Amending the Description of Intraproboscis sanghae Amin, Heckmann, Sist, Basso, 2021 (Acanthocephala: Gigantorhynchidae) from a New Host, Phataginus tricuspis Rafinesque in Central Africa
Intraproboscis sanghae Amin, Heckmann, Sist, Basso, 2021 (Acanthocephala: Gigantorhynchidae) was described from 4 females of 7 specimens obtained from a black-bellied pangolin, Phataginus tetradactyla Linn. (Manidae) in the Central African Republic. No males were available at the time. We, however, came upon two specimens of the same species, a male and a female, collected from a white-bellied pangolin, Phataginus tricuspis Rafinesque in central Africa which makes this finding a new host record for I. sanghae. The female specimen was whole and its measurements fell within the range of measurements of females in the original description. The male specimen, designated as the allotype, was incomplete; missing the posterior extremity, but provided good comparative measurements of structures of females. All measured male structures were smaller than those of the females. The generic diagnosis of Intraproboscis is emended to include the allotype male. Basically, the description of I. sanghae is now more complete, except for the absence of the posterior portion of the male specimen.
Introduction
The white-bellied or tree pangolin, Phataginus tricuspis Rafinesque, is the new host of Intraproboscis sanghae Amin, Heckmann, Sist, Basso, 2021 (Gigantorhynchidae) originally reported from females in the Central African Republic. The semi-arboreal nocturnal tree pangolin is the most common forest pangolin in equatorial Africa spreading from Guinea through Sierra Leone and West Africa to Central Africa, Kenya and Tanzania, then to Angola and Zambia as well as to Senegal, Gambia and Guinea-Bissau [1]. Intraproboscis sanghae was originally described from females by Amin, et al. [2] from its close relative, the black-bellied (long-tailed) pangolin Phataginus tetradactyla Lin_. Phatiginus tricuspis_ has myrmecophagous diet including ants and termites from their nests and other insects moving on trees [3]. Other species of pangolins feed also on termites, and soft-bodied invertebrates like bee larvae, flies, worms, earthworms and crickets [4].
In the first comprehensive checklist of parasites of all 8 spe- cies of pangolins, Mohapatra, et al. [5] reported a total of 34 genera of parasites including 13 genera of helminths (8 intestinal): Cylicospirura, Leipernema, Cylicospirura, Leiper-
nema, Manistrongylus, Necator, Strongyloides, Trichochenia, Ancylostoma, Gendrespirura) but no acanthocephalans, 8 genera of ticks, 2 genera of mites and 7 genera of bacteria. A comprehensive systematic treatise of pangolins with empha- sis on conservation, evolution, ecology, taxonomy, behavior, and protection was recently published by Challender, et al. [6]. Amin, et al. [2] reviewed the parasitic fauna of African and Asian pangolins. Barton, et al. [7] reviewed the nema- todes and acanthocephalans of confiscated Sunda pangolins, Manis javanica Desmarest and updated the list of the para- sites of pangolins.
Conservation efforts to “save” the pangolins are also directed to better understanding their relationships to public health since pangolins are also known reservoirs of human pathogenic bacteria such as Proteus vulgaris, Klebsiella pneumoniae, and Escherichia coli. Pseudomonas aeruginosa and K. pneumoniae are opportunistic pathogens in humans that cause a variety of bronchopulmonary infections including pneumonia [8, 9, 10]. Although most E. coli are not pathogenic, some serotypes possess virulence factors and cause gastroenteritis in humans [9, 10]. Our research efforts with the black-bellied and white-bellied pangolins, P. tetradactyla and P. tricuspis in Central Africa add a new dimension to the understanding of the parasitic picture of pangolins.
Materials and Methods
Collections
The present study is based on 2 pseudo-segmented acanthocephalans collected from one white-bellied pangolin, P. tricuspis. The worms were extracted and preserved in 70% ethanol. These 2 worms were originally obtained from New Jersey vets working on an illegal shipment of pangolins from an unrecorded geographical location in central Africa. These worms ultimately made their way to Dr. Heather Walden at the University of Florida College of Veterinary Medicine at Gainesville, who provided them to us in December, 2020 at the Institute of Parasitic Diseases, Arizona for diagnosis and investigation. The specimens were routinely processed for microscopy and studied. We have subsequently transferred the custody of the 2 specimens to the University of Nebraska’s State Museum’s Harold W. Manter Laboratory (HWML), Lincoln collection for final deposit. We have attempted to obtain males of I. sanghae from P. tetradactyla and P. tricuspis in different locations in Central Africa but have been unsuccessful. Many specimens and other intestinal material collected from vomit and feces of these 2 host species in 2018 and 2020 were provided to us courtesy of Drs. Tamar Cassidy and Marine Montblanc of the Sangha Pangolin Project at Bayanga in the Central African Republic but they included no acanthocephalans.
Methods for Microscopical Studies
The worms were punctured with a fine needle and subsequently stained in Mayer’s acid carmine, destained in 4% hydrochloric acid in 70% ethanol, dehydrated in ascending concentrations of ethanol (24 hr each), and cleared in 100% xylene then in 50% Canada balsam and 50% xylene (24 hr each). Worms were then whole mounted in Canada balsam. Measurements are in micrometers or in mm as noted. The range is followed by mean values between parentheses. Width measurements represent maximum width. Trunk length does not include proboscis.
Line Drawing
Line drawings were created by using a Ken-A-Vision micro-projector (Ward’s Biological Supply Co., Rochester, N.Y.) which uses cool quartz iodine 150W illumination. Color-coded objectives, 10X, 20X, and 43X lenses, are used. Images of stained whole mounted specimens were projected vertically on 300 series Bristol draft paper (Starthmore, Westfield, Massachusetts), then traced and inked with India ink. Projected images were identical to the actual specimens being projected. The completed line drawings were subsequently scanned at 600 pixels, transferred on a USB and subsequently downloaded on a computer.
Optical Microscope Images
Optical microscope images were acquired using a BH2 light Olympus microscope (Olympus Optical Co., Osachi- shibamiya, Okaya, Nagano, Japan) attached to an AmScope 1000 video camera (United Scope LLC, dba AmScope, Irvine, California), linked to an ASUS laptop equipped with HDMI high-definition multimedia interface system (Taiwan- USA, Fremont, California). Images from the microscope are transferred from the laptop to a USB and stored for subsequent processing on a computer.
Deposits
The allotype male and paratype female were deposited in the University of Nebraska’s State Museum’s Harold W. Manter Laboratory (HWML) collection in Lincoln, Nebraska.
Results
The following is a morphological account of the 2 specimens of I. sanghae collected from P. tricuspis described herein.
The Female
The female specimen was 81.25 mm long and 1.37 mm at its widest point. All other measurements fell within the range of those of the other females described by Amin, et al. [2] and noted in Table 1. This one specimen was, however, unique in having a partially retracted proboscis (Figure 1) which provided an opportunity to assess the operation of the receptacle which otherwise is totally embedded within the proboscis when normally extended as described in Amin, et al. [2].
| Specimens | Females N = 4 (whole) | Male N = 1 (anterior) |
|---|---|---|
| Source | Amin, et al. [2] | This paper |
| Host | Phataginus tetradactyla Linn. | Phataginus tricuspis Rafinesque |
| Locality | Dzanga Sangha, Central African Republic | |
| Trunk length (mm) | 63.75 – 180.00 (108.35) | 94.25 |
| Max. trunk width (mm) | 1.12 – 2.00 (1.50) | 1.5 |
| Proboscis length (mm) | 1.56-1.87 (1.74) | 1.37 |
| Ant. proboscis width (mm) | 0.27 – 0.41 (0.34) | 0.29 |
| Post. proboscis width (mm) | 0.58 – 0.74 (0.67) | 0.44 |
| Proboscis hook rows | 34 -36 | 32 |
| Proboscis hooks per row | 6-7 | 6 |
| Spine-like hooks per row | 15 – 17 (not regularly alternating) | 15 (not regularly alternating) |
| Ant. Hook length X width (µm) | 38 - 44 X 9 - 11 | 33 X 8 |
| Mid. hook length X width (µm) | 40 – 50 X 12 - 14 | 37 X 11 |
| Post. hook length X width (µm) | 47 – 54 X 15 - 16 | 42 X 13 |
| Spine-like hook length X width (µm) | 20 – 25 X 5 - 7 | 18 X 4 |
| Proboscis receptacle length (mm) | 0.77 – 1.04 (0.90) | 0.68 |
| Proboscis receptacle Width (mm) | 0.26 – 0.36 (0.31) | 0.21 |
| Cephalic ganglion length X width (µm) | 166–229 (197) X 60–100 (80) | 208 X 94 |
| Lemnisci length (mm) | 2.08 -3.28 (2.86) | 4.00 |
| Lemnisci width (mm) | 0.22 – 0.38 (0.28) | 0.42 |
| Anterior testis length X width (mm) | N/A | 2.90 X 0.60 |
| Posterior Testis length X width | N/A | 1.32 X 0.55 |
| Ant. Trunk to ant. testis (mm) | N/A | 88.25 |
Table 1: Comparative morphometrics of male and female specimens of Intraproboscis sanghae from pangolins in Central Africa.
The Male
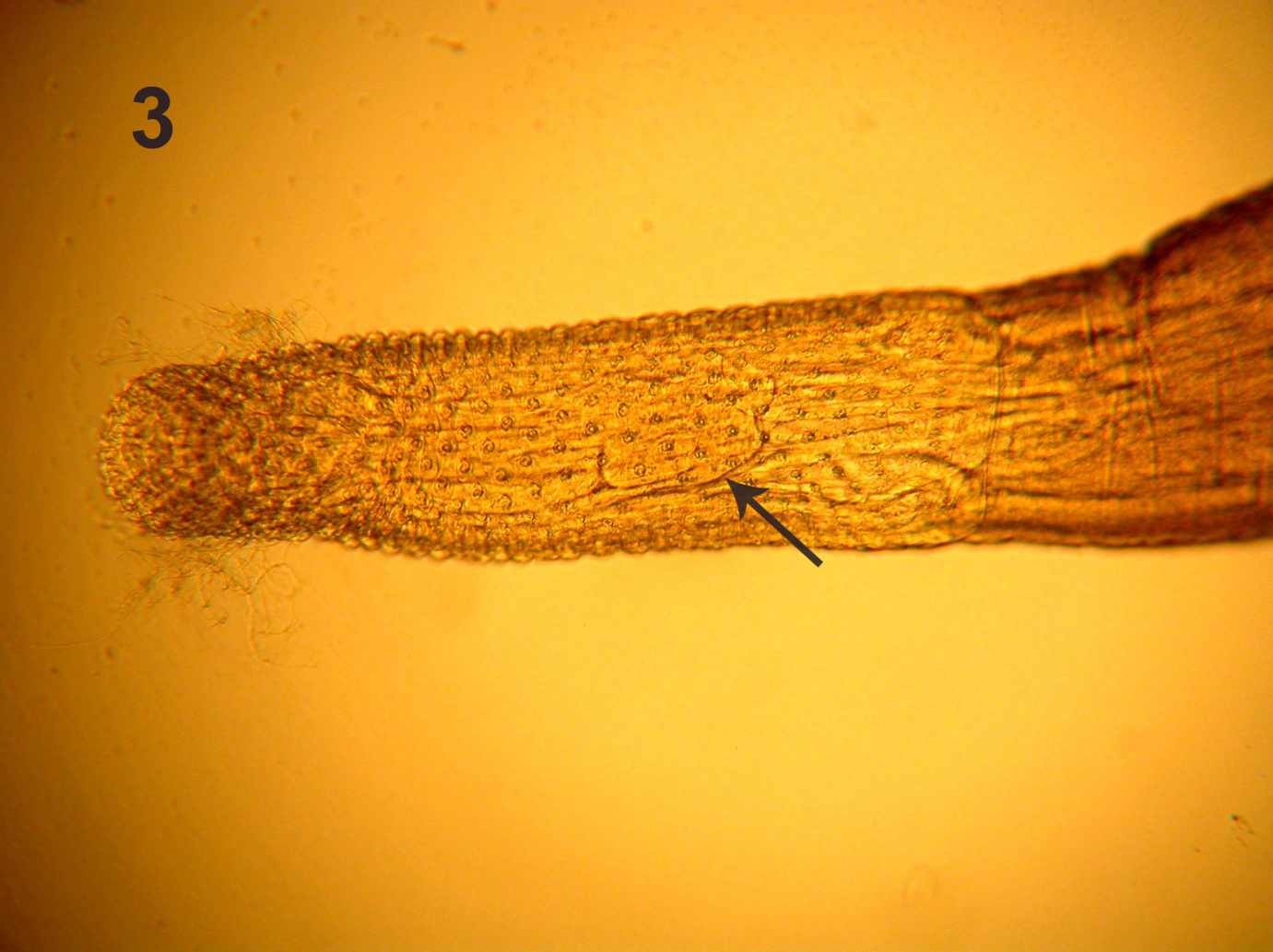
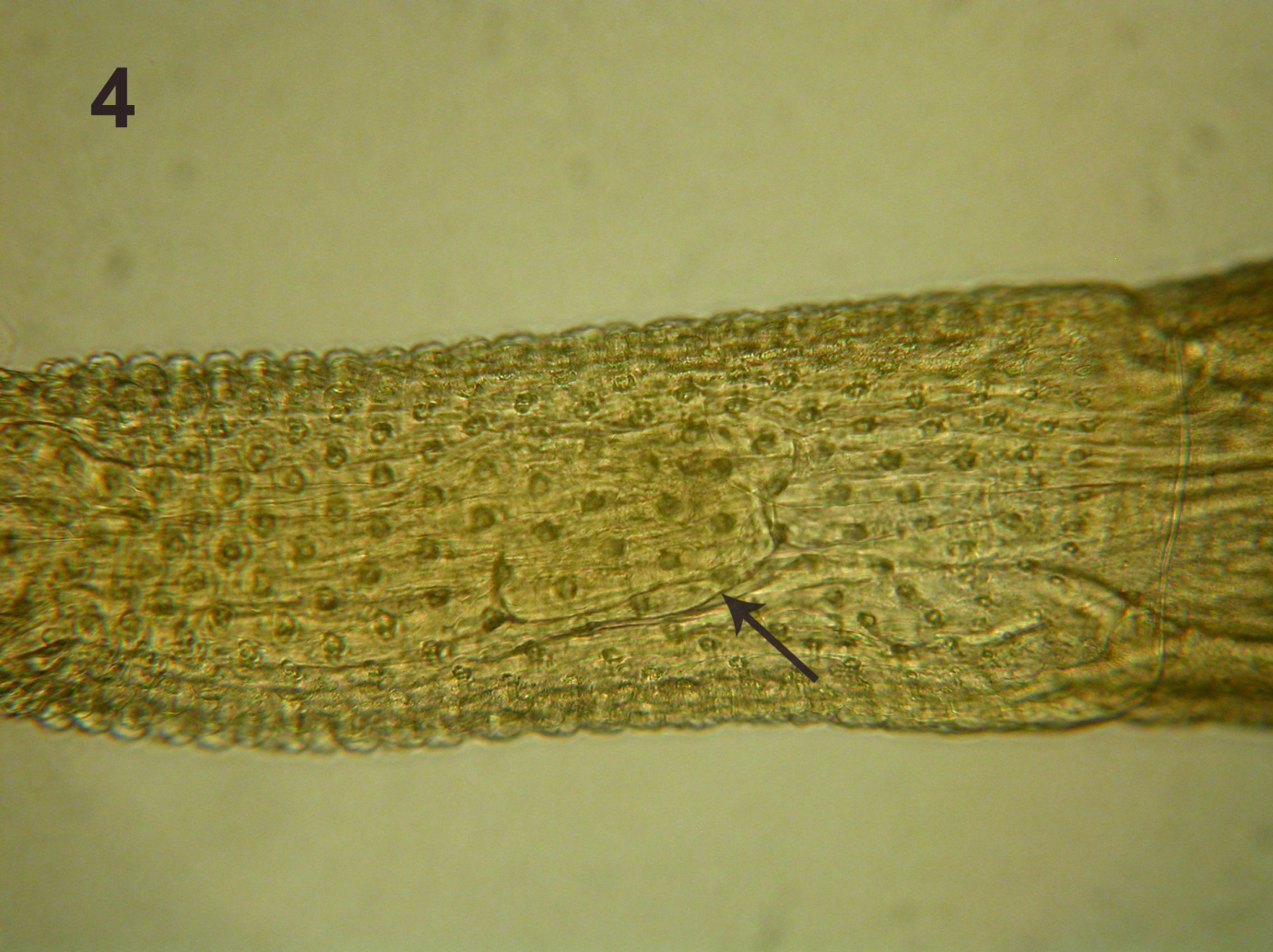
This perfect specimen was incomplete posteriorly extending from the fully evaginated proboscis to the partially wrinkled posterior testis (Figure 2). The long lemnisci, Taenia-like pseudo-segmentation of trunk, and the imposing shape and size of the anterior testis were clearly demonstrable. Measurements of this male specimen are compared with those of the females of the same species as reported by Amin, et al. [2] (Table 1). Measurements of all structures in the male were smaller than those of the females but the lemnisci were slightly longer. Being the only male specimen of I. sanghae available, it is herein designated as the Allotype.
Taxonomic Summary
Host: Phataginus tricuspis Rafinesque. (Manidae) Type host: Phataginus tetradactyla Linn. (Manidae) Locality: Undetermined location in Central Africa. Type locality: SW Central African Republic at the Dzanga Sangha Protected Area (3°6′N 16°20′E). Site of infection:
Intestine.
Specimens: University of Nebraska’s State Museum’s Harold W. Manter Laboratory (HWML), Lincoln coll. no. 216916 (allotype male on one slide; Figure 2) and no. 216917 (paratype female on two slides; Figure 1). The HWML accession form included the following statements.
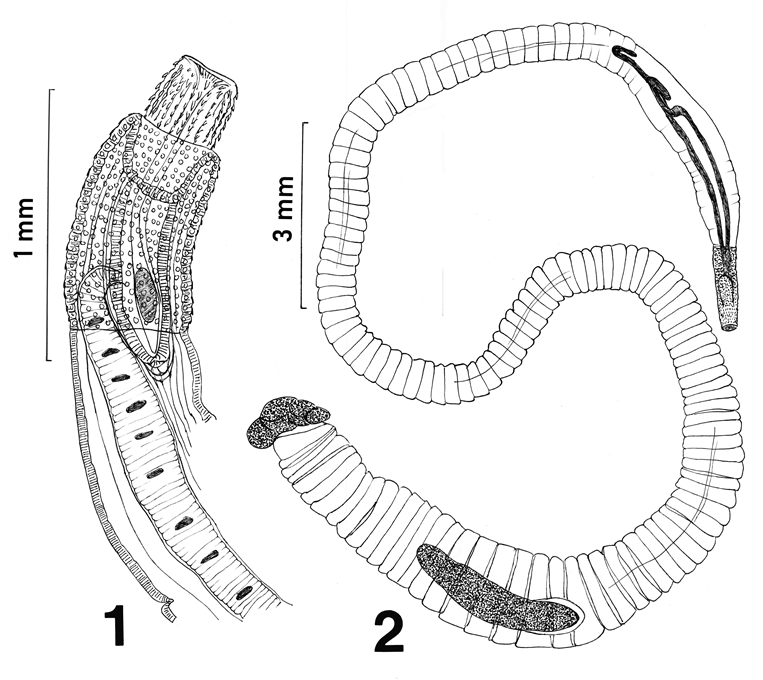
Line drawings of a female and male specimens of Intraproboscis sanghae collected from Phataginus tricuspis in Central Africa. Figure 1: The anterior part of the female specimen showing the retracted anterior proboscis, the posterior displacement of the proboscis receptacle, and insertion of a lemniscus and the receptacle. Figure 2: The posteriorly incomplete male specimen showing the characteristic pseudo-segmentation, everted proboscis, long lemnisci, and complete anterior testis. “Additional notes: specimens were collected in New Jersey while working on an illegal shipment of pangolins.
The specimens were sent to Dr. Heather D. S. Walden (University of Florida College of Veterinary Medicine, Department of Comparative, Diagnostic and Population Medicine) who sent them to Omar Amin for further processing, identification and study. No detailed records are available on where and when the pangolins were originally collected except that they are from somewhere Central Africa.”
Discussion
The Female
Structural details of the female specimen agreed closely with those of the other 4 females in the original description and its measurements fell within the range of reported measurements [2]. The proboscis was, however, partially retracted. The type of proboscis characteristic of these worms apparently only retracts but is not equipped to invaginate (Figure 1). Upon retraction, the receptacle appeared to get displaced more posteriorly extending well beyond the posterior end of the proboscis. The insertion and detail of the anteriorly nucleated lemnisci are newly documented. Compare the line drawing (Fig. 1) with the line drawing of the anterior part of a female specimen with totally everted proboscis in Amin, et al. [2].
The Male
Although the posterior-most part of the male specimen was missing, the morphology and measurements of the anterior larger part of the specimen including the proboscis, hooks, receptacle and lemnisci were conclusive of its identity as I. sanghae (Fig. 2) and justifying its assignment as the allotype. This statement is more readily documented when considering the similarities of this male specimens with those of the females in Amin, et al. [2] (Figures 1-4 of anterior trunk, proboscis, hooks, receptacle, and lemnisci). Practically, all male measurements were comparable to those of female I. sanghae except for being somewhat smaller than those of the females (Table 1) depicting the usual sexual differentiation between male and female acanthocephalans. The only exception is the size of lemnisci that appeared to be slightly longer in the male specimen compared to the females. Sample size of 1 may be a factor. In addition to the size and shape of characters in common with the females (Table 1), the size, shape, position of the anterior testis and its distance from anterior trunk, and the fact that testes are not contiguous are important diagnostic features of males of I. sanghae. The generic diagnosis is herein emended to reflect this additional information of the male anatomy (Fig. 2).
Addendum to the Diagnosis of Intraproboscis
Shared structures of males morphologically similar to and relatively smaller than those of females. Testes large, oblong, not contiguous. Anterior testis distant from anterior end of trunk.
Conclusion
This study completes the description of I. sanghae as best as possible given the availability of specimens. Amin, et al. [2] outlined the health and conservation issues related to the pangolin “industry” in central Africa that often targets pangolins for intensive exploitation for bush meat and for subsequent trades utilizing their hard scales for various industries and drugs. Our project highlights one of the various ways by which pangolins are being exploited and their populations degraded. Completing the description of one of the pathogenic organisms that would adversely affect their health is one step to the better understanding of the host-parasite relationships affecting their survival and well- being and to evaluating their conservation strategies.
Acknowledgment
We recognize the New Jersey vets who have been the original source of studied specimens from the illegal shipment of pangolins originating in central Africa and Dr. Heather Walden of the University of Florida College of Veterinary Medicine at Gainesville, who made them available to us for analysis. We also extend our gratitude to Drs. Tamar Cassidy and Marine Montblanc of the Sangha Pangolin Project at Bayanga in the Central African Republic for their tireless efforts to procure male specimens of I. sanghae. I am also especially grateful to Dr. Nataliya Yu. Rubtsova at the Parasitology Center, Inc. (PCI), Scottsdale for her expert help in the preparation of the presented visuals (Figures 1-4). This project was supported by an Institutional Grant from PCI.
Declarations
- Compliance with Ethical Standards
- Conflict of Interest: The authors declare no conflicts of interest or competing interests.
- Ethical Approval: The authors declare that they have observed all applicable ethical standards.
- Availability of Data: All presented and related data are available by contacting the author. Approval protocols for retrieval of these two specimens were not available as the confiscated pangolins were smuggled from Central Africa to New Jersey where they were examined for the first time.
References
-
Kingdon J, Hoffman M (2013) Genus _Phataginus_ Tree Pangolins_,_ Mammals of Africa, Volume 5, Carnivores, Pangolins, Equids and Rhinoceroses, Bloomsbury Publishers.
-
Amin OM, Heckmann RA, Sist B, Basso WU (2021) A review of the parasite fauna of the black-bellied pangolin, _Phataginus tetradactyla_ Lin. (Manidae), from central Africa with the description of _Intraproboscis sanghae_ n. gen., n. sp. (Acanthocephala: Gigantorhynhidae). J Parasitol 107: 222-238.
-
Jansen R, Sodeinde O, Soewu D, Pietersen DW, Alempijevic D (2020) Challender DWS. Nash HC, Waterman C (Eds.), Chapter 9-White-bellied pangolin _Phataginus tricuspis_ (Rafinesque, 1820). _Pangolins_, Biodiversity of World: Conservation from Genes to Landscapes, Academic Press, pp: 139-156.
-
Shi VQ, Wang YQ (1985) Food habits in Pangolin. Chinese Journal of Wildlife 6: 42-43.
-
Mohapatra RK, Panda S, Nair MV, Lakshini NA (2015) Checklist of parasites and bacteria recorded from pangolins. J Parasitic Dis 10:1-7.
-
Challender DWS, Nash HC, Waterman C (2020) Pangolins: Science. Society and conservation (eds.). In Biodiversity of the World, Conservation from Genes to Landscapes. Academic Press (Elsevier), London, UK, pp: 630.
-
Barton DP, Martelli P, Worthington BM, Lam TTY, Zhu X, et al. (2022) Nematode and acanthocephalan parasites of confiscated Sunda pangolins, _Manis javanica_ (Mammalia: Pholidota: Manidae), with an updated list of the parasites of pangolins. Diversity 14 (12): 1039.
-
Domingo P, Ferre A, Baraldes MA, Ris J, Sanchez F (1998) _Pseudomonas aeruginosa_ bronchopulmonary infection in patients with AIDS, with emphasis on relapsing infection. Eur Respir J 12: 107-112.
-
Lightfoot NF (2003) Bacteria of potential health concern. In: Bartram J, Cotruvo J, et al. (Eds.), Heterotrophic plate counts and drinking-water safety pp: 119-137.
-
Amin OM (2011) The contribution of pathogenic bacteria to GI symptoms in parasite-free patients. J Bacteriol Parasitol 2: 2.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework