industry. Their protective function is due to their surface protein layer, providing an antimicrobial inhibitory effect in competition for binding sites on the surface of epithelial cells and in antagonism for entry and replication of viruses, but not for attachment. The genus contains strains producing (HP+) and not producing hydrogen peroxide (HP-). Depending on the transitory balance, lactobacilli producing hydrogen peroxide (HP+) act in a protective, indifferent or abdication state (patients with infection). The ability of lactobacilli to adhere to epithelial surfaces is essential for maintaining persistent colonization in the gut and other tissues of mammals, but a microbiota dominated by the genus Lactobacillus does not always reduce the risk of some infections. A diet rich in dairy products is associated with a richer and more diverse microbiota. A study shows that in dairy farmers, the nasal microbiota is more complex and protects them from infection, as well as limiting colonization by S. aureus [351]. Regular coffee consumption is associated with a fourfold higher amount of the intestinal bacterium Lawsonibacter asaccharolyticus, which necessitates studying the influence of specific foods on the intestinal microbiota. This necessitates ruling out other causes for this, as the increased amount is observed in the European population, while the opposite is true in the Asian population. A study by Qiu, et al. indicated that dietary supplementation with Bacillus subtilis significantly increased the number of Lactobacillus and Bifidobacterium in the ileum and cecum and decreased Coliforms, and Clostridium perfringens in the cecum.
Diet during infectious diseases can have unintended effects and aggravate the course of the disease. It is important to limit the use of histamine-rich foods during the disease such as nuts, tomato paste, mature cheese, cocoa, chocolate, sauerkraut, smoked fish, salami, ham, dried fruits, some fruits and vegetables, yeast and others, as they can intensify neurological, gastrointestinal and respiratory complaints. Almost all organs and systems have the four types of histamine receptors - H1, H2, H3 and H4. The enzymes responsible for the breakdown of histamine are two. DAO (diamine oxidase), which is produced by the intestinal mucosa and breaks down extracellular (free histamine). With reduced secretion or insufficient amount in the intestine, the latter accumulates in the blood. HNMT (histamine N-methyltransferase) is the other and is responsible for the decomposition of intracellular histamine in the bronchial mucosa, CNS, kidneys and liver. Calcium, magnesium and casein in dairy products, as well as iron in dietary supplements, can affect the action of quinolones and tetracyclines.
Therapeutic options Probiotics are increasingly entering our daily lives. Fuller defines them as living organisms contained in food, which when ingested can change the intestinal microbiota and stimulate the immune system. Taken in sufficient quantities, they have certain benefits for human and animal health. In poultry farming, they have already proven themselves as an alternative to antibiotics [352]. Their important properties are: acid resistance, specificity, lack of side effects, reduction of microbial numbers of pathogens and viability during storage. But before using them we must know them well if we want to achieve maximum effect. Microorganisms live together as natural, cultivated and synthetic communities. Their interactions are highly intertwined. The observed interactions between organisms are: positive: a) one- way(commensalism) and b) two-way-cooperation, mutualism and syntrophy, and negative-competition, deception and parasitism. In positive interactions, physical contact between organisms is often necessary – flagellum, nanotubes, membrane and chains of vesicles [353]. Their interactions are based on the sharing of a large set of molecules - enzymes, QS molecules, siderophores and others [354]. Natural self-assembled communities are easier to use in our dietary interventions. Probiotics are mostly assembled cooperative communities - consortia of 2 or more strains. With proper selection and cultivation, this leads to better combined metabolic efficiency (DOL), higher fitness of members and avoidance of competition. Microorganisms grown in the laboratory often have a harder time surviving in real life, due to the specific growing conditions and nutrients that do not always correspond to those in nature. This makes them more vulnerable to the unfavorable environment of the outside world and easy prey for the natural predators of bacteria - bacteriophages. The possible use of synthetic assembled communities could lead to their difficulty in being detected or killed by our immune system and serious health problems. Another problem we may encounter is that, according to the Black Queen hypothesis, during the evolution of natural communities, positive interactions are strengthened by gene loss, leading to dependencies between interacting microorganisms. When interactions between strains become complex, this can lead to individual strains not being able to thrive independently. Transitional strains can introduce evolutionarily lost genes or simply replace commensals, with the result not always being predictable. Transitional strains can introduce evolutionarily lost genes, strains that aid in the synthesis of missing essential amino acids and even strains that may replace commensals, with the result not always being predictable. Knowledge of the ecological niches they inhabit is also extremely important for us clinicians. The biggest challenge before us is to direct the preparations specific to the causative agent and the specific ecological niche so that they reach only the target area for maximum effect. It has been proven that many lactic acid bacteria (LAB) cannot produce the precursors of RNA and DNA or, more precisely, are auxotrophic for nucleosides and nucleic acids [28]. Not all LAB strains can grow or grow poorly in environments poor in vitamins (mainly from the B group - pantothenate, pyridoxine, riboflavin, niacin and biotin), peptides and amino acids (mainly leucine, isoleucine, valine, methionine and glutamate). L. lactis is auxotrophic for 14 of the 20 amino acids, while L. plantarum and L. mesenteroides require 3 to 11 amino acids. Some LAB strains have adapted to hydrolyze proteins in the environment by means of cell wall proteinase (CEP) [355, 356, 357, 358, 359]. They differ in their ability to utilize external resources and oligopeptide intracellular transport system to acquire amino acids. In dysbiosis due to our pharmacological interventions in the nose and throat, bacteria from these niches can migrate to the gut, overcoming physical and chemical barriers and promoting inflammation and intestinal microbial imbalance. We need to reduce the risk of such microbial transfer. Therefore, the correct combination of symbiotic strains is of paramount importance. The use of transient strains leads to increased competition for adhesion and nutrients with those with pathogenic potential (pathobionts and exogenous pathogens) and we will be able to limit the invasion and overgrowth of the latter to some extent [360].
In addition to the “classic probiotics” (Lactobacillus and Bifidobacteria), new bacterial and fungal strains with potential for improving health have been identified. Some of them are: Clostridium butyricum CBM588 or Miyairi strain, Saccharomyces boulardii and Bacillus subtilis.
Proper selection of dietary components, vaccine strains and probiotics will allow us to: • Significantly reduce nasal colonization with potential pathogens such as S. aureus, S. pneumoniae and β-hemolytic streptococci, and hence reduce respiratory infections using Lactobacillus rhamnosus GG, Bifidobacterium Spp, Lactobacillus acidophilus and Streptococcus thermophilus [361, 362, 363].
- Lower incidence of respiratory tract infections in newborns receiving prebiotics and probiotics [364, 365, 366].
- Significantly reduce the abundance of S. aureus and increase the total number of beneficial microorganisms using S. salivarius and S. Oralis [367].
- Anti-inflammatory effects and change in the IL-10/IL- 12 ratio by the use of Bifidobacterium infantis. Use of its immunomodulatory effect expressed in an increase in the number of mucosal dendritic cells and a decrease in Th1 and Th17 helper T cells.
- Induction of antibodies against S. pneumoniae, due to the expression of the surface protein antigen PspA of Lactobacillus casei [368].
- Oral administration of Bifidobacterium longum, observed in mice, induces reactive oxygen species in alveolar macrophages and enhances protection against pulmonary infection caused by Klebsiella pneumoniae [369].
- Probiotics containing strains of the genus Lactobacillus attenuate viral diarrhea caused by rotavirus infection and together with commensals of the genera Lactobacillus and Bacteroides, inhibit the attachment of rotaviruses by modulating glycan receptors on intestinal epithelial cells [370]. _
- _Saccharomyces boulardii CNCM 1-745 secretes a protease that degrades the A169 toxin of C. difficile.
- Separately Everard, et al. found that administration of Saccharomyces boulardi_i changes the gut microbiota, reduces liver fat accumulation, inflammation and total fat mass in mouse models of obesity and T2D. In healthy humans, its administration has no effect on the microbiota, but after diarrhea it helps restore eubiosis of the gut microbiota [371].
- _Clostridium butyricum CBM588 of the genus Furmicutes uses indigestible fiber and releases short-chain fatty acids (SCFA) - acetate and butyrate. They are excreted in the colon. Its effects include modulation of immune homeostasis, improvement of intestinal barrier function, and influence on inflammation [372]. Genomic analyses of some strains of Clostridium butyricum have shown that they lack genes responsible for pathogenicity and encoding toxin production [373]. The observed effects are:
- Butyrate stimulates mucin production by goblet cells in vitro and increases the thickness of the mucus layer [374]. _
- _Clostridium butyricum modulates immune homeostasis by stimulating IL-17 production by γδ T cells, a subset of intraepithelial T cells that act as part of the first line of defense in the lamina propria of the colon [375, 376].
- In mouse models, Clostridium butyricum promotes IL- 10 secretion by Tregs cells through the production of anti-inflammatory lipid metabolites-palmitoleic acid, prostaglandin metabolites and others - in colonic tissues [46]. It also suppresses DC maturation and the appearance of an inflammatory phenotype as well as the levels of proinflammatory cytokines of Th1 and Th17 cells-IFNγ and IL-17 [377].
- Allergic diseases are provoked by inappropriate activation of the Th2 immune response. Th2 cytokines (IL-4 and IL-5) provoke B cells to produce antibodies - immunoglobulin E (IgE) and activate eosinophils (Eo), basophils (Ba) and MCs - immune cells responding to allergy. Although the Th2 response is considered anti- inflammatory, in allergic disease, the overproduction of Th2 cells causes harmful immune responses. Clostridium butyricum can alleviate the allergic reaction, probably by modulating Tregs [378].
- In mouse models of asthma, Clostridium butyricum CGMCC0313.1 reduces airway hyperreactivity and lung inflammation by suppressing the Th2 response and increasing IL-10 production. MCs infiltrate the airways and upon degranulation release inflammatory molecules and proteases, leading to tissue edema in asthmatic lungs. In this model, the number of infiltrating MCs, levels of Th2 cytokines (IL-4 and IL-5), markers of MC degranulation and IgE levels are reduced [379, 380].
- Antibiotic-associated diarrhea(AAD). Seki reported a reduction in the incidence of diarrhea from 59% in the group receiving antibiotics alone to 5% in the group receiving Clostridium butyricum.
- It is suggested that it can modulate the composition of the gut microbiome, increasing bacterial taxa from the genera Lactobacillus and Bifidobacteria.
- Induced phages prevent infection of commensal bacteria by other lysogenic or lytic phages through a phenomenon of superimmunity [381, 382].
- Bacillus subtilis to significantly increase the number of Lactobacillus and Bifidobacteria in the ileum and cecum, and reduce the amount of coliforms and Clostridium perfringens in the cecum.
- The use of probiotics in allergies also brings certain benefits: 1. L. acidophilus significantly improves nasal symptoms in some patients with perennial allergic rhinitis. 2. L. paracasei affects the quality of life in allergic rhinoconjunctivitis in children. 3. L. rhamnosus GG improves the immune response in immunotherapy.
- Much work remains to be done to clarify when and in which patients it is appropriate to use them in conjunction with other therapeutic strategies [383, 384, 385, 386]?
- The use of topical vaccines or oral supplements containing α-hemolytic streptococci - AHS (Streptococcus sanguis, Streptococcus mitis, Streptococcus salivarius and Streptococcus oralis) is also an option to increase microbial diversity.
• Study and use of the metabolic pathways of Lactobacillus microbes. 1. Their use in the decomposition of dietary purines for the production of energy and molecular building blocks by certain strains of the Lactobacillus genus helps break down urates, reduce their reabsorption and hyperuricemia. 2. An important component of the renal microbiota is the uroprotective bacterium Lactobacillus crispatus, which, through the inclusion of CaOx inhibitors and promoters together with a strain of E. coli, controls the crystallization of CaOx and the formation of kidney stones. The use of antibiotics changes the renal microbiota and shifts the balance from beneficial Lactobacillus to E. coli, which stimulates the formation of stones. It is assumed that Lactobacillus is a protective factor and opposite Enterobacteriaceae are pathogenic factor contributing to the formation of urate kidney stones.
Discussion
We are yet to clarify whether it is more correct to consider the phenotype as made up of two components – static (the part of the commensal communities unique to each individual) and dynamic (changing throughout our lives and responsible for newly acquired phenotypes). Based on the “Black Queen” hypothesis for the evolution of natural communities, can it be assumed that regardless of the advantages that food-type dependencies provide, the loss of genes does not make the static component more susceptible to adverse environmental influences such as viral infections, antibiotics and others. Viral infections, by suppressing the host’s defense mechanisms, help replace the normal microbiota and antibiotics affect microbial diversity, especially pronounced in anaerobic species of the intestinal microbiota. Complex interactions between viruses, bacteria, and fungi, and possibly archaea, underlie normal physiological functions or eubiosis. By altering the static component of the phenotype, dysbiosis has profound effects on human health, triggering many diseases.
It was assumed that the cause of paranasal sinus infections is the correspondence between pathogens of the nasopharynx and the osteomeatal complex. However, gene sequencing of the microbiota of patients after FESS revealed a microbiome similar to that of the anterior nose. It remains similar to that of the nasopharynx after surgery for about 6 weeks and then returns to its original state. After antibiotic treatment, the microbiome composition largely returns to its pre-infection state, but the predisposition to re-infection in these individuals is higher. (Ravel J.) Is the restoration of permanent relationships not an expression of the „memory“ of the static component of the niche and is it possible that the intake of prebiotics can accelerate the restoration of the microbiota after antibiotic therapy?
Studies in mouse models have shown that changes in number and diversity in the microbial composition of the gastrointestinal tract during antibiotic therapy can have a negative effect on the patient‘s response to immunotherapy for malignant neoplasms. We cannot exclude a similar effect on the immune response after viral and bacterial infections. Taking an antibiotic shortly before or after starting therapy with immune checkpoint inhibitors (ICIs) significantly affects overall survival (OS) and progression-free survival (PFS). It is believed that taking prebiotics is more appropriate. During digestion, prebiotics are broken down to short- chain fatty acids by bacterial enzymes and support the growth of a certain microbiota. Should we prophylactically administer prebiotics during immunotherapy? Is the use of Saccharomyces boulardii appropriate to restore the intestinal microbiota, as after diarrhea in such patients?
Phenotype formation during the first three years of life, especially the first, is in its infancy because host colonization has not been completed and permanent relationships in the holobiont have not been established. During this period, our interventions must be extremely balanced so as not to push phenotype formation in one direction or another, the consequences of which are currently unpredictable.
Transient members are part of most of the probiotics sold. It is time to define the value of “sufficient amount” for probiotics. We are yet to clarify how long they should be administered and the duration of their life in the gastrointestinal tract? Despite being transient strains, do they have the opportunity to become permanent members of the microbiota? And finally. If they become permanent members, what will happen to the composition of the given ecological niche, especially if they become dominant species? It should not be forgotten that microbes constantly pass through the commensal-pathogen continuum. This will help to avoid problems that may arise in the foreseeable future, similar to those with antibiotics. It will be much easier for us to foresee possible risks than to look for a solution to problems we have caused. The selection and application of probiotics must be differentiated, as some viruses(noroviruses) can suppress many of the beneficial functions of symbiotic bacteria, which has been demonstrated in animal models [387].
The assumption that thanks to our interventions we can observe the possible extinction of a viral strain sounds too good, especially for members of the young and much more aggressive RNA branch of the virus kingdom. It is much more likely, due to their taxonomic affiliation to families, that we observe one or another variant of a hierarchical or time pattern of homologous interaction preventing its manifestation. Circulating asymptomatically after a certain period of time, a „stimulus“ can provoke the initiation of its replication.
We must not forget, that аny microorganism to survive in a given ecosystem passing permanently through the continuum commensal-pathogen depending on the conditions and co- infections. S. sanguis is the first colonizer and commensal in the oral cavity, a vaccine and probiotic strain, but in some patients it can cause severe endocarditis and even death. Conversely, Pneumococcus, although considered the main cause of pneumonia, in eubiosis is a member of the healthy nasopharynx, as well as Staphylococcus aureus and other microorganisms.
The concept of the holobiont will help us understand how this complex, relatively static and at the same time dynamically evolving ecosystem adapts to continuous environmental challenges. Mapping the ecological niches in our organism with the species that colonize them will allow us to predict possible unintended effects, such as a decrease in beneficial bacteria under adverse conditions. The use of probiotics, topical vaccines and dietary components to achieve artificially modeled co-colonies during the first 48 hours of illness may help us to reduce and partially compensate for the impaired functions of the natural community. We also have to answer whether pandemics are not simply a quantitative expression of the qualitative critical changes accumulated in the hologenome due to seasonal viral infections. We also come to the reasonable question of whether the dynamic equilibrium or eubiosis of the holobiont returns to the same equilibrium point as it was before the disease, or does a new equilibrium occur between the components of the holobiont, which in certain situations is responsible for unlocking various diseases, including tumors that were not manifested until then If so, then probably the phenotype is only relatively constant and the aforementioned accumulations lead to continuous partial changes allowing us to adapt to emerging challenges and how not to unconsciously change the equilibrium point with our interventions.
Conclusion
Understanding the interactions between viral and bacterial pathogens and the components of the holobiont is important for us. Individual phenotypes are an expression of the interactions between the host and the associated microbial genomes. Some of the components are unique to each of us and we should strive to avoid abrupt changes in the phenotype. In the personalized treatment of patients with infectious diseases, respiratory PCR panels and platforms for proving viral and bacterial carriage are important but not always available.
We should not be too “strict” on commensal and transient microorganisms that make up the microbiota because of the diagnostic and therapeutic challenges they create for us. They have many advantages. First, billions of years before the emergence of humanity, they have developed mechanisms for survival under adverse conditions. Second, they do not read our studies to make us comfortable in our work. Third, not only the depth, but also the generalization of knowledge is important in understanding the relationships in the holobiont. The ability of four kingdoms that make up the microbiota to live in equilibrium to create conditions for the survival of species and last but not least, unintentional and unmeasured interventions can probably cause changes in the hologenome that trigger new diseases.
Despite the rapid progress of medicine, we still have a lot of work to do. Our search should be directed not only towards the development of new antibiotics, but also towards means that help restore the microbiota. Knowing the relationships in the holobiont will help us avoid superficial handling of them and using pre- and probiotic strains before the main drug treatment. But even with their seemingly correct use, there is a risk of unintentionally triggering a cascade of negative interactions during our interventions, leading to serious risks to patients' health. When we treat one of the components of the microbiota, we indirectly affect the others, which must be taken into account in our therapeutic plan. For now, our task is to find the right time, type and volume of pharmacological, non-pharmacological and dietary interventions. In the future, it is likely that the knowing and use of the information about 10130 protein molecules, 10700 circuits and 101000 metabolic pathways that have been evolutionarily established in nature will allow us to control the course of diseases by using metabolic remodeling of the interactions of the components of the holobiont and pathogens.
References
-
Parvez MK, Parveen S (2017) Evolution and emergence of patogenic viruses: Past, present, and future. Intervirology 60: 1-7.
-
(2022) Weekly epidemiological update on COVID-19-31 August 2022. WHO 107.
-
Mahy BWJ, Collier L (1998) Topley and Wilson’s Microbiology and Microbial Infection. In: Virology, 9th (Edn.), J Clin Pathol 52(3): 237-238.
-
Wever PC, Bergen LV (2014) Death from 1918 pandemic influenza during the First World War: a perspective from personal and anecdotal evidence. Influenza Other Respir Viruses 8: 538-546.
-
Dawood FS, Iuliano AD, Reed C, Reed C, Meltzer MI, Shay DK, et al. (2012) Estimated global mortality associated with the first 12 months of 2009 pandemic influenza A H1N1 virus circulation: a modelling study. Lancet Infect Dis 12: 687-695.
-
(2015) Summary of probable SARS cases with onset of illness from 1 November 2002 to 31July 2003. WHO.
-
Lu S, Xi X, Zheng Y, Cao Y, Liuet X, et al. (2013) Analysis of the clinical characteristics and treatment of two patients with avian influenza virus(H7N9). Biosci Trends 7: 109- 112.
-
Bos KL, Schuenemann VJ, Golding GB, Burbano HA, Waglechner N, Coombes BK, et al. (2011) A draft genome of Yersinia pestis from victims of the Black Death. Nature 478: 506-510.
-
Benedictow OJ (2004) The Black Death, 1346-1353: The Complete History (Boydell Press,2004).
-
Nadhanan RR, Thomas CJ (2014) Colpoda secrete viable Listeria monocytogenes within faecal pellets. Environ Microbiol 16: 396-404.
-
Gourabathini P, Brandl MT, Redding KS, Gunderson JH, Berk SG (2008) Interactions between food-borne pathogens and protozoa isolated from lettuce and spinach. Appl Environ Microbiol 74: 2518-2525.
-
Berk SG, Faulkner G, Garduno E, Joy MC, Ortiz- Jimenez MA, et al. (2008) Packaging of live Legionella pneumophila into pellets expelled by Tetrahymena spp. does not require bacterial replication and depends on a dot/I cm-mediated survival mechanism. Appl Environ Microbiol 74: 2187-2199.
-
Macher J (1999) Bioaerosols: Assessment and Control. Cincinnati, OH: American Conference of Governmental Industrial Hygienists.
-
Lamoth F, Greub G (2009) Amoebal pathogens as emerging causal agents of pneumonia. FEMS Microbiol Rev 34: 260-280.
-
Simon JC, Marchesi JR, Mougel C, Selosse MA (2019) Host-microbiota interactions: from holobiont theory to analysis. Microbiome 7: 5.
-
Bordenstein SR, Theis KR (2015) Host Biology in Light of the Microbiome: Ten Principles of Holobionts and Hologenomes. Plos Biology 13(8): e1002226.
-
Faure D, Simon JC, Heulin T (2018) Holobiont: a conceptual framework to explore the eco-evolutionary and functional implications of host-microbiota interactions in all ecosystems. New Phytologist 218: 1321-1324.
-
Margulis L (1991) Symbiosis as a source of evolutionary innovation: speciation and morphogenesis. In: Cambridge MA MLFR (Edn.), Symbiogenesis and Symbionticism, MIT Press, pp: 1-14.
-
Rosenberg E, Koren O, Reshef L, Efrony R, Rosenberg IZ (2007) The role of microorganisms in coral health, disease and evolution. Nature Reviews Microbiology 5: 355.
-
Zilber-Rosenberg I, Rosenberg E (2008) Role of microorganisms in the evolution of animals and plants: the hologenome theory of evolution. FEMS Microbiol Rev 32(5): 723-735.
-
Huttenhower C, Gevers D, Knight R, Abubucker S, Badger JH, et al. (2012) Structure, function and diversity of the healthy human microbiome. Nature 486(7402): 207- 214.
-
Lloyd-Price J, Abu-Ali G, Huttenhower C (2016) The healthy human microbiome. Genome Med 8(1): 51.
-
European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2015. EFSA J 14: e04634.
-
Zhang, HL, Mnzava KW, Mitchell ST, Melubo ML, Kibona TJ, et al. (2016) Mixed Methods Survey of Zoonotic Disease Awareness and Practice among Animal and Human Healthcare Providers in Moshi, Tanzania. PLoS Negl Trop Dis 10: e0004476.
-
Morris JJ, Lenski RE, Zinser ER (2012) The Black Queen Hypothesis: evolution of dependencies through adaptive gene loss. mBio 3(2): e00036.
-
Sachs JL, Hollowell AC (2012) The origins of cooperative bacterial communities. mBio 3: e00099-12.
-
Zelezniak A, Andrejev S, Ponomarova O, Mende DR, Bork P, et al. (2015) Metabolic dependencies drive species co- occurrence in diverse microbial communities. Proc Natl Acad Sci, USA, 112: 6449-6454.
-
Canon F, Nidelet T, Guédon E, Thierry A, Gagnaire V (2020) Understanding the Mechanisms of Positive Microbial Interactions That Benefit Lactic Acid Bacteria Co-cultures. Front Microbiol 4(11): 2088.
-
Manohar P, Tamhankar, AJ, Leptihn S, Ramesh N (2019) Pharmacological and immunological aspects of phage therapy. Infect Microbes Dis 1: 2.
-
Hannigan GD, Duhaime MB, Ruffin MT, Koumpouras CC, Schloss PD (2018) Diagnostic potential and interactive dynamics of the colorectal Cancer Virome. MBio 9: e02248-e02218.
-
Ungaro F, Massimino L, D’Alessio S, Danese S (2019) The gut virome in inflammatory bowel disease pathogenesis: from metagenomics to novel therapeutic approaches. United European Gastroenterology Journal 7(8): 999- 1007.
-
Wang X, Kim Y, Ma Q (2010) Cryptic prophages help bacteria cope with adverse environments. Nature Communications 1(1): 147.
-
Modi SR, Lee HH, Spina CS, Collins JJ (2013) Antibiotic treatment expands the resistance reservoir and ecological network of the phage metagenome. Nature 499(7457): 219-222.
-
Mowat AM, Agace WW (2014) Regional specialization within the intestinal immune system. Nature Reviews Immunology 14(10): 667-685.
-
Gu BH, Kim M, Yun CH (2021) Regulation of gastrointestinal immunity by metabolites. Nutrients 13(1): 167.
-
Allaire JM, Crowley SM, Law HT, Chang SY, Ko HJ, et al. (2018) The intestinal epithelium: central coordinator of mucosal immunity. Trends in Immunology 39(9): 677- 696.
-
Lebedev K, Ponyakina I (2006) Immunophysiology of epithelial cells and pattern-recognition receptors. Human Physiology 32: 224-234.
-
Keir ME, Yi T, Lu TT, Ghilardi N (2020) The role of IL-22 in intestinal health and disease. Journal of Experimental Medicine 217(3): e20192195.
-
Ohno H (2016) Intestinal M cells. The Journal of Biochemistry 159(2): 151-160.
-
Medina-Contreras O, Geem D, Laur O (2011) CX3CR1 regulates intestinal macrophage homeostasis, bacterial translocation, and colitogenic Th17 responses in mice. The Journal of Clinical Investigation 121(12): 4787- 4795.
-
Łusiak-Szelachowska M, Weber-Dąbrowska B, Jończyk- Matysiak E, Wojciechowska R, Górski A (2017) Bacteriophages in the gastrointestinal tract and their implications. Gut Pathogens 9(1): 44.
-
Champagne-Jorgensen K, Luong T, Darby T, Roach DR (2023) Immunogenicity of bacteriophages. Trends Microbiol 31(10): 1058-1071.
-
Popescu M, Van Belleghem JD, Khosravi A, Bollyky PL (2021) Bacteriophages and the immune system. Annu. Rev. Virol 8(1): 415-435.
-
Wahida A, Tang F, Barr JJ (2021) Rethinking phage- bacteria-eukaryotic relationships and their influence on human health. Cell Host Microbe 29(5): 681-688.
-
Gliźniewicz M, Miłek D, Olszewska P, Czajkowski A, Serwin N, et al. (2024) Advances in bacteriophage- mediated strategies for combating polymicrobial biofilms. Front. Microbiol 14: 1320345.
-
Majewska J, Kaźmierczak Z, Lahutta K, Lecion D, Szymczak A, et al. (2019) Induction of phage-specific antibodies by two therapeutic staphylococcal bacteriophages administered per os. Front. Immunol 10: 2607.
-
Hurst TP, Magiorkinis G (2015) Activation of the innate immune response by endogenous retroviruses. Journal of General Virology 96(6): 1207-1218.
-
Kassiotis G, Stoye JP (2017) Making a virtue of necessity: the pleiotropic role of human endogenous retroviruses in cancer. Philosophical Transactions of the Royal Society B: Biological Sciences 372(1732): 20160277.
-
Chuong EB, Elde NC, Feschotte C (2016) Regulatory evolution of innate immunity through co-option of endogenous retroviruses. Science 351(6277): 1083- 1087.
-
Santora T Ancient Viruses in Our DNA Hold Clues to Cancer Treatment, national cancer institute.
-
Foster SC, Kumar N, Anonye BO, Almeida A, Viciani E, et al. (2019) A human gut bacterial genome and culture collection for improved metagenomic analyses. Nat. Biotechnol 37(2): 186-192.
-
Leigh BA (2019) Cooperation among conflict: prophages protect bacteria from phagocytosis. Cell Host & Microbe 26(4): 450-452.
-
Focà A, Liberto MC, Quirino A, Marascio N, Zicca E, et al. (2015) Gut inflammation and immunity: what is the role of the human gut virome? Mediators of Inflammation 2015(1): 326032.
-
Cao, Z, Sugimura N, Burgermeister E, Ebert MP, Zuo T, et al. (2022) The gut virome: a new microbiome component in health and disease. Ebio Medicine 81: 104113.
-
Tiamani K, Luo S, Schulz S, Xue J, Costa R, et al. (2022) The role of virome in the gastrointestinal tract and beyond. FEMS Microbiol Rev 46(6): 1-12.
-
Dahlman S, Avellaneda-Franco L, Barr J (2021) Phages to shape the gut microbiota? Curr Opin Biotechnol 68: 89-95.
-
Xu HM, Xu WM, Zhang L (2022) Current status of phage therapy against infectious diseases and potential application beyond infectious diseases. Int J Clin Pract 2022(1): 4913146.
-
Brüssow H, Canchaya C, Hardt WD (2004) Phages and the evolution of bacterial pathogens: from genomic rearrangements to lysogenic conversion. Microbiol Mol Biol Rev 68(3): 560-602.
-
Johnson G, Banerjee S, Putonti C (2022) Diversity of _Pseudomonas aeruginosa_ temperate phages. mSphere 7(1): e0101521.
-
Fortier LC, Sekulovic O (2013) Importance of prophages to evolution and virulence of bacterial pathogens. Virulence 4(5): 354-365.
-
Kirsch JM, Brzozowski RS, Faith D, Round JL, Secor PR, Duerkop BA (2021) Bacteriophage-Bacteria interactions in the gut: from invertebrates to mammals. Ann Rev Virol 8: 95-113.
-
Sausset R, Petit MA, Gaboriau Routhiau V, De Paepe M (2020) New insights into intestinal phages. Mucosal Immunol 13: 205-215.
-
Sinha A, Maurice C F (2019) Bacteriophages: uncharacterized and dynamic regulators of the immune system. Mediat Inflamm: 3730519.
-
Clark SE (2020) Commensal bacteria in the upper respiratory tract regulate susceptibility to infection, Curr Opin Immunol. Oct 66: 42-49.
-
Huxley EJ, Viroslav J, Gray WR, Pierce AK (1978) Pharyngeal aspiration in normal adults and patients with depressed consciousness. Am J Med 64: 564-568.
-
Bassis CM, Erb Downward JR, Dickson RP, Freeman CM, Schmidt TM, et al. (2015) Analysis of the upper respiratory tract microbiotas as the source of the lung and gastric microbiotas in healthy individuals. mBio 6 e00037-15-10.
-
Segal LN, Alekseyenko AV, Clemente JC, Kulkarni R, Wu B, et al. (2013) Enrichment of lung microbiome with supraglottic taxa is associated with increased pulmonary inflammation. Microbiome 1: 337-12.
-
Bosch AATM, de Steenhuijsen Piters WAA, van Houten MA, Chu MLJN, Biesbroek G, et al. (2017) Maturation of the infant respiratory microbiota, environmental drivers, and health consequences. A prospective cohort study. Ame J Respir Crit Care Med 196: 1582-1590.
-
Teo SM, Mok D, Pham K, Kusel M, Serralha M, et al. (2015) The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe 17: 704-715.
-
Ivanov II, Honda K (2012) Intestinal commensal microbes as immune modulators. Cell Host & Microbe 12: 496-508.
-
Clemente JC, Ursell LK, Parfrey LW, Knight R (2012) The impact of the gut microbiota on human health: Аn integrative view. Cell, 148: 1258-1270.
-
Honda K, Littman DR (2012) The microbiome in infectious disease and inflammation. Annu Rev Immunol 30: 759-795.
-
Wei M, Shinkura R, Doi Y, Maruya M, Fagarasan S, et al. (2011) Mice carrying a knock-in mutation of Aicda resulting in a defect in somatic hypermutation have impaired gut homeostasis and compromised mucosal defense. Nat Immunol 12: 264-270.
-
Kawamoto S, Tran TH, Maruya M, Suzuki K, Doi Y, et al. (2012) The inhibitory receptor PD-1 regulates IgA selection and bacterial composition in the gut. Science 336: 485-489.
-
Olszak T, An D, Zeissig S, Vera MP, Richter J, et al. (2012) Microbial exposure during early life has persistent effects on natural killer T cell function. Science 336: 489- 493.
-
Kim HJ, Jo A, Jeon YJ, An S, Lee K-M, et al. (2019) Nasal commensal Staphylococcus epidermidis enhances interferon-λ- dependent immunity against influenza virus. Microbiome, 7 :80.
-
Bradley KC, Finsterbusch K, Schnepf D, Crotta S, Llorian M, et al. (2019) Microbiota-driven tonic interferon signals in lung stromal cells protect from influenza virus infection. Cell Rep 28: 245-256.
-
Liu Q, Meng H, Lv H, Liu Y, Liu J, et al. (2020) Staphylococcus epidermidis contributes to healthy maturation of the nasal microbiome by stimulating antimicrobial peptide production. Cell Host Microbe 27: 68-78.
-
Janek D, Zipperer A, Kulik A, Krismer B, Peschel A (2016) High frequency and diversity of antimicrobial activities produced by nasal Staphylococcus strains against bacterial competitors. PLoS Pathog 12: e1005812-e1005820.
-
Latham RD, Gell DA, Fairbairn RL, Lyons AB, Shukla SD, et al. (2017) An isolate of Haemophilus haemolyticus produces a bacteriocin-like substance that inhibits the growth of nontypeable _Haemophilus influenzae_. Int J Antimicrob Agents 49: 503-506.
-
Engen SA, Valen Rukke H, Becattini S, Jarrossay D, Blix IJ, et al. (2014) The oral commensal Streptococcus mitis shows a mixed memory Th cell signature that is similar to and cross-reactive with _Streptococcus pneumoniae_. PLoS One 9: e104306-e104309.
-
Zipperer A, Konnerth MC, Laux C, Berscheid A, Janek D, et al. (2016) Human commensals producing a novel antibiotic impair pathogen colonization. Nature 535: 511-516.
-
Manning J, Dunne EM, Wescombe PA, Hale JDF, Mulholland EK, et al. (2016) Investigation of Streptococcus salivarius- mediated inhibition of pneumococcal adherence to pharyngeal epithelial cells. BMC Microbiol 16: 225.
-
Bomar L, Brugger SD, Yost BH, Davies SS, Lemon KP (2016) Corynebacterium accolens releases antipneumococcal free fatty acids from human nostril and skin surface triacylglycerols. mBio 7: 144-13.
-
Ramsey MM, Freire MO, Gabrilska RA, Rumbaugh KP, Lemon KP (2016) _Staphylococcus aureus_ shifts toward commensalism in response to Corynebacterium species. Front Microbiol 7: 279-15.
-
Santagati M, Scillato M, Patanè F, Aiello C, Stefani S (2012) Bacteriocin-producing oral streptococci and inhibition of respiratory pathogens. FEMS Immunol Med Microbiol 65: 23-31.
-
Ballanti M, Antonetti L, Mavilio M, Casagrande V, Moscatelli A, et al. (2024) Decreased circulating IPA levels identify subjects with metabolic comorbidities: A multi-omics study. Pharmacol Res 204: 107207.
-
Mason K L, Erb Downward J R, Mason K D, Falkowski N R, Eaton K A, et al. (2012) Candida albicans and bacterial microbiota interactions in the cecum during recolonization following broad-spectrum antibiotic therapy. Infection and Immunit 80(10): 3371-3380.
-
Hogan DA, Vik A, Kolter R A (2004) _Pseudomonas_ _aeruginosa_ quorum-sensing molecule influences Candida albicans morphology. Mol Microbiol 54: 1212- 1223.
-
Kostic AD, Xavier RJ, Gevers D (2014) The microbiome in inflammatory bowel disease: current status and the future ahead. Gastroenterology 146(6): 1489-1499.
-
Huang YJ, Boushey HA (2015) The microbiome in asthma. J Allergy Clin Immunol 135(1): 25-30.
-
Barcik W, Pugin B, Westermann P, Perez NR, Ferstl R, et al. (2016) Histamine-secreting microbes are increased in the gut of adult asthma patients. J Allergy Clin Immunol 138(5): 1491-1494.
-
Early antibiotic exposure can trigger long term susceptibility to asthma, Monash University.
-
Sanders D J, Inniss S, Sebepos Rogers G , Rahman F Z, Smith A M (2021) The role of the microbiome in gastrointestinal inflammation. Biosci Rep 41: 3850.
-
Costerton JW, Stewart PS, Greenberg EP (1999) Bacterial biofilms: a common cause of persistent infections. Science 284(5418): 1318-1326.
-
Nadell C D, Drescher K, Foster K R (2016) Spatial structure, cooperation and competition in biofilms. Nat Rev Microbiol 14: 589-600.
-
Di Cagno R, De Angelis M, Limitone A, Minervini F, Simonetti M C, et al. (2007) Cell-cell communication in sourdough lactic acid bacteria: a proteomic study in Lactobacillus sanfranciscensis CB1. Proteomics 7: 2430- 2446.
-
Fontaine L, Boutry C, Guedon E, Guillot A, Ibrahim M, et al. (2007) Quorum-Sensing regulation of the production of Blp bacteriocins in Streptococcus thermophilus. J Bacteriol 189: 7195-7205.
-
Wood AJ, Fraser JD, Swift S, Patterson Emanuelson EA, Amirapu S, et al. (2012) Intramucosal bacterial microcolonies exist in chronic rhinosinusitis without inducing a local immune response. Am J Rhinol Allergy 26: 265-70.
-
Pendharkar S, Brandsborg E, Hammarstrom L, Marcotte H, Larsson (2015) P-G Vaginal colonisation by probiotic lactobacilli and clinical outcome in women conventionally treated for bacterial vaginosis and yeast infection. BMC Infect Dis 15: 255.
-
Allonsius CN, Vandenheuvel D, EFM Oerlemans, Mariya I Petrova, GGG Donders et al. (2019) Inhibition of Candida albicans morphogenesis by chitinase from Lactobacillus rhamnosus GG. Sci Rep 9: 2900.
-
Wang S, Qiangyi Wang, Ence Yang, Ling Yan, Tong Li, et al. (2017) Antimicrobial compounds produced by vaginal _Lactobacillus crispatus_ are able to strongly inhibit Candida albicans growth, hyphal formation and regulate virulence-related gene expressions. Front Microbiol 8: 564.
-
Kaewsrichan J, Peeyananjarassri K, Kongprasertkit J (2006) Selection and identification of anaerobic lactobacilli producing inhibitory compounds against vaginal pathogens. FEMS Immunol Med Microbiol 48: 75-83.
-
Jang SJ, Lee K, Kwon B, You HJ, Ko G (2019) Vaginal lactobacilli inhibit growth and hyphae formation of Candida albicans. Sci Rep 9: 8121.
-
MacAlpine J, Martin DI, Zhongle Liu, Junko Yano, Nicole MR, et al. (2021) A small molecule produced by Lactobacillus species blocks Candida albicans filamentation by inhibiting a DYRK1-family kinase. Nat Commun 12: 1-16.
-
Liang W, Guan G, Dai Yu, ChengJun C, Tao Li, et al. (2016) Lactic acid bacteria differentially regulate filamentation in two heritable cell types of the human fungal pathogen Candida albicans. Mol Microbiol 102(3): 506-519.
-
Yu YX, Fei FU, Kong WN, Xuan QK, Wen DH, et al. (2018) Streptococcus agalactiae inhibits Candida albicans hyphal development and diminishes host vaginal mucosal TH17 response. Front Microbiol 9: 198.
-
Bandara HMHN, Cheung BPK, Watt RM, Jin LJ, Samaranayake LP (2013) Secretory products of Escherichia coli biofilm modulate Candida biofilm formation and hyphal development. J. Investig Clin Dent 4: 186-199.
-
Kim Y, Mylonakis E (2011) Killing of Candida albicans filaments by Salmonella enterica serovar Typhimurium is mediated by sopB effectors, parts of a type III secretion system. Eukaryot Cell 10: 782-790.
-
Cruz MR, Graham CE, Gagliano BC, Lorenz MC, Garsin DA (2013) Enterococcus faecalis inhibits hyphal morphogenesis and virulence of Candida albicans. Infect Immun 81: 189-200.
-
Graham CE, Cruz MR, Garsin DA, Lorenz MC (2017) Enterococcus faecalis bacteriocin EntV inhibits hyphal morphogenesis, biofilm formation, and virulence of Candida albicans. Proc Natl Acad Sci USA 114: 4507- 4512.
-
Brown AO, Graham CE, Cruz MR, Singh KV, Murray BE, et al. (2019) Antifungal activity of the Enterococcus faecalis peptide EntV requires protease cleavage and disulfide bond formation. mBio 10: 01334-19.
-
van Leeuwen PT, van der Peet JM, Bikker FJ, Hoogenkamp MA, Oliveira Paiva AM, et al. (2016) Interspecies interactions between Clostridium difficile and Candida albicans. mSphere 1(6): 00187-16.
-
Boon C, Deng Y, Wang LH, Yawen He, Xu JL, et al. (2008) A novel DSF-like signal from Burkholderia cenocepacia interferes with Candida albicans morphological transition. ISME J 2: 27-36.
-
Guinan J, Wang S, Hazbun TR, Yadav H, Thangamani S (2019) Antibiotic-induced decreases in the levels of microbial-derived short-chain fatty acids correlate with increased gastrointestinal colonization of Candida albicans. Sci Rep 9(1): 8872.
-
Ost KS, O’Meara TR, Zac Stephens W, Tyson Chiaro, Haoyang Zhou, et al. (2021) Adaptive immunity induces mutualism between commensal eukaryotes. Nature 596: 114-118.
-
Tso GHW, Calderon JAR, Tan ASM, XiaoHui Sem, Le GTT, et al. (2018) Experimental evolution of a fungal pathogen into a gut symbiont. Science 362(6414): 589- 595.
-
D Andes, J Nett, P Oschel, R Albrecht, K Marchillo, et al. (2004) Development and characterization of an in vivo central venous catheter Candida albicans biofilm model. Infect Immun 72(10): 6023-6031.
-
Lohse MB, Gulati M, Johnson AD, Nobile CJ (2017) Development and regulation of single- and multi-species Candida albicans biofilms. Nat Rev Microbiol 16: 19-31.
-
Dhamgaye S, Qu Y, Peleg AY (2016) Polymicrobial infections involving clinically relevant Gram-negative bacteria and fungi. Cell Microbiol 18(12): 1716-1722.
-
Wittaker Leclair L, Hogan DA (2010) Mixed bacterial- fungal infections in the CF respiratory tract. Med Mycol 48: 125-132.
-
O’Donnell LE, Millhouse E, Leighann S, Kean R, Malcolm J, et al. (2015) Polymicrobial Candida biofilms: friends and foe in the oral cavity. FEMS Yeast Res 15 15(7).
-
Xu H, Sobue T, Thompson A, Xie Z, Poon K, et al. (2014) Streptococcal co-infection augments Candida pathogenicity by amplifying the mucosal inflammatory response. Cell Microbiol 16: 214-231.
-
Morse DJ, Wilson MJ, Wei X, Bradshaw DJ, Lewis MAO, et al. (2019) Modulation of Candida albicans virulence in in vitro biofilms by oral bacteria. Lett Appl Microbiol 68(4): 337-343.
-
Jenkinson HF, Lala HC, Shepherd MG (1990) Coaggregation of Streptococcus sanguis and other streptococci with Candida albicans. Infect. Immun 58(5):1429-1436.
-
Bamford CV, Anita M, Nobbs AH, Dutton CL, Vickerman MM, et al. (2009) Streptococcus gordonii modulates Candida albicans biofilm formation through intergeneric communication. Infect Immun 77 (9): 3696-3704.
-
Jack AA, Daniels ED, Jepson MA, Vickerman MM, Lamoni RJ, et al. (2015) Streptococcus gordonii comCDE (competence) operon modulates biofilm formation with Candida albicans. Microbiology 161(2): 411-421.
-
Morse DJ, Wilson MJ, Wei X, Bradshaw DJ, Lewis DW, et al. (2019) Modulation of Candida albicans virulence in in vitro biofilms by oral bacteria. Let. Appl Microbiol 68(4): 337-343.
-
Bachtiar EW, Bachtiar BM, Jarosz LM, Amir RL, Sunarto H, et al. (2014) AI-2 of Aggregatibacter actinomycetemcomitans inhibits Candida albicans biofilm formation. Front Cell Infect Microbiol 4: 1-8.
-
Holcombe LJ, McAlester G, Munro CA, Enjalbert B, Brown AJP, et al. (2010) _Pseudomonas aeruginosa_ secreted factors impair biofilm development in Candida albicans. Microbiology 156(5): 1476-1485.
-
Man WH, Piters SWAA, Bogaert D (2017) The microbiota of the respiratory tract: gatekeeper to respiratory health. Nat Rev Microbiol 15: 259-270.
-
McAlpine J, Robbins N, Cowen LE (2022) Bacterial- fungal interactions and their impact on microbial pathogenesis. Mol Ecol 32(10): 2565-2581.
-
Herricks JR, Hotez PJ, Wanga V, Coffeng LE, Haagsma JA, et al. (2017) The global burden of disease study 2013: what does it mean for the NTDs?. PLoS Neglected Tropical Diseases 11(8): e0005424.
-
Fumagalli M, Pozzoli U, Cagliani R, et al. (2009) Parasites represent a major selective force for interleukin genes and shape the genetic predisposition to autoimmune conditions. Journal of Experimental Medicine 206(6): 1395-1408.
-
McSorley HJ, Maizels RM (2012) Helminth infections and host immune regulation. Clinical Microbiology Reviews 25(4): 585-608.
-
Wammes L, Mpairwe H, Elliott A, Yazdanbakhsh M (2014) Helminth therapy or elimination: epidemiological, immunological, and clinical considerations. The Lancet Infectious Diseases 14(11): 1150-1162.
-
Ryan SM, Eichenberger RM, Ruscher R, Giacomin PR, Loukas A (2020) Harnessing helminth-driven immunoregulation in the search for novel therapeutic modalities. PLoS Pathogens 16(5): e1008508.
-
Reese T, Wakeman BS, Choi HS, Hufford MM, Huang SC, et al. (2014) Helminth infection reactivates latent γ-herpesvirus via cytokine competition at a viral promoter. Science 345(6196): 573-577.
-
Boyle WF, Riggs JL, Oshiro LS, Lennette EH (1973) Electron microscopic identification of papova viruses in laryngeal papillomata. Laryngoscope 83(7): 1102-1108.
-
Arnold W (1976) Tubular forms of papova viruses in human laryngeal papilloma. Arch Otorhinolaryngol 225(1): 15-19.
-
Mackenzie M (1871) Essays on growths in the larynx. London J & A Churchill.
-
Vetskova EK, Muhtarova MN, Avramov TI, Stefanova TR, Chalakov IJ, et al. (2013) Immunomodulatory effects of BCG in patients with recurrent respiratory papillomatosis. Folia Med 55(1):49-54.
-
Avramov T, Vetckova E, Nikolova M, Valev D, Manolova A, et al. (2014) Therapeutic approaches to the treatment of recurrent respiratory papillomatosis of the aerodigestive tract (clinical study). Biotechnology & Biotechnological Equipment 28(4): 668-673.
-
Israr M, DeVoti JA, Lam F, Abramson AL, Steinberg BM, et al. (2020) Altered monocyte and langerhans cell immunity in pathints with Recurrent Respiratory Papillomatosis (RRP). Front Immun 11.
-
Brown RL, Sequeira RP, Clarke TB (2017) The microbiota protects against respiratory infection via GM-CSF signaling. Nat Commun 8: 1512.
-
Cait A, Hughes MR, Antignano F, Cait J, Dimitriu PA, et al. (2017) Microbiome-driven allergic lung inflammation is ameliorated by short-chain fatty acids. Mucosal Immunol 11: 785-795.
-
Flynn M, Dooley J (2021) The microbiome of the nasopharynx. J Med Microbiol 70(6).
-
Man WH, Clerc M, Piters SWAA, Houten MA, Chu MLJN, et al. (2019) Loss of microbial topography between oral and nasopharyngeal microbiota and development of respiratory infections early in life. Am J Respir Crit Care Med 200(6): 760-770.
-
Wylie K, Mihindukulasuriya K, Sodergren E, Weinstock G, Storch G (2012) Sequence analysis of the human virome in febrile and afebrile children. PLOS ONE 7: e27735.
-
Mahy BWJ, Collier L (1998) Topley and Wilson’s Microbiology and Microbial Infection. Virology, UK, 1.
-
Migone TS, Subramanian M, Zhong J, Healy ML, Corey AI, et al. (2009) Raxibacumab for the treatment of inhalational anthrax. N Engl J Med 361(2): 135-144.
-
Shenoy A, Gottlieb A (2019) Probiotics for oral and vulvovaginal candidiasis: A review. Dermatol Ther 32: e12970.
-
Ichinohe T, Lee HK, Ogura Y, Flavell R, Iwasaki A (2009) Inflammasome recognition of influenza virus is essential for adaptive immune responses. J Exp Med 206(1): 79-87.
-
Thomas PG, Dash P, Aldridge JR, Ellebedy AH, Reynolds C, et al. (2009) The intracellular sensor NLRP3 mediates key innate and healing responses to influenza A virus via the regulation of caspase-1. Immunity 30(4): 566-575.
-
Sarvestani ST, McAuley JL (2017) The role of the NLRP3 inflammasome in regulation of antiviral responses to influenza A virus infection. Antiviral Res 148: 32-42.
-
McAuley JL, Tate MD, Kenzie-Kludas CJ, Pinar A, et al. (2013) Activation of the NLRP3 inflammasome by IAV virulence protein PB1-F2 contributes to severe pathophysiology and disease. PLoS Pathog 9: e1003392.
-
Chung WC, Kang HR, Yoon H, Kang SJ, Ting JP, et al. (2015) Influenza A virus NS1 protein inhibits the NLRP3 inflammasome. PLoS One 10: e0126456.
-
Pinar A, Dowling JK, Bitto NJ, Robertson AA, Latz E, et al. (2017) PB1-F2 peptide derived from avian influenza A virus H7N9 induces inflammation via activation of the NLRP3 inflammasome. J Biol Chem 292(3): 826-836.
-
Chen IY, Moriyama M, Chang MF, Ichinohe T (2019) Severe acute respiratory syndrome coronavirus viroporin 3a activates the NLRP3 inflammasome. Front Microbiol 10: 50.
-
Bustamante-Marin XM, Ostrowski LE (2017) Cilia and mucociliary clearance. Cold Spring Harb Perspect Biol 9: a028241.
-
Paules C, Subbarao K (2017) Influenza. Lancet 390: 697-708.
-
Xiong X, Martin SR, Haire LF, Wharton SA, Daniels RS, et al. (2013) Receptor binding by an H7N9 influenza virus from humans. Nature 499: 496-499.
-
Riel D, Munster VJ, de Wit E, Rimmelzwaan GF, Fouchier RA, et al. (2006) H5N1 virus attachment to lower respiratory tract. Science 312(5772): 399.
-
Nicholls JM, Chan MC, Chan WY, Wong HK, Cheung CY, et al. (2007) Tropism of avian influenza A (H5N1) in the upper and lower respiratory tract. Nat Med 13: 147- 149.
-
Subbarao K, Mahanty S (2020) Respiratory virus infections: understanding COVID-19. Immunity 52(6): 905-909.
-
VKovski P, Kratzel A, Steiner S, Stalder H, Thiel V (2020) Coronavirus biology and replication: implications for SARS-CoV-2. Nat Rev Microbiol 19: 155-170.
-
Vanarsdall AL, Johnson DC (2012) Human cytomegalovirus entry into cells. Curr Opin Virol 2(1): 37-42.
-
Bohmwald K, Espinoza JA, Pulgar RA, Jara EL, Kalergis AM (2017) Functional impairment of mononuclear phagocyte system by the human respiratory syncytial virus. Front Immunol 8: 1643.
-
Grant RA, Morales-Nebreda L, Markov NS, Swaminathan S, Querrey M, et al. (2021) Circuits between infected macrophages and T cells in SARS- CoV-2 pneumonia. Nature 590: 635-641.
-
Anabelle W, Laura ABG, Elizabeth G, Michael B, Sarah CK, et al. (2023) REVIEW The interactions of SARS- CoV-2 with cocirculating pathogens: Epidemiological implications and current knowledge gaps. PLOS Pathogens.
-
Kitchen A, Shackelton LA, Holmes EC (2011) Family level phylogenies reveal modes of macroevolution in RNA viruses. Proc Natl Acad Sci 108(1): 238-243.
-
Hausler WJ, Sussman M (1998) Topley and Wilson’s Microbiology and Microbial Infection. In: 9th (Edn.), Bacterial Infections. Arnold, London, United Kingdom 3.
-
Cai X, Wang Q, Lin G, Cai, Lin C et al. (2014) Respiratory virus infections among children in South China. J Med Virol 86(7): 1249-1255.
-
Huang IC, Li W, Sui J, Marasco W, Choe H, et al. (2008) Influenza A virus neuraminidase limits viral superinfection. J Virol 82(10): 4834-4843.
-
Laurie KL, Horman W, Carolan LA, Chan KF, Layton D, et al. (2018) Evidence for viral interference and crossreactive protective immunity between influenza B virus lineages. J Infect Dis 217(4): 548-559.
-
Chan KF, Carolan LA, Korenkov D, Druce J, McCaw J, et al. (2018) Investigating viral interference between influenza A virus and human respiratory syncytial virus in a ferret model of infection. J Infect Dis 218(3): 406- 417.
-
Schneider WM, Chevillotte MD, Rice CM (2014) Interferon stimulated genes: a complex web of host defenses. Annu Rev Immunol 32: 513-545.
-
Voroshilova MK (1989) Potential use of nonpathogenic enteroviruses for control of human disease. In: Melnick JL (Eds.), Progress in medical virology. Basel, Karger 36: 191-202.
-
Chumakov MP, Voroshilova MK, Antsupova AS, Boiko VM, Blinova MI, et al. (1992) Live enteroviral vaccines for the emergency nonspecific prevention of mass respiratory diseases during fall-winter epidemics of influenza and acute respiratory diseases. Zh Mikrobiol Epidemiol Immunobiol (11-12): 37-40.
-
Shinjoh M, Omoe K, Saito N, Matsuo N, Nerome K (2000) _In vitro_ growth profiles of respiratory syncytial virus in the presence of influenza virus. Acta Virol 44(2): 91-97.
-
Piret J, Boivin G (2022) Viral interference between respiratory viruses, Emerg Infect Dis 28(2): 273-281.
-
Piret J, Boivin G (2022) Viral interference between respiratory viruses. Emerg Infect Dis 28(2): 273-281.
-
Hausler WJ, Sussman M (1998) Topley and Wilson’s Microbiology and Microbial Infection. In: 9th (Edn.), Bacterial Infections. Arnold, London, United Kingdom 3.
-
Nickbakhsh S, Mair C, Matthews L, Reeve R, Johnson PCD, et al. (2019) Virus-Virus interactions impact the population dynamics of influenza and common cold. Proc of Nat Acad of Science 116(52): 27142-27150.
-
Rott R., Klenk HD, Nagai Y, Tashiro M (1995) Influenza viruses, cell enzymes and pathogenicity. Am J Respir Crit Care Med 152(4): S16-S19.
-
Tashiro M, Ciborowski P, Klenk H, Pulverer G, Rott R (1987) Role of staphylococcus protease in the development of influenza pneumonia. Nature 325: 536-
-
Tashiro M, Ciborowski P, Reinacher M, Pulverer G, Klenk HD, et al. (1987) Synergistic role of staphylococcal proteases in the induction of influenza virus pathogenicity. Virology 157(2): 421-430.
-
Akaike T, Molla A, Ando M, Araki S, Maeda H (1989) Molecular mechanism of complex infection by bacteria and virus analyzed by a model using serratial protease and influenza virus in mice. J Virol 63(5): 2252-2259.
-
Kilbourne ED (1987) Influenza. Plenum Publishing Corp, New York, N.Y.
-
Schwarzmann SW, Adler JL, Sullivan RJ, Marine WM (1971) Bacterial pneumonia during the Hong Kong influenza epidemic of 1968-69. Arch Intern Med 127(6): 1037-1041.
-
Taubenberger JK, Reid AH, Fanning TG (2000) The 1918 influenza virus: a killer comes into view. Virology 274: 241-245.
-
Cartwright KAV, Jones DM, Smith AJ, Stuart JM, Kaesmarski EB, et al. (1991) Influenza A and meningococcal disease. Lancet 338(8766): 554-557.
-
Kim PE, Musher DM, Glezen WP, Rodriguez-Barradas MC, Nahm WK, et al. (1996) Association of invasive pneumococcal disease with season, atmospheric conditions, air pollution and isolation of respiratory viruses. Clin Infect Dis 22(1): 100-106.
-
Reynolds HY, Root EK (1991) Bronchiectasis and broncholithiasis. In: Wilson JD, Braunwald E, et al. (Eds.), Harrison’s Principles of Internal Medicine, McGraw-Hill, New York, NY, pp: 1069-1071.
-
Tristam D, Hicks W, Hard R (1998) Respiratory syncytial virus and human bronchial epithelium. Arch Otolaryngol Head Neck Surg 124(7): 777-783.
-
Walsh J, Dietlein L, Low F, Burch G, Mogabgab W (1960) Bronchotracheal response in human influenza. Arch Intern Med 108(3): 376-388.
-
Puchelle E, Zahm JM, Tournier JM, Coraux C (2006) Airway epithelial repair, regeneration, and remodeling after injury in chronic obstructive pulmonary disease. Proc Am Thorac Soc 3(8): 726-733.
-
Liu H, Lee MJ, Solis NV, Phan QT, Swidergall M, et al. (2016) Aspergillus fumigatus CalA binds to integrin alpha5beta1 and mediates host cell invasion. Nat Microbiol 2: 16211.
-
Kerr SC, Fischer GJ, Sinha M, McCabe O, Palmer JM, et al. (2016) FleA expression in Aspergillus fumigatus is recognized by fucosylated structures on mucins and macrophages to prevent lung infection. PLoS Pathog 12: e1005555.
-
Cowley AC, Thornton DJ, Denning DW, Horsley A (2017) Aspergillosis and the role of mucins in cystic fibrosis. Pediatr Pulmonol 52(4): 548-555.
-
Wezensky SJ, Cramer RA (2011) Implications of hypoxic microenvironments during invasive aspergillosis. Med Mycol 49(S1): S120-S124.
-
Ami R, Lewis RE, Leventakos K, Kontoyiannis DP (2009) Aspergillus fumigatus inhibits angiogenesis through the production of gliotoxin and other secondary metabolites. Blood 114: 5393-5399.
-
Shepardson KM, Ngo LY, Aimanianda V, Latge JP, Barker BM, et al. (2013) Hypoxia enhances innate immune activation to Aspergillus fumigatus through cell wall modulation. Microbes Infect 15: 259-269.
-
Kowalski CH, Beattie SR, Fuller KK, McGurk EA, Tang YW, et al. (2016) Heterogeneity among isolates reveals that fitness in low oxygen correlates with Aspergillus fumigatus virulence. mBio 7: e01515-01516.
-
Veerdonk FL, Gresnigt MS, Romani L, Netea MG, Latge JP (2017) Aspergillus fumigatus morphology and dynamic host interactions. Nat Rev Microbiol 15: 661- 674.
-
Pinsky DJ, Naka Y, Liao H, Oz MC, Wagner DD, et al. (1996) Hypoxia-induced exocytosis of endothelial cell Weibel-Palade bodies. A mechanism for rapid neutrophil recruitment after cardiac preservation. J Clin Invest 97: 493-500.
-
Hirano T, Kurono Y, Ichimiya I, Suzuki M, Mogi G (1999) Effects of influenza A virus on lectin-binding patterns in murine nasopharyngeal mucosa and on bacterial colonization. Otolaryngol. Head Neck Surg 121: 616-621.
-
Sanford BA, Shelokov A, Ramsay MA (1978) Bacterial adherence to virus infected cells: a cell culture model of bacteria super-infection. J Infect Dis 137: 176-181.
-
Genovese A, Bouvet JP, Florio G, Schummert B, Bjorck L, et al. (2000) Bacterial immunoglobulin superantigen proteins A and L activate human heart mast cells by interacting with immunoglobulin E. Infect Immun 68: 5517-5524.
-
Komisar J, Rivera J, Vega A, Tseng J (1992) Effects of staphylococcal enterotoxin B on rodent mast cells. Infect. Immun 60: 2969-2975.
-
Ono HK, Nishizawa M, Yamamoto Y, Hu DL, Nakane A, et al. (2012) Submucosal mast cells in the gastrointestinal tract are a target of staphylococcal enterotoxin type A. FEMS Immunol Med Microbiol 64: 392-402.
-
Kobayashi M, Kitano T, Nishiyama S, Sanjo H, Onozaki K, et al. (2019) Staphylococcal superantigen- like 12 activates murine bone marrow derived mast cells. Biochem. Biophys Res Commun 511: 350-355.
-
Sussman M (1998) Eds., Topley and Wilson’s Microbiology and Microbial Infection. Bacterial Infections. In: 9th (Edn.), Arnold, London, United Kingdom.
-
Austin RM, Daniels CA (1987) The role of protein A in attachment of staphylococci to influenza-infected cells. Lab Investig 39: 128-132.
-
Raza MW, Blackwell CC, Ogilvie MM, Saadi AT, Stewart J, et al. (1994) Evidence for the role of glycoprotein-G of respiratory syncytial virus in binding of Neisseria meningiditis to HEp-2 cells. FEMS Immunol. Med Microbiol 10: 25-30.
-
Larson HE, Parry RP, Gilchrist C, Luquetti A, Tyrrell DAJ (1977) Influenza viruses and staphylococci in vitro: some interactions with polymorphonuclear and epithelial cells. Br J Exp Pathol 58: 281-292.
-
Warr GA, Jakab GJ, Chan TW, Tsan MF (1979) Effects of viral pneumonia on lung macrophage lysosomal enzymes. Infect Immun 24: 577-579.
-
Pang G, Clancy R, Gong M, Ortega M, Ren ZG, et al. (2000) Influenza virus inhibits lysozyme secretion by sputum neutrophils in subjects with chronic bronchial sepsis. Am J Respir Crit Care Med 161: 718-722.
-
Ogunbiyi PO, Black WD, Eyre P (1988) Parainfluenza-3 virus-induced enhancement of histamin release from calf lung mast cells-effect of levamisole. J Vet Pharmacol Ther 11: 338-344.
-
Allen IC, Scull MA, Moore CB, Holl EK, TeKippe E, et al. (2009) The NLRP3 inflammasome mediates in vivo innate immunity to influenza A virus through recognition of viral RNA. Immunity 30: 556-565.
-
Bansal S, Yajjala VK, Bauer C, Sun K (2018) IL-1 signaling prevents alveolar macrophage depletion during influenza and _Streptococcus pneumoniae_ coinfection. J Immunol 200: 1425-1433.
-
Cervantes L, Pampena MB, Meng W, Rosenfeld AM, Ittner CAG, et al. (2020) Comprehensive mapping of immune perturbations associated with severe COVID-19. Sci Immunol 5: eabd 7114.
-
Bourboulis EJ, Netea MG, Rovina N, Akinosoglou K, Antoniadou A, et al. (2020) Complex immune dysregulation in COVID-19 patients with severe respiratory failure. Cell Host Microbe 27: 992-1000.
-
Arunachalam PS, Wimmers F, Mok CKP, Perera R, Scott M, et al. (2020) Systems biological assessment of immunity to mild versus severe COVID-19 infection in humans. Science 369: 1210-1220.
-
Wilk AJ, Rustagi A, Zhao NQ, Roque J, Colon GJ, et al. (2020) A single-cell atlas of the peripheral immune response in patients with severe COVID-19. Nat Med 26: 1070-1076.
-
Schrepping J, Reusch N, Paclik D, Bassler K, Schlickeiser S, et al. (2020) Severe COVID-19 is marked by a dysregulated myeloid cell compartment. Cell 182: 1419-1440.
-
Silvin A, Chapuis N, Dunsmore G, Goubet AG, Dubuisson A, et al. (2020) Elevated calprotectin and abnormal myeloid cell subsets discriminate severe from mild COVID-19. Cell 182: 1401-1418.
-
Su Y, Chen D, Yuan D, Lausted C, Choi J, et al. (2020) Multi-omics resolves a sharp disease-state shift between mild and moderate COVID-19. Cell 183: 1479-1495.
-
Kreutmair S, Unger S, Nunez NG, Ingelfinger F, Alberti C, et al. (2021) Distinct immunological signatures discriminate severe COVID-19 from non-SARS-CoV-2- driven critical pneumonia. Immunity 54: 1578-1593.
-
Wilk AJ, Lee MJ, Wei B, Parks B, Pi R, et al. (2021) Multi-omic profiling reveals widespread dysregulation of innate immunity and hematopoiesis in COVID-19. J Exp Med 218: e2021082.
-
Kvedaraite E, Hertwig L, Sinha I, Ponzetta A, Myrberg I, et al. (2021) Major alterations in the mononuclear phagocyte landscape associated with COVID-19 severity. Proc Natl Acad Sci USA 118: e2018587118.
-
Szabo PA, Dogra P, Gray JI, Wells SB, Connors TJ, et al. (2021) Longitudinal profiling of respiratory and systemic immune responses reveals myeloid cell-driven lung inflammation in severe COVID-19. Immunity 54: 797-814.e6.
-
Thompson EA, Cascino K, Ordonez AA, Zhou W, Vaghasia A, et al. (2021) Metabolic programs define dysfunctional immune responses in severe COVID-19 patients. Cell Rep 34: 108863.
-
Wheeler JL, Martin KC, Lawrence BP (2013) Novel cellular targets of AhR underlie alterations in neutrophilic inflammation and inducible nitric oxide synthase expression during influenza virus infection. J Immunol 190: 659-668.
-
Teske S, Bohn AA, Regal JF, Neumiller JJ, Lawrence BP (2005) Activation of the aryl hydrocarbon receptor increases pulmonary neutrophilia and diminishes host resistance to influenza A virus. Am J Physiol Lung Cell Mol Physiol 289: L111-L124.
-
White MR, Tecle T, Crouch EC, Hartshorn KL (2007) Impact of neutrophils on antiviral activity of human bronchoalveolar lavage fluid. Am J Physiol Lung Cell Mol Physiol 293: L1293-L1299.
-
Bradley LM, Douglass MF, Chatterjee D, Akira S, Baaten BJ (2012) Matrix metalloprotease 9 mediates neutrophil migration into the airways in response to influenza virus-induced toll-like receptor signaling. PLoS Pathog 8: e1002641.
-
Zhang J, Liu J, Yuan Y, Huang F, Ma R, et al. (2020) Two waves of pro-inflammatory factors are released during the influenza A virus (IAV)-driven pulmonary immunopathogenesis. PLoS Pathog 16: e1008334.
-
Ellis GT, Davidson S, Crotta S, Branzk N, Papayannopoulos V, et al. (2015) TRAIL+ monocytes and monocyte-related cells cause lung damage and thereby increase susceptibility to influenza-_Streptococcus_ _pneumoniae_ coinfection. EMBO Rep 16: 1203-1218.
-
Raza MW, Blackwell CC, Elton RA, Weir DM (2000) Bactericidal activity of a monocytic cell line (THP-1) against common respiratory tract bacterial pathogens is depressed after infection with respiratory syncytial virus. J Med Microbiol 49: 227-233.
-
Channappanavar R, Fehr AR, Vijay R, Mack M, Zhao J, et al. (2016) Dysregulated type I interferon and inflammatory monocyte-macrophage responses cause lethal pneumonia in SARS-CoV-infected mice. Cell Host Microbe 19:181-193.
-
Channappanavar R, Perlman S (2017) Pathogenic human coronavirus infections: causes and consequences of cytokine storm and immunopathology. Semin Immunopathol 39: 529-539.
-
Israelow B, Song E, Mao T, Lu P, Meir A, et al. (2020) Mouse model of SARS-CoV-2 reveals inflammatory role of type I interferon signaling. J Exp Med 217: e20201241.
-
Kamphuis E, Junt T, Waibler Z, Forster R, Kalinke U (2006) Type I interferons directly regulate lymphocyte recirculation and cause transient blood lymphopenia. Blood 108: 3253-3261.
-
Galani IE, Triantafyllia V, Eleminiadou EE, Koltsida O, Stavropoulos A, et al. (2017) Interferon-lambda mediates non-redundant front-line antiviral protection against influenza virus infection without compromising host fitness. Immunity 46: 875-890.
-
Bellelli V, Ettorre G, Celani L, Borrazzo C, Ceccarelli G, et al. (2019) Clinical significance of lymphocytopenia in patients hospitalized with pneumonia caused by influenza virus. Crit Care 23: 330.
-
Gooskens J, Jonges M, Claas EC, Meijer A, Kroes AC (2009) Prolonged influenza virus infection during lymphocytopenia and frequent detection of drug- resistant viruses. J Infect Dis 199: 1435-1441.
-
Tan L, Wang Q, Zhang D, Ding J, Huang Q, et al. (2020) Lymphopenia predicts disease severity of COVID-19: a descriptive and predictive study. Signal Transduct Target Ther 5: 33.
-
Rich HE, McCourt CC, Zheng WQ, McHugh KJ, Robinson KM, et al. (2019) Interferon lambda inhibits bacterial uptake during influenza superinfection. Infect Immun 87: e00114-e00119.
-
Planet PJ, Parker D, Cohen TS, Smith H, Leon JD, et al. (2016) Lambda interferon restructures the nasal microbiome and increases susceptibility to _Staphylococcus aureus_ superinfection. mBio 7: e01939-e01915.
-
Sposito B, Broggi A, Pandolfi L, Crotta S, Clementi N, et al. (2021) The interferon landscape along the respiratory tract impacts the severity of COVID-19. Cell 184: 4953-4968.
-
Loske J, Rohmel J, Lukassen S, Stricker S, Magalhaes VG, et al. (2022) Pre-activated antiviral innate immunity in the upper airways controls early SARS-CoV-2 infection in children. Nat Biotechnol.
-
Rynda-Apple A, Harmsen A, Erickson AS, Larson K, Morton RV, et al. (2014) Regulation of IFN-gamma by IL-13 dictates susceptibility to secondary postinfluenza MRSA pneumonia. Eur J Immunol 44: 3263-3272.
-
Sun K, Metzger DW (2008) Inhibition of pulmonary antibacterial defense by interferon-gamma during recovery from influenza infection. Nat Med 14: 558-564.
-
Li N, Fan X, Xu M, Zhou Y, Wang B (2020) Flu virus attenuates memory clearance of pneumococcus via IFN- gamma-dependent Th17 and independent antibody mechanisms. I Science 23: 101767
-
Hadjadj J, Yatim N, Barnabei L, Corneau A, Boussier J, et al. (2020) Impaired type I interferon activity and inflammatory responses in severe COVID-19 patients. Science 369: 718-724.
-
Arunachalam PS, Wimmers F, Mok CKP, Perera R, Scott M, et al. (2020) Systems biological assessment of immunity to mild versus severe COVID-19 infection in humans. Science 369: 1210-1220.
-
Combes AJ, Courau T, Kuhn NF, Hu KH, Ray A, et al. (2021) Global absence and targeting of protective immune states in severe COVID-19. Nature 591: 124- 130.
-
Severa M, Diotti RA, Etna MP, Rizzo F, Fiore S, et al. (2021) Differential plasmacytoid dendritic cell phenotype and type I Interferon response in asymptomatic and severe COVID-19 infection. PLoS Pathog 17: e1009878.
-
Onodi F, Bonnet-Madin L, Meertens L, Karpf L, Poirot J, et al. (2021) SARS-CoV-2 induces human plasmacytoid predendritic cell diversification via UNC93B and IRAK4. J Exp Med 218: e20201387.
-
Cervantes-Barragan L, Vanderheiden A, Royer CJ, Davis-Gardner ME, Ralfs P, et al. (2021) Plasmacytoid dendritic cells produce type I interferon and reduce viral replication in airway epithelial cells after SARS-CoV-2 infection. BioRxiv.
-
Sadaka C, Marloie-Provost MA, Soumelis V, Benaroch P (2009) Developmental regulation of MHC II expression and transport in human plasmacytoid-derived dendritic cells. Blood 113: 2127-2135.
-
Smed-Sorensen A, Chalouni C, Chatterjee B, Cohn L, Blattmann P, et al. (2012) Influenza A virus infection of human primary dendritic cells impairs their ability to cross-present antigen to CD8 T cells. PLoS Pathog 8: e1002572.
-
Frleta D, Yu CI, Klechevsky E, Flamar AL, Zurawski G, et al. (2009) Influenza virus and poly(I:C) inhibit MHC class I-restricted presentation of cell-associated antigens derived from infected dead cells captured by human dendritic cells. J Immunol 182: 2766-2776.
-
Zhou R, To KK, Wong YC, Liu L, Zhou B, et al. (2020) Acute SARS-CoV-2 infection impairs dendritic cell and T cell responses. Immunity 10: 1016.
-
Rezinciuc S, Bezavada L, Bahadoran A, Duan S, Wang R, et al. (2020) Dynamic metabolic reprogramming in dendritic cells: an early response to influenza infection that is essential for effector function. BioRxiv.
-
Barton ES, White DW, Cathelyn JS (2007) Herpesvirus latency confers symbiotic protection from bacterial infection. Nature 447(7142): 326-329.
-
Yao Y, Liu N, Zhou Z, Shi L (2019) Influence of ERAP1 and ERAP2 gene polymorphisms on disease susceptibility in different populations. Hum Immunol 80: 325-334.
-
Saulle I, Vicentini C, Clerici M, Blasin M (2020) An overview on ERAP roles in infectious diseases. Cells 9: 720.
-
Laufer V (2019) Genetic influences on susceptibility to rheumatoid arthritis in African- Americans. Hum Mol Genet 28: 858-874.
-
Wang Y (2021) Identification of 38 novel loci for systemic lupus erythematosus and genetic heterogeneity between ancestral groups. Nat Commun 12: 771.
-
Klunk J (2022) Evolution of immune genes is associated with the Black Death. Nature 611: 312-319.
-
Sorden SD, Castleman WL (1991) Brown Norway rats are high responders to bronchiolitis, pneumonia, and bronchiolar mastocytosis induced by parainfluenza virus. Exp Lung Res 17: 1025-1045.
-
Dakhama A, Lee YM, Ohnishi H, Jing X, Balhorn A, et al. (2009) Virus-specific IgE enhances airway responsiveness on reinfection with respiratory syncytial virus in newborn mice. J Allergy Clin Immunol 123: 138- 145.
-
Al-Afif A (2015) Respiratory syncytial virus infection of primary human mast cells induced the selective production of type I interferons, CXCL10 and CCL4. J Allergy Clin Immunol 136: 1346-1354.
-
Marcet CW (2013) Limited replication of influenza A virus in human mast cells. Immunol Res 56: 32-43.
-
Liu H, Tan J, Liu J, Feng H, Pan D (2019) Altered mast cell activity in response to rhinovirus infection provides novel insight into asthma. J Asthma 18: 1-9.
-
Akoto C, Davies DE, Swindle EJ (2017) Mast cells are permissive for rhinovirus replication: Potential implications for asthma exacerbations. Clin Exp Allergy 47: 351-360.
-
Hosoda M, Yamaya M, Suzuki T, Yamada N, Kamanaka M, et al. (2002) Effects of rhinovirus infection on histamine and cytokine production by cell lines from human mast cells and basophils. J Immunol 169: 1482- 1491.
-
Kilbourne ED (1987) Influenza. Plenum Publishing Corp, New York.
-
Taubenberger JK, Reid AH, Fanning TG (2000) The 1918 influenza virus: a killer comes into view. Virology 274: 241-245.
-
Cartwright KAV, Jones DM, Smith AJ, Stuart JM, Kaesmarski EB, Palmer SR (1991) Influenza A and meningococcal disease. Lancet 338(8766): 554-557.
-
Kim PE, Musher DM, Glezen WP, Rodriguez-Barradas MC, Nahm WK, et al. (1996) Association of invasive pneumococcal disease with season, atmospheric conditions, air pollution and isolation of respiratory viruses. Clin Infect Dis 22(1): 100-106.
-
Fiore AE, Iverson C, Messmer T, Erdman D, Lett SM, et al. (1998) Outbreak of pneumonia in a long term care facility: antecedent human parainfluenza virus 1 infection may predispose to bacterial pneumonia. J Am Geriatr Soc 46(9): 1112-1117.
-
Makela MJ, Puhakka T, Ruuskanen O, Leinonen M, Saikku P, et al. (1998) Viruses and bacteria in the etiology of the common cold. J Clin Microbiol 36(2): 539-542.
-
Klenk HD, Rott R, Orlich M (1977) Further studies on the activation of influenza virus by proteolytic cleavage of the haemagglutinin. J Gen Virol 36(1): 151-161.
-
Scheiblauer H, Reinacher M, Toshiro M, Rott R (1992) Interaction between bacteria and influenza A virus in the development of influenza pneumonia. J Infect Dis 166(4): 783-791.
-
Moore H, Jacoby P, Taylor A, Harnett G, Bowman J (2010) The interaction between respiratory viruses and pathogenic bacteria in upper respiratory tract of asymptomatic aboriginal and nonaboriginal children. Ped Infect Dis J (29)60: 540-545.
-
Barker W, Mullooly J (1982) Pneumonia and influenza deaths during epidemics: implications for prevention. Arch Intern Med 142(1): 85-89.
-
Floret D (1997) Virus-bacteria co-infections. _Arch_ _Pediatr_ 4: 1119-1124.
-
Heiskanen-Kosma T, Korppi M, Jokinen C, Kurki S, Heiskanen L, et al. (1998) Etiology of childhood pneumonia: serologic results of a prospective, population based study. Pediatr Infect Dis J 17(11): 986-991.
-
Nelson W, Hopkins R, Roe M, Glode M (1986) Simultaneous infection with _Bordetella pertussis_ and respiratory syncytial virus in hospitalized children. Pediatr Infect Dis J 5(5): 540-544.
-
Gatchalian S, Halonen P, Lucero M, Sombrero L, Paladin F, et al. (1998) Co-infection is common in measles-associated pneumonia. Pediatr Infect Dis J 17(2): 89-93.
-
Taubenberger J, Reid A, Fanning T (2000) The 1918 influenza virus: a killer comes into view. Virology 274: 241-245.
-
Howard C, Stott E, Taylor G (1978) The effect of pneumonia induced in mice with Mycoplasma pulmonis on resistance to subsequent bacterial infection and the effect of respiratory infections with Sendai virus on resistance of mice to Mycoplasma pulmonis. J Gen Microbiol 109(1): 79-87.
-
Klein J, Green G, Tiles J, Kass E, Finland M (1969) Effect of intranasal virus infection on antibacterial activity of mouse lung. J Infect Dis 119(1): 43-50.
-
Hamilton J, Overall J (1978) Synergistic infection with murine cytomegalovirus and _Pseudomonas_ _aeruginosa_ in mice. J Infect Dis 137(6): 775-782.
-
Blackwell CC, MacKenzie DAC, James VS, Elton RA, Zorgani AA, et al. (1999) Toxigenic bacteria and sudden infant death syndrome (SIDS): nasopharyngeal flora during the first year of life. FEMS Immunol Med Microbiol 25(1-2): 51-58.
-
Telford DR, Morris JA, Hughes P, Conway AR, Lee S, et al. (1989) The nasopharyngeal bacterial flora in the sudden infant death syndrome. J Infect 18(2): 125-130.
-
Regev-Yochay G, Dagan R, Raz M, et al. (2004) Association between carriage of _Streptococcus_ _pneumoniae_ and _Staphylococcus aureus_ in children. JAMA 292(6):716-720.
-
Bogaert D, De Groot R, Hermans PWM (2004) _Streptococcus pneumoniae_ colonisation: the key to pneumococcal disease_._ Lancet Infect Dis 4(3): 144-154.
-
Watson K, Carville K, Bowman J, et al. (2006) Upper respiratory tract bacterial carriage in Aboriginal and non-Aboriginal children in semi-arid area of Western Australia. Pediatr Infect Dis J 25(9): 782-790.
-
Chan WY, Cohen J, Brown J (2016) The new first-line defense: the potential of nasopharyngeal colonization in vaccine strategies. Vaccine Dev Ther 6: 47-57.
-
Engen SA, Valen Rukke H, Becattini S, Jarrossay D, Blix IJ, et al. (2014) The oral commensal Streptococcus mitis shows a mixed memory Th cell signature that is similar to and cross-reactive with _Streptococcus_ _pneumoniae_. PLoS One 9: e104306-e104309.
-
Revai K, Mamidi D, Chonmaitree T (2008) Association of nasopharyngeal bacterial colonization during upper respiratory tract infection and the development of acute otitis media. Clin Infect Dis 46(5): e34-e37.
-
Bogaert D, De Groot R, Hermans PWM (2004) _Streptococcus pneumoniae_ colonisation: the key to pneumococcal disease. Lancet Infect Dis 4(3): 144-154.
-
Watson K, Carville K, Bowman J, Peter J, Victor RT, et al. (2006) Upper respiratory tract bacterial carriage in Aboriginal and non-Aboriginal children in semi-arid area of Western Australia. Pediatr Infect Dis J 25(9): 782- 790.
-
Chan WY, Cohen J, Brown J (2016) The new first-line defense: the potential of nasopharyngeal colonization in vaccine strategies. Vaccine Dev Ther 6:47-57.
-
Shahid M, Tripathi T, Sobia F, Moin S, Siddiqui M, et al. (2009) Histamine, histamine receptors, and their role in immunomodulation: an updated systematic review. Open Immunol J 2(1): 9-41.
-
Gallagher T, Snell EE, Hackert ML (1989) Pyruvoyl- dependent histidine decarboxylase: active site structure and mechanistic analysis. J Biol Chem 264(21): 12737- 12743.
-
Parsons ME, Ganellin CR (2006) Histamine and its receptors. Br J Pharmacol 147(Suppl 1): S127-S135.
-
White MV, O’Mahony L, Akdis M, Akdis CA, Shahid M, et al. (1990) The role of histamine in allergic diseases. J Allergy Clin Immunol 86(4): 599-605.
-
O’Mahony L, Akdis M, Akdis CA, Shahid M, Tripathi T, et al. (2011) Regulation of the immune response and inflammation by histamine and histamine receptors. J Allergy Clin Immunol 128(6): 1153-1162.
-
Shahid M, Tripathi T, Sobia F, Moin S, Siddiqui M, et al. (2009) Histamine, histamine receptors, and their role in immunomodulation: an updated systematic review. Open Immunol J 2(1): 39-41.
-
Pishko EJ, Robertus JD, Potter KA, Shahid M, Tripathi T, et al. (1993) Site-directed alteration of three active-site residues of a pyruvoyl-dependent histidine decarboxylase. Biochemistry 32: 4943-4948.
-
Pishko EJ, Potter KA, Robertus JD, Shahid M, Tripathi T, et al. (1995) Site-directed mutagenesis of intersubunit boundary residues in histidine decarboxylase, a pH- dependent allosteric enzyme. Biochemistry 34(18): 6069-6073.
-
Alvarez MA, Moreno-Arribas MV, White MV, O’Mahony L, Akdis M, et al. (2014) The problem of biogenic amines in fermented foods and the use of potential biogenic amine-degrading microorganisms as a solution. Trends Food Sci Technol 39(2): 146-155.
-
Ladero V, Calles-Enriquez M, Fernandez MA, Alvarez MA, O’Mahony L, et al. (2010) Toxicological effects of dietary biogenic amines. Curr Nutr Food Sci 6(2): 145- 156.
-
Krulwich TA, Sachs G, Padan E, White MV, O’Mahony L, et al. (2011) Molecular aspects of bacterial pH sensing and homeostasis. Nat Rev Microbiol 9(5): 330-343.
-
Diaz M, Del Rio B, Ladero V, Redruello B, Fernández M, et al. (2020) Histamine production in Lactobacillus vaginalis improves cell survival at low pH by counteracting the acidification of the cytosol. Int J Food Microbiol 321: 108548.
-
Kohanski MA, Dwyer DJ, Collins JJ, White MV, O’Mahony L, et al. (2010) How antibiotics kill bacteria: from targets to networks. Nat Rev Microbiol 8: 423-435.
-
Mukhopadhya I, Hansen R, El-Omar EM, Hold GL, Shahid M, et al. (2012) IBD - what role do Proteobacteria play? Nat Rev Gastroenterol Hepatol 9(4): 219-230.
-
Falony G, Joossens M, Vieira-Silva S, Wang J, Darzi Y, et al. (2016) Population-level analysis of gut microbiome variation. Science 352(6285): 560-564.
-
Guarner F, Malagelada JR, Shahid M, Tripathi T, Sobia F, et al. (2003) Gut flora in health and disease. Lancet 361(9356): 512-519.
-
Calles-Enriquez M, Eriksen BH, Andersen PS, Rattray FP, Johansen AH, et al. (2010) Sequencing and transcriptional analysis of the Streptococcus thermophilus histamine biosynthesis gene cluster: factors that affect differential hdcA expression. Appl Environ Microbiol 76(18): 6231-6238.
-
Economou V, Gousia P, Kemenetzi D, Sakkas H, Papadopoulou C, et al. (2017) Microbial quality and histamine-producing microflora analysis of the ice used for fish preservation. J Food Saf 37(1): e12285.
-
Mu Q, Tavella VJ, Luo XM, Shahid M, Tripathi T, et al. (2018) Role of _Lactobacillus reuteri_ in human health and diseases. Front Microbiol 9: 757.
-
Gao C, Major A, Rendon D, Lugo M, Jackson V, et al. (2015) Histamine H2 receptor-mediated suppression of intestinal inflammation by probiotic _Lactobacillus_ _reuteri_. MBio 6(6): e01358-15.
-
Simmons SS, Isokpehi RD, Brown SD, McAllister DL, Hall CC, et al. (2011) Functional annotation analytics of Rhodopseudomonas palustris genomes. Bioinform Biol Insights 5: 115-129.
-
Oda Y, Larimer FW, Chain PSG, Malfatti S, Shin MV, et al. (2008) Multiple genome sequences reveal adaptations of a phototrophic bacterium to sediment microenvironments. Proc Natl Acad Sci U S A 105(47): 18543-18548.
-
Ceapa C, Davids M, Ritari J, Lambert J, Wels M, et al. (2016) The variable regions of Lactobacillus rhamnosus genomes reveal the dynamic evolution of metabolic and host-adaptation repertoires. Genome Biol Evol 8(6): 1889-1905.
-
Donaldson GP, Lee SM, Mazmanian SK, White MV, O’Mahony L, et al. (2016) Gut biogeography of the bacterial microbiota. Nat Rev Microbiol 14(1): 20-32.
-
Zhang J, Guo Z, Xue Z, Sun Z, Zhang M, et al. (2015) A phylo-functional core of gut microbiota in healthy young Chinese cohorts across lifestyles, geography and ethnicities. ISME J 9(9): 1979-1990.
-
Calles-Enriquez M, Eriksen BH, Andersen PS, Rattray FP, Johansen AH, et al. (2010) Sequencing and transcriptional analysis of the Streptococcus thermophilus histamine biosynthesis gene cluster: factors that affect differential hdcA expression. Appl Environ Microbiol 76(18): 6231-6238.
-
Economou V, Gousia P, Kemenetzi D, Sakkas H, Papadopoulou C, et al. (2017) Microbial quality and histamine-producing microflora analysis of the ice used for fish preservation. J Food Saf 37(1): e12285.
-
Mu Q, Tavella VJ, Luo XM, Shahid M, Tripathi T, et al. (2018) Role of _Lactobacillus reuteri_ in human health and diseases. Front Microbiol 9: 757.
-
Ceapa C, Davids M, Ritari J, Lambert J, Wels M, et al. (2016) The variable regions of Lactobacillus rhamnosus genomes reveal the dynamic evolution of metabolic and host-adaptation repertoires. Genome Biol Evol 8(6): 1889-1905.
-
Marri PR, Hao W, Golding GB, White MV, O’Mahony L, et al. (2006) Gene gain and gene loss in Streptococcus: is it driven by habitat? Mol Biol Evol 23(12): 2379-2391.
-
Stepinska M, Olszewska-Sosinska O, Lau-Dworak M, Zielnik-Jurkiewicz B, Trafny E, et al. (2013) Identification of intracellular bacteria in adenoid and tonsil tissue specimens: the efficiency of culture versus fluorescent in situ hybridization (FISH). Curr Microbiol 68: 21-29.
-
Klein EY, Van Boeckel TP, Martinez EM, Pant S, Gandra S, et al. (2018) Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc Natl Acad Sci U S A 115: E3463-E3470.
-
Neuman H, Forsythe P, Uzan A, Avni O, Koren O, et al. (2018) Antibiotics in early life: dysbiosis and the damage done. FEMS Microbiol Rev 42: 489-499.
-
Francino MP (2015) Antibiotics and the human gut microbiome: dysbiosis and accumulation of resistance. Front Microbiol 6:1543.
-
Fletcher JR, Pike CM, Parsons RJ, AJ Rivera, MH Foley, et al. (2021) Clostridoides difficile exploits toxin- mediated inflammation to alterthe host nutritional landscape and exclude competitors from the gut microbiota. Nat Commun 12: 462.
-
Logan LK, Weinstein RA (2017) The epidemiology of carbapenem-resistant Enterobacteriaceae: the impact and evolution of global menace. J Infect Dis 215(suppl_1): S28-S36.
-
Duerkop BA (2018) Bacteriophages shift the focus of the mammalian microbiota. PLoS Pathog 14: e1007310.
-
Sanders DJ, Inniss S, Sebepos-Rogers G, Rahman FZ, Smith AM (2021) The role of the microbiome in gastrointestinal inflammation. Biosci Rep 41: 3850.
-
Boling L, Cuevas DA, Grasis JA, Kang HS, Knowles B, et al. (2020) Dietary prophage inducers and antimicrobials: toward landscaping the human gut microbiome. Gut Microbes 11: 721-734.
-
Gutiérrez B, Domingo-Calap P (2020) Phage therapy in gastrointestinal diseases. Microorganisms 8: 1420.
-
Costea PI, Hildebrand F, Arumugam M, Backhed F, Blaser MJ, et al. (2018) Enterotypes in the landscape of gut microbial community composition. Nat Microbiol 3: 8-16.
-
Wu GD, Chen J, Hoffmann C, Bittinger K, Chen YY, et al. (2011) Linking long-term dietary patterns with gut microbial enterotypes. Science 334: 105-108.
-
J Walter, Heng NC, Hammes WP, Loach DM, Tannock GW, et al. (2003) Identification of _Lactobacillus reuteri_ genes specifically induced in the mouse gastrointestinal tract. Appl Environ Microbiol 69: 2044-2051.
-
Shukla SK, Ye Z, Sandberg S, Reyes I, Fritsche TR, et al. (2017) The nasal microbiota of dairy farmers is more complex than oral microbiota, reflects occupational exposure, and provides competition for staphylococci. PLoS One 12: e0183898.
-
Soren S, Mandal GP, Roy B, Samanta I, Hansda RN (2023) Assessment of Bacillus subtilis based probiotics on health and productive performance of poultry: A review. Indian J Anim Health 62: 2.
-
Giri S, Waschina S, Kaleta C, Kost C (2019) Defining division of labor in microbial communities. J Mol Biol 431: 4712-4731.
-
West SA, Diggle SP, Buckling A, Gardner A, Griffin AS (2007) The social lives of microbes. Annu Rev Ecol Evol Syst 38: 53-77.
-
Kilstrup M, Hammer K, Ruhdal Jensen P, Martinussen J (2005) Nucleotide metabolism and its control in lactic acid bacteria. FEMS Microbiol Rev 29: 555-590.
-
Cocaign-Bousquet M, Garrigues C, Novak L, Lindley ND, Loublere P (1995) Rational development of a simple synthetic medium for the sustained growth of Lactococcus lactis. J Appl Bacteriol 79: 108-116.
-
Kunji ERS, Mierau I, Hagting A, Poolman B, Konings WN (1996) The proteotytic systems of lactic acid bacteria. Antonie Van Leeuwenhoek 70: 187-221.
-
Teusink B, Van Enckevort FHJ, Francke C, Wiersma A, Wegkamp A, et al. (2005) In silico reconstruction of the metabolic pathways of Lactobacillus plantarum: comparing predictions of nutrient requirements with those from growth experiments. Appl Environ Microbiol 71: 7253-7262.
-
Zhang G, Mills DA, Block DE (2009) Development of chemically defined media supporting high-cell-density growth of Lactococci, Enterococci, and Streptococci. Appl Environ Microbiol 75: 1080-1087.
-
McFarland LV (2008) Antibiotic-associated diarrhea: epidemiology, trends and treatment. Future Microbiol 3: 563-578.
-
Konstantinov SR, Smidt H, de Vos WM, Bruijns SCM, Singh SK, et al. (2008) S layer protein A of Lactobacillus acidophilus NCFM regulates immature dendritic cell and T cell functions. Proc Natl Acad Sci USA 105: 19474- 19479.
-
Gluck U, Gebbers J-O (2003) Ingested probiotics reduce nasal colonisation with pathogenic bacteria (_Staphylococcus aureus_, _Streptococcus pneumoniae_ and _β hemolytic streptococci_). Am J Clin Nutr 77(2): 517-520.
-
Bernstein JM, Haase E, F Scannapieco, Dryja D, Wolf J, et al. (2006) Bacterial interference of penicillin-sensitive and resistant _Streptococcus pneumoniae_ by _Streptococcus_ _oralis_ in adenoid organ culture: Implications for the treatment of recurrent upper respiratory tract infections in children and aduts. Ann Otol Belg Laryngol 115(5): 350-356.
-
Di Pierro F, Donato G, Formia F, Adami T, Careddu D, et al. (2012) Preliminary pediatric clinical evaluation of the oral probiotic _Streptococcus salivarius_ K12 in preventing recurrent pharyngitis and/or tonsillitis caused by _Streptococcus pyogenes_ and recurrent acute otitis media. Int J Gen Med 5: 991-997.
-
Luoto R, Ruuskanen O, Waris M, Kalliomäki M, Salminen S, et al. (2014) Prebiotic and probiotic supplementation prevents rhinovirus infections in preterm infants: a randomized, placebo-controlled trial. J Allergy Clin Immunol 133: 405-413.
-
Panigrahi P, Parida S, Nanda NC, Satpathy R, Pradhan L, et al. (2017) A randomized synbiotic trial to prevent sepsis among infants in rural India. Nature 548: 407- 412.
-
De Grandi R, Drago L, Bidossi A, Bottagisio M, Gelardi M, et al. (2019) Putative microbial population shifts attributable to nasal administration of Streptococcus salivarius 24SMBc and Streptococcus oralis 89a. Probiotics Antimicrob Proteins 11: 1219-1226.
-
Campos IB, Darrieux M, ferreira DM, Miyaji EN, Silva DA, et al. (2008) Nasal immunization of mice with Lactobacillus casei expressing the pneumococcal surface proteinA: induction of antibodies, complement deposition and partial protection against _Streptococcus_ _pneumoniae_ challenge. Microbes Infect 10(5): 481-488.
-
Vieira AT, Rocha VM, Tavares L, Garcia CC, Teixeira MM, et al. (2016) Control of Klebsiella pneumonae pulmonary infection and immunomodulation by oral treatment with commensal probiotic Bifidobacterium longum 51A. Microbes Infect 18: 180-189.
-
Varyukhina S, Freitas M, Bardin S, Robillard E, Tavan E, et al. (2012) Glycan-modifying bacteria-derived soluble factors from Bacteroides thetaiotaomicron and Lactobacillus casei inhibit rotavirus infection in human intestinal cells. Microbes and Infection14(3): 273-278.
-
More MI & Swidsinski A (2015) Saccharomyces boulardii CNCM I-745 supports regeneration of the intestinal microbiota after diarrheic dysbiosis - a review. Clin Exp Gastroenterol 8: 237-255.
-
Koh A, De Vadder F, Kovatcheva-Datchary P, Bäckhed F (2016) From Dietary Fiber to Host Physiology: short- ChainFatty Acids as Key Bacterial Metabolites. Cell 165: 1332-1345.
-
Cassir N, Benamar S, La Scola B (2016) Clostridium butyricum: from beneficial to a new emerging pathogen. Clin Microbiol Infect 22: 37-45.
-
Gaudier E, Jarry A, Blottière HM, De Coppet P, Buisine MP, et al. (2004) Butyrate specifically modulates MUC gene expression in intestinal epithelial goblet cells deprived of glucose. Am J Physiol Gastrointest Liver Physiol. 287: G1168-G1174.
-
Hoebler C (2004) Butyrate specifically modulates MUC gene expression in intestinal epithelial goblet cells deprived of glucose. Am J Physiol Gastrointest Liver Physiol 287: G1168-G1174.
-
Hagihara M, Kuroki Y, Ariyoshi T, Higashi S, Fukuda K, et al. (2020) Clostridium butyricum Modulates the Microbiome to Protect Intestinal Barrier Function in Mice with Antibiotic-Induced Dysbiosis. iScience 23: 100772.
-
Lee JS, Tato CM, Joyce-Shaikh B, Gulen MF, Cayatte C, et al. (2015) Interleukin-23-Independent IL-17 Production Regulates Intestinal Epithelial Permeability. Immunity 43: 727-738.
-
Zhang H-Q (2009) Therapeutic effects of Clostridium butyricum on experimental colitis induced by oxazolone in rats. World J Gastroenterol 15: 1821.
-
Berger A (2000) Th1 and Th2 responses: what are they. BMJ 321: 424.
-
Juan Z, Zhao-Ling S, Ming-Hua Z, Chun W, Hai-Xia W, et al. (2017) Oral administration of Clostridium butyricumCGMCC0313-1 reduces ovalbumin-induced allergic airway inflammation in mice. Respirology 22: 898-904.
-
Krystel-Whittemore M, Dileepan KN, Wood JG (2015) Mast Cell: a Multi-Functional Master Cell. Front Immunol 6: 620.
-
Leigh BA (2019) Cooperation among conflict: prophages protect bacteria from phagocytosis. Cell Host & Microbe 26(4): 450-452.
-
Keen EC, Dantas G (2018) Close encounters of three kinds: bacteriophages, commensal bacteria and host immunity. Trends in Microbiology 26(11): 943-954.
-
Ishida Y, Nakamura F, Kanzato H, Sawada D, Hirata H, Nishimura A, et al. (2005) Clinical effects of Lactobacillus acidophilus strain L-92 on perennial allergic rhinitis: a double-blind, placebo-controlled study. J Dairy Sci 88: 527-533.
-
Lin WY, Fu LS, Lin HK, Shen CY, Chen YJ (2014) Evaluation of the effect of Lactobacillus paracasei (HF. A00232) in children (6-13 years old) with perennial allergic rhinitis: a 12-week, double-blind, randomized, placebo-controlled study. Pediatr Neonatol 55: 181-188.
-
Jerzynska J, Stelmach W, Balcerak J, Woicka- Kolejwa K, Rychlik B, et al. (2016) Effect of Lactobacillus rhamnosus GG and vitamin D supplementation on the immunologic effectiveness of grass-specific sublingual immunotherapy in children with allergy. Allergy Asthma Proc 37: 324-334.
-
Kang HM, Kang JH (2021) Effects of nasopharyngeal microbiota in respiratory infections and allergies. Clin Exp Pediatr 64(11): 543-551.
-
Fulci V, Stronati L, Cucchiara S, Laudadio I, Carissimi C (2021) Emerging roles of gut virome in pediatric diseases. International Journal of Molecular Sciences 22(8): 4127.
- 4th Branchial Cleft Sinus Anomaly Presenting as Recurrent Thyroid Abscess in A Child: A Case Report
- Parotid Duct Injury Repaired Using an Angiocatheter Stent: A Case Report
- Organization and Functionality of the Referral and Counter-Referral System for ENT Disorders in District Hospitals of N’Djamena, Chad: A Cross-Sectional Analytical Study
- Facial Metastases from a Gastrointestinal Stromal Tumor: A Case Report
- Panorama of Ent Cancers and Literature Review: Epidemiological Profile and Therapeutic Management
- Could Antimicrobial Resistance Prove to Be Both a Threat and an Opportunity for us?